
АКАДЕМИЯ НАУК СССР
ИНСТИТУТ БИОЛОГИЧЕСКОЙ ФИЗИКИ
с. э. шноль
ФИЗИКО
ХИМИЧЕСКИЕ
ФАКТОРЫ
БИОЛОГИЧЕСКОЙ
ЭВОЛЮЦИИ
ИЗДАТЕЛЬСТВО «НАУКА» МОСКВА 1979
Физико-химические факторы биологической эволюции. Ш н о л ь С. Э.
М., 1979, 263 с.
Рассмотрены физико-химические факторы биологической эволюции, приводящие к возникновению ферментов, аппаратов перемещения в пространстве, систем обеспечения энергией, внутриклеточных структур, клеток, нервной системы. Книга представляет интерес для широкого круга читателей — биологов, физиков, химиков, философов.
Табл. 11, ил. 34, библиогр. 474 назв.
Ответственные редакторы
академик Г. М. ФРАНК, член-корр. АН СССР Г. Р. ИВАНИЦКИЙ
21005—125 2001040000 © Издтельство «Наука*, 1979 г.
055(02)—79
ПРЕДИСЛОВИЕ (ОТВЕТСТВЕННОГО РЕДАКТОРА
В наше время замечательных достижений в биологии поток новых фактов буквально заливает литературу. Быстро изменяются представления о биофизических и биохимических основах явлений жизни. В такой ситуации понятно стремление оглядеться, попытаться представить себе, в какой мере классические основы биологии сочетаются с новыми данными.
Такую попытку и представляет собой книга С. Э. Шноля. Автор исследует в ней возможность дедуктивного метода в современной биологии. С этой целью он прослеживает возможный путь эволюционного становления основных биофизических, биохимических и физиологических свойств живых организмов, а затем кратко рассматривает данные о тех же явлениях, полученные в результате достижений современной науки. Такой подход, интересный сам по себе, позволяет обратить внимание читателя на некоторые узловые вопросы биофизики и биохимии, еще ожидающие своего решения.
Конечно же, не во всех своих разделах анализ автора достаточно последователен и совершенен. Однако основная ценность этой книги — опыт единого в сущности подхода к явлениям жизни на всех этапах биологической эволюции. Автор справедливо подчеркивает, что единственной общебиологической теорией является дарвинизм. Его попытка соединения дарвинизма с современными достижениями биофизики, биохимии, молекулярной биологии в значительной мере есть следствие появления более 50 лет назад классической работы А. И. Опарина «О происхождении жизни».
Книга С. Э. Шноля выходит в свет в период повышенного интереса к проблемам биологической эволюции. Широкую известность, в частности, получила недавняя работа М. Эйгена (1973) «Саморегуляция материи и эволюция биологических макромолекул». Следует отметить, что обе эти работы удачно дополняют друг друга.
М. Эйген сэсредоточил свое внимание в основном на выведении уравнений, связывающих скорость размножения биологических макромолекул данного вида со скоростью и качеством их репликации и гибели.
С. Э. Шноль использует введенное им понятие «кинетическое совершенство» для анализа не только самых начальных, но и последующих этапов эволюции. Примененный им метод («принцип предельного совершенства на каждом этапе эволюционного пути») аналогичен популярному в физике и математике методу предельного перехода.
Следует отметить также, что рассмотрение даже самых серьезных вопросов С. Э. Шноль проводит живо, хорошим литературным языком, не опасаясь местами патетики и даже юмора. Это — хорошо, поскольку показывает до некоторой степени читателк> живую атмосферу научного поиска.
В целом можно рекомендовать эту книгу читателю, интересующемуся проблемами общей биологии, биохимии и биофизики, во всяком случае как источник вопросов, побуждающих думать над их возможными решениями
22 ноября 1973 г.
{Академик Г. М. Франк
Высшим долгом физиков является поиск тех общих элементарных законов, из которых путем чистой дедукции можно получить картину мира.
А. Эйнштейн. Физика и реальность.
Мы унаследовали от наших предков острое стремление к цельному, всеобъемлющему знанию. Само название высших институтов познания— университеты — напоминает нам, что с давних пор и на протяжении многих столетий универсальный характер знаний — единственное, к чему может быть полное доверие...
Э. Шредингер. Что такое жизнь?
ПРЕДИСЛОВИЕ АВТОРА
Широко распространено убеждение в первенствующей роли биологии среди других наук во второй половине XX в. Мысль эта наиболее четко высказывалась физиками (Жолио Кюри, Бернал). Многие выдающиеся физики стали в той или иной форме заниматься биологией и многие внесли в биологию значительный непосредственный вклад (Гамов, Крик, Полинг, Сциллард, Дельбрюк и др.). В высказываниях физиков и в устремлении физиков в биологию проявилась связь наук — то, что наука о природе едина и лишь в человеческом сознании, в силу «невозможности объять необъятное», а также в результате стремления к последовательности и к систематизации знаний она разделена на математику, физику, биологию и др.
Однако дело не только в этом. Именно в физике в начале нашего века произошла общеизвестная революция — были созданы теория относительности и «вантовая механика.
Бурный расцвет новой физики, когда принципиально важные открытия следовали одно за другим с драматической быстротой и приводили к важнейшим теоретическим и практическим результатам, документирован и описан в ряде книг.
Естественно, что именно физики, только недавно пережившие революцию в своей науке, знающие «как это делается», обратились к биологии, где все, как казалось, предвещало аналогичные события.
Их ожидания оправдались. В биологии в самом деле произошла революция, и она действительно явилась результатом прогресса физико-химического направления в биологических ис^ следованиях.
Принято датировать начало революции в биологии 1953 г., когда была опубликована статья Уотсона и Крика о строении ДНК- За прошедшие 20—25 лет преобразились наши знания омолекулярных механизмах наследственности, изменчивости, превращениях энергии в клетке, о механизме биосинтеза основных компонентов протоплазмы — белков, нуклеиновых кислот, липидов, полисахаридов.
Возникла новая наука — молекулярная биология. Но революция в биологии отнюдь не началась статьей Уотсона и Крика. Это было лишь закономерное продолжение открытий Дарвина, Менделя, Моргана, лишь закономерное развитие действительно фундаментальной и революционной идеи XX в. — идеи матричного механизма репликации молекул биополимеров, принадлежащей Н. К- Кольцову. Устремление физиков и физики в биологию действительно оказало большое влияние на темп революционных событий в биологии. Дело не только в проникновении в биологию методов физического и химического исследований. Физика влияет на биологию своим примером, стилем работы, методологией мышления. Именно в физике особо плодотворной оказалась теория. Теоретическая физика замечательна своим ярко выраженным дедуктивным характером, тем, что она является не столько обобщением фактов, сколько развитием исходных общих положений. Триумфы современной физики — это экспериментальное подтверждение предсказаний теории. Получение основных положений какой-либо науки из общих соображений — необходимая завершающая стадия развития мысли в данном направлении. И здесь теоретическая биология сильно отстает от теоретической физики.
Фундаментом современной теоретической биологии служит дарвинизм. Однако после упомянутых физико-химических, «молекулярных» потрясений, после создания «молекулярной биологии» стала ясной недостаточная развитость теоретической биологии. Необходима такая теория биологии, в которой соединены классический эволюционный и новый молекулярный подходы. Возможно по аналогии с физикой и разделение теоретической биологии на макро- и микротеории, однако истинная теория должна быть единой и всеобъемлющей.
Эта предлагаемая вниманию читателя книга — результат убеждения в том, что время для теоретического соединения молекулярной биологии и дарвинизма наступило. Необходимо пытаться получить основные закономерности биологии из общих соображений, построить теорию, позволяющую предсказывать как динамику и траектории эволюционного процесса, так и молекулярные механизмы, функционирующие в биологических системах. Весьма заманчиво получить такие предсказания, а потом посредством эксперимента или обобщения ранее найденных данных сопоставить предсказанное с действительностью. Так биология приблизится к физике в самом главном—в методе научного познания.
Пытаясь рассмотреть биологические закономерности с единых позиций, я прослеживаю далее физические и физико-химическиеусловия начала биологической эволюции и основные критерии естественного отбора в системах полимерных молекул, для которых характерно матричное воспроизведение. При таком исследовании критериями отбора служат физические и физико-химические свойства макромолекул и их комплексов.
Более того, весь процесс биологической эволюции от первичных матричных молекул до высоко совершенных животных и растений можно рассматривать как последовательное, поочередное действие различных физических и физико-химических критериев естественного отбора, т. е. последовательное «включение» различных факторов эволюции.
В настоящее время, конечно, нереально, руководствуясь только общими соображениями, проследить, например, посредством ЭВМ, точные траектории эволюционного процесса. Однако это пока и не нужно. Сейчас вполне достаточен термодинамический подход — выявление лишь начальных и конечных состояний в каждом направлении эволюции. Тогда оказываются ненужными детальные вычисления и становится возможной оценка предельных состояний. Так, если ведущим критерием естественного отбора (т. е. фактором эволюции) при становлении биохимических реакций служит их скорость, легко в принципе оценить, когда дальнейшее совершенствование ферментов станет бесполезным из-за медленности диффузионных потоков реагентов и продуктов. Ведущим критерием отбора, фактором эволюции станет интенсивность перемешивания, и предел возможностей биологической эволюции на данном этапе также можно будет оценить по предельно достижимой скорости перемещения организмов в пространстве. Применительно к биологии такой термодинамический подход выглядит как «принцип предельного совершенства» при анализе основных этапов эволюции.
Можно представить себе опасения, возникающие у читателя книги, озаглавленной «Физико-химические факторы биологической эволюции». Не сводит ли автор биологию только к физике и химии? В сущности, это опасение понятно. И чтобы рассеять его, нужно сначала определить, что такое собственно биология? Не служит ли установление физико-химических механизмов биологических явлений доказательством отсутствия биологической специфики? Нет. Не служит. Биологическая специфика состоит в эволюционном способе (в ходе естественного отбора) формирования всех механизмов и свойств биологических систем. Поэтому собственно биологической является именно теория эволюции. Однако сам ход, этапы, траектории эволюционного процесса вполне подлежат изучению с позиций математики, физики, физической химии (см. нашу статью [340]).
Именно учитывая биологическую специфику, эволюционный способ возникновения всех «механизмов» в биологических системах, автор рассматривает физико-химические факторы эволюции, общие закономерности хода эволюционного процесса, матричного воспроизведения, ферментативного* катализа, превращения энергии, образования дискретных организмов, возникновение ионной асимметрии, появления внутриклеточных структур, механизмы морфогенеза и перемещения в пространстве. Из общих соображений выведена необходимость многоклеточности, наличия рецепторов, нервной системы, мозга.
Естественно, что весь широкий круг вопросов нельзя рассмотреть с равной компетентностью. Я остро ощущаю свой дилета- низм в очень многих затронутых проблемах, но тем не менее, не могу преодолеть привлекательности попытки общего взгляда на современную биологию.
Эта книга сложилась в результате многократного чтения соответствующего курса лекций студентам-биофизикам физического факультета МГУ. Обычно лекции проходили на Беломорской биостанции МГУ. Часто на открытом воздухе, вблизи нового лабораторного корпуса, среди заросших лишайниками валунов, около моря, под шум сосен и крики чаек мы обсуждали общие проблемы биологии. Живая реакция слушателей стимулировала работу над текстом лекций и рукописи. Мне хочется с благодарностью отметить огромный труд директора Беломорской биостанции Н. А. Перцова по созданию этого уникального места для педагогической и научной работы.
Работа над текстом рукописи в ее разных вариантах длилась много лет (с 1967 г.). За это время состоялось множество обсуждений, семинаров, докладов, дискуссий. Особенно полезными были обсуждения затронутых в книге вопросов с Н. В. Тимофее- вым-Ресовским, Л. А. Блюменфельдом, А. М. Молчановым, М. Н. Кондрашовой, Б. Я- Виленкиным, В. И. Дещеревским, Н. Г. Есиповой, Ю. Н. Полянским, М. В. Георгадзе, С. В. Багоц- ким, В. Н. Веховым, В. И. Брусковым, В. Н. Морозовым, Ф. И. Атауллахановым, А. П. Сарвазяном.
Я очень благодарен Г. М. Франку, А. И. Опарину, Г. Р. Иваницкому, Н. А. Райской, В. П. Скулачеву, Э. Э. Шнолю, прочитавшим рукопись и сделавшим ряд ценных критических замечаний. Н. Г. Есиповой написана часть текста гл. 3 и 11. Приложение написано совместно с А. С. Кондрашовым.
Наконец, я должен выразить особую благодарность сотрудникам ОНТИ Института биофизики за неоценимую помощь в подготовке рукописи к изданию.
Работа над текстом этой книги в основном была завершена осенью 1973 г. За прошедшее время некоторые предположения получили независимое развитие в работах других исследователей. То, что казалось экстравагантным, стало обычным. Одной из трудных проблем, возникших в связи с долгим изданием этой книги, оказалось составление списка литературы. Автор чувствовал себя в литературном море как моряк в дырявой лодке — новая литература течет в книгу «сквозь все щели». Дополнения списка литературы лишь отчасти решают эту проблему.
Глава 1
ФИЗИКО-ХИМИЧЕСКИЕ МЕХАНИЗМЫ и биологическая специфичность
Дарвинизм — основа теоретической биологии.
Краткий исторический очерк.
Физические и физико-химические критерии биологического прогресса. Термодинамические характеристики и кинетическое совершенство.
Наиболее важным достижением биологии XIX в. является дарвинизм, лежащий в основе теоретической биологии *. Центральная идея биологии XX в. — концепция матричного молекулярного механизма размножения организмов. Важная задача современной теоретической биологии состоит в выяснении связи матричной концепции с дарвинизмом [17, 23, 24, 129, 262, 288, 289].
Формулирование матричного механизма репликации молекул нуклеиновых кислот как молекулярной основы размножения организмов не только не уменьшает значения дарвинизма, но помогает выявить его действительный фундамент, позволяет понять молекулярные механизмы основных биологических явлений, составляющих предмет дарвинизма. В этом смысле современная молекулярная биология ■— составная часть дарвинизма, теоретической биологии.
Правильность сказанного следует из логического развития следствий матричной концепции. В результате матричного воспроизведения соответствующих молекул в определенных условиях происходят такие процессы: 1) конкуренция размножающихся матричных молекул за вещество, энергию, пространство; 2) вариабельность (изменчивость) матричных молекул; 3) воспроизведение в матричных копиях всех вариантов (конвариантноевоспроизведение, редубликация [288]); 4) естественный отбор, т. е. сохранение и размножение лишь более приспособленных; 5) эволюция, т. е. развитие матричных систем в направлении все большего совершенства, все большей итоговой интенсивности преобразования веществ окружающей среды в вещества данного вида (кинетическое совершенство [339]). Возможность такого в принципе независимого от осуществленного Ч. Дарвиным дедуктивного формулирования дарвинизма посредством анализа молекулярных основ биологии представляет собой важнейшее следствие происшедшей в биологии революции.
1 С современным состоянием и проблемами дарвинизма можно ознакомиться, по статьям, книгам [3, 9, 15, 105, 106, 107, 109, 127, 173, 183, 189, 189а, 199, 202, 241, 252, 289, 290, 304, 332, 333, 359].
Одна из основных задач этой книги состоит в иллюстрации «физико-химической обусловленности дарвинизма» ‘.
Установление обусловленности биологической эволюции фи- зико-химическими свойствами матричных молекул, физико-химический анализ основных ее этапов отнюдь не означает сведения биологии к физике или химии.
Биология отличается ют других .наук тем, что ее объекты представляют результат более или менее длительной эволюции. Они являются, в известном смысле, «концентратами прошедшего времени».
Одна из причин излишних споров «о сводимости» состоит в отсутствии достаточно точных определений основных понятий биологической эволюции, жизни, живого организма.
Я думаю, что настало время попытаться определить их, имея в виду сказанное о дарвинизме как основе теоретической биологии, и о синтезе дарвинизма с молекулярной биологией (см. [339, 340]).
Критерием правильности теории является возможность дедуктивных построений, соответствующих действительности. Поэтому основное содержание предлагаемой книги — опыт дедуктивного построения, попытки «выведения» в качестве следствий из общих, по преимуществу физико-химических соображений основных этапов биологической эволюции, основных свойств живых организмов.
Прежде чем приступить к выполнению этих задач, мне кажется необходимым кратко проследить историю становления концепции матричного механизма размножения организмов. Матричный механизм был сформулирован сначала на основании аналогии между размножением организмов и размножением кристаллов. Аналогия роста (размножения) кристаллов и живых организмов казалась первоначально лишь внешней. Она не имела (и не могла иметь в момент своего возникновения) серьезных оснований. Мысль о такой аналогии возникала на протяжении последних столетий неоднократно. Эту мысль в яркой форме высказывал еще Бюффон (середина XVIII в.). В прошлом веке аналогия с кристаллами была развита Геккелем для описания размножения надмолекулярных агрегатов (пластидул), из которых состоит протоплазма. Л. Я- Бляхер так излагает взгляды Геккеля: «...по мысли Геккеля, пластидулы не размножаются делением, а возникают в питательной жидкости подобно тому, как появляются новые кристаллы под влиянием кристалла, помещенного в насыщенный раствор. Деятельностью пластидул Геккель стремился объяснить размножение, наследственность, приспособление и эволюцию организмов...» [31, с. 31].
Аналогичную концепцию развивал в 1890 г. Альтман, полагавший, что протоплазма состоит из кристаллоподобных гранул; Бляхер пишет о его концепции: «...Альтман высказал предположение, что фибриллярные и сетчатые структуры, как и гомогенная по виду протоплазма, тоже построены из мельчайших гранул-биобластов, которые, по его мнению, являются элементарными единицами жизни. Альтман считал, что гранулы подобно клеткам размножаются делением, почему положение Вирхова «каждая клетка — от клетки» он заменил афоризмом «каждая гранула — от гранулы». Касаясь природы биобластов, Альтман сравнил их с кристаллами, которые по его представлению отличаются от неорганических кристаллов характером роста: неорганические кристаллы растут вследствие отложения вещества из маточного раствора на их поверхности..., тогда как биобласты питаются за счет проникновения надлежащих веществ внутрь» [31, с. 33].
В 1913 г. издательство «Образование» начало выпуск серии книг «Новые идеи в биологии». В первом сборнике этой серии, озаглавленном «Что такое жизнь», был помещен перевод статьи Г. Пржибрама «Обзор мнений авторов о значении аналогии между кристаллами и организмом».
Пржибрам пишет: «...Каждого биолога, углубившегося в кристаллографические изыскания, так сильно поражает «самодеятельность» кристаллов, что граница между неорганическим и органическим миром не кажется ему непреодолимой пропастью..».
Из сходства размножения с кристаллизацией следовали и более широкие биологические аналогии. А. И. Опарин подробно рассматривает эту аналогию в своей знаменитой книге, вышедшей в 1924 г. [227]. В связи со способностью организмов к воспроизведению он пишет[1]: «...указанное свойство присуще не только организмам, но всем без исключения телам, обладающим определенным строением... Возьмем кристалл какого-либо вещества... расколем его на две половинки и бросим последние в перенасыщенный раствор того же вещества. И вот оказывается, что брошенные в раствор половинки кристалла сравнительно быстро восполнят недостающие у них грани, углы, ребра».
А. И. Опарин приводит следующую цитату из книги Каруса Штерне, переведенной с 3-го немецкого издания на русский язык в 1909 г.: «...Здесь происходят самые изумительные явления, которые ...дают ключ к пониманию сложнейших явлений того же порядка. Сера, как известно, простое тело, смотря по температуре, при которой оно переходит из жидкого состояния в твердое, может принимать весьма различный вид — октаэдрическую или призматическую форму. Если опустить два таких кристалла на тонких платиновых проволоках в перенасыщенный раствор серы в бензоле, то по соседству с призматическим кристаллом начинаютобразовываться новые призмы, поблизости же от октаэдрической формы — октаэдры; когда обе армии кристаллов сблизятся между собой, то при первом столкновении последняя форма окажется побежденной. Вот пример борьбы за существование в царстве кристаллов»! [349, с. 200].
Таким образом Карус Штерне в конце прошлого — начале нашего века увидел не только аналогию размножения кристаллов и живых организмов — он обратил внимание на явления «борьбы за существование в царстве кристаллов». Это был очень важный шаг. Для перехода от физической химии к биологии необходимо от «борьбы за существование» кристаллов перейти к естественному отбору. Это становится возможным на основании созданной Н. К. Кольцовым концепции матричного воспроизведения биологических макромолекул [144], концепции конвариант- ной редубликации [288, 467, 468].
Г1о-видимому, именно гипотезы Н. К. Кольцова можно считать идейным началом современной молекулярной биологии. Сам Кольцов [144] утверждал, что мысль о матричном воспроизведении содержалась в докладе съезду естествоиспытателей и врачей, прочитанном А. А. Колли в 1893 г. [141, 142]. На докладе Колли присутствовал студент биологического факультета Московского университета Н. К- Кольцов, развивший потом эту мысль и доложивший свои соображения в 1927 г. на 3-м Всесоюзном съезде зоологов, анатомов, гистологов. У меня эта версия вызывает сомнение, поскольку при чтении текста доклада А. А. Колли мне не удалось усмотреть в нем даже намека на принцип матричного воспроизведения больших молекул. Единственное, что могло послужить толчком для Кольцова в докладе Колли — это вывод о малом числе молекул в бактериальной клетке. Отсюда следовало, что большое число признаков определяется малым числом молекул. Но как именно определяется — это додумал Н. К. Кольцов. Я не могу не привести слова, сказанные им в 1927 г. в своей речи на первом торжественном собрании 3-го Всесоюзного съезда зоологов, анатомов, гистологов в Ленинграде 12 декабря 1927 г.:
«...Процесс «ассимиляции», т. е. точного уподобления вновь возникающих из аминокислот белковых молекул тем образцам молекул, которые уже существуют в данном пункте, является одним из самых загадочных жизненных процессов.
Наиболее простой гипотезой для объяснения ассимиляции является, как мне кажется, заключение, что процесс синтеза белковых молекул сводится к кристаллизации вокруг уже существующих белковых молекул или их агрегатов — кристаллитов, являющихся затравками. Аминокислотные ионы прикладываются своими боковыми сродствами к тем пунктам уже существующих молекул, где находятся соответствующие аминокислоты, совершенно так же, как ионы Na+ и С1“, рассеянные в водном растворе, складываются е определенную решетку вокруг кристалликаповаренной соли... Много понадобилось времени, чтобы установить, что организм возникает только от другого организма из яйца Omne vivum ex ovo, Omnis cellula ex cellula, Omnis nucleus ex nucleo.
Теперь мы можем прибавить еще один новый тезис: каждая белковая молекула возникает в природе из белковой молекулы путем кристаллизации вокруг нее находящихся в растворе аминокислот и других белковых обломков: Omnis molecula ex mole- cula. Значит, размножение не есть исключительное свойство живых организмов, но является наиболее вероятным способом возникновения в природе всех сложных векториальных систем...» [144, с. 482].
Идеи Н. К- Кольцова, опубликованные в 20-е и 30-е годы, были развиты и распространены его учениками. Один из них — Н. В. Тимофеев-Ресовский оказал особенно существенное влияние «а мировоззрение тогда еще молодых физиков, обратившихся к теоретической биологии, например, на М. Дельбрюка {59, 83, 378]. В 1944 г. под впечатлением эгих идей Э. Шредингер написал свою знаменитую книгу «Что такое жизнь» [344]. Ученик Дельбрюка биолог Дж. Уотсон и физик Ф. Крик, занявшиеся биологией под влиянием книги Шредингера, на базе экспериментальных данных М. Уилкинса и Р. Франклин совершили одно из самых важных открытий биологии XX в. — установили структуру молекул ДНК (см. [297, 298]).
Я проследил кратко историю становления матричной концепции для того, чтобы, с одной стороны, показать преемственность идей нашего времени с идеями XVIII и XIX вв., а с другой — подчеркнуть давнее существование теоретической биологии.
Столь же глубоко, а быть может и еще глубже в историю науки уходят и корни эволюционной теории. Подчеркиваю я это потому, что в последнее время вновь стало «признаком хорошего тона» говорить о теоретической биологии в будущем времени как о дисциплине, которую еще лишь предстоит создать. Доля истины здесь, конечно, есть. Теоретическую биологию, как и любую другую теоретическую науку — теоретическую физику например, — отнюдь нельзя считать завершенной.
Однако она существует уже много лет и сейчас не стоит давать названия «На пути к теоретической биологии» книгам, посвященным развитию теоретической биологии (см., например, 1193,194,296]).
Необходимой и достаточной основой теоретической биологии является дарвинизм. Современная молекулярная биология позволяет дать физико-химическую интерпретацию дарвинизму.
Здесь необходимо напомнить, что долгое время совершались (и продолжаются ныне) попытки построить теоретическую биологию не на эволюционной основе, а посредством нахождения особых физических -или химических свойств живой материи. Фи-зико-химическая интерпретация биологических явлений, биохимический и биофизический анализы биологических объектов не имеют, ничего общего с таким неэволюционным подходом к теоретической биологии.
Целеустремленный поиск физического и (или) химического своеобразия биологических объектов был начат в XIX в. Когда прогресс физики и химии определил направление поиска «сущности жизни» в особых химических и физических свойствах веществ, составляющих основу «живой материи». Возникло убеждение, что жизнь — проявление особых свойств особого вещества.
После Лавуазье под влиянием его работ, показавших аналогию процессов брожения, дыхания и горения, распространилось мнение, что жизнь представляет собой особый химический процесс. Сам Лавуазье утверждал, что «жизнь — это химическая функция». По мнению Гегеля, «жизнь есть увековеченный химический процесс» (цит. по [352]). Такое особое, «наделенное жизнью» вещество получило многозначительное название «первичное», «протеин» (отнюдь не соответствующее современному понятию «белок»). Замечательным завершением этого этапа развития теоретической биологии стали широко известные формулировки Ф. Энгельса: «Жизнь — способ существования белковых тел...» [355, 356, с. 77].
Однако отождествление понятий «протеины» и «белковые тела», «протеины» и «белки» не соответствует смыслу определений Ф. Энгельса, что он и сам неоднократно отмечал. В конце XIX в. биохимики выделили из протоплазмы, не отличимой ранее от протеина, ряд существенных фракций. Одна из главных фракций протоплазмы и была названа собственно белком. Тогда же были начаты фундаментальные исследования Э. Фишера, показавшего, что белки — полипептиды, т. е. линейные полимеры, состоящие из аминокислотных остатков, соединенных пептидными
е,ВЯЗЯМИ.
И тут отчасти вследствие семантических неточностей, отчасти из-за стремления возможно быстрее понять сущность жизни, на белки была «возложена» общебиологическая «нагрузка» протеинов. Возникли гипотезы об обусловленности жизни особым составом, состоянием, свойствами молекул белка. Так возникло представление о «живом белке». Поскольку обмен веществ представлялся главным признаком жизни, были предприняты попытки найти причину обмена веществ в особых свойствах молекул белков. Предполагалось, в частности, особое неустойчивое, неравновесное состояние молекул живого белка, допускалось, что спонтанный распад этих молекул служит причиной обмена веществ — обновления молекулярного состава белковых макромолекул *.
Более чем столетний период развития науки в этом направлении завершился появлением книги Э. С. Бауэра «Теоретическая биология» [13]. Бауэр исходил из возможности определения всех основных свойств живых систем посредством анализа физических свойств молекул живой материи.
Согласно Бауэру, фундаментальное отличие живой материи от неживой характеризуется «принципом устойчивого равновесия». Этот принцип гласит: «Все и только живые системы никогда не бывают в равновесии и исполняют за счет своей свободной энергии постоянно работу против равновесия, требуемого законами физики и химии при существующих внешних условиях». Затем Бауэр в качестве следствий из этого принципа «выводит» основные проявления жизни — обмен веществ, рост, размножение. Бауэр, по-видимому, был неправ, постулируя, что устойчивое неравновесие возникает вследствие особого «напряженного» состояния белковых молекул. «Деформированное» состояние белковых молекул не является основным их свойством. Однако принцип Бауэра верен в эволюционном смысле, а именно, в смысле постоянно возрастающей в ходе эволюции «невероятности», а следовательно, и термодинамической неравновесности биологических макромолекул, «невероятности», проявляющейся в уникальности чередования мономеров в полимерных цепях белков и нуклеиновых кислот.
Сам опыт построения Бауэром теоретической биологии, его логика, конструкция созданной им теории, представляют большую ценность и в наше время. Однако как уже отмечено, его теоретическая биология основана на предположении особых физических свойств живой материи, а именно, особых термодинамических характеристик белков. Бауэр не объясняет эти свойства исходя из каких-либо общих соображений, а «формулирует» их посредством анализа отличий живых организмов от неживых объектов. В этом смысле его подход — неэволюционный. Он признает фундаментальное значение дарвинизма, но не применяет эволюционного подхода для построения теоретической биологии. В этом главный, на мой взгляд, недостаток концепции Бауэра.
Теоретическую биологию можно построить лишь при условии выяснения способа, путей эволюционного становления всех особенностей живых организмов, в том числе становления особенностей химических и физических свойств молекул, составляющих основу живых организмов.
Пути эволюционного становления всех особенностей живых организмов могут быть выявлены посредством применения теории Дарвина с учетом современных достижений генетики, биохимии, биофизики, объединенных в направлении, называемом молекулярной биологией.
Следует отметить, однако, что в последнее время вновь, после Длительного перерыва стали высказываться сомнения в верности дарвинизма (см. [206]). А. А. Любищев классифицировалразличные теории эволюции, полагая при этом дарвинизм лишь одной из нескольких возможных теорий эволюции [185в, г].
С одной стороны, делаются попытки противопоставления теории эволюции в результате естественного отбора случайных мутаций и теории эволюции как закономерного развертывания «внутренних» свойств биологических систем (номогенез Л. С. Берга [21]). Детальное обсуждение этих противопоставлений выходит за пределы задач этой книги (см. [118, 105—107]). Однако противопоставление эволюции в результате естественного отбора и эволюции, как реализации определенных закономерностей совершенно неверно. Закономерность эволюционных траекторий — обязательное следствие, а не противоречие дарвинизму. Поэтому, в частности, неверно рассмотрение закона гомологических рядов Н. И. Вавилова, как противоречие случайной изменчивости и эволюции в результате естественного отбора [21, 206]. Гомологические ряды Н. И. Вавилова лишь подтверждают, в соответствии с теорией Дарвина, закономерный характер эволюции [43, 44, 107]. Гомологичные результаты естественного отбора отнюдь не являются проявлением неслучайного характера изменчивости, а лишь свидетельствуют о действии гомологичных (и даже одинаковых) критериев естественного отбора.
Таким образом, в результате естественного отбора случайных изменений, мутаций, происходит закономерная эволюция. Эволюционные траектории детерминированы, определены физикохимическими и собственно биологическими факторами — это основное содержание настоящей книги. Нет никаких оснований поэтому противопоставлять теорию Дарвина теориям закономерной эволюции. Но при этом речь идет не о таинственных «внутренне присущих системе» законах, а о физических, химических, биологических, доступных рациональному анализу закономерностях.
С другой стороны, высказываются сомнения в какой-либо закономерности эволюции, полагая ее траектории случайным, не зависящим от естественного отбора результатом появления, накопления, комбинации «нейтральных» мутаций [411—414]. «Нейтралистская» концепция возникла при анализе мутационных замен в полипептидных (полинуклеотидных) последовательностях биополимеров [377]. Некоторая часть таких замен не приводит к регистрируемым изменениям функциональных свойств макромолекул. Однако отсюда не следует, что эти нейтральные мутации определяют направления эволюции. Прекрасный анализ «нейтралистской теории эволюции» дан Р. Левонтином. Он пишет: «Никто не настаивает на том, что почти все мутации нейтральны или что эволюция протекает без естественного отбора, главным образом путем случайного закрепления нейтральных мутаций. Оба эти утверждения явно неверны и совершенно чужды духу предлагаемых объяснений. Напротив, мы настаиваем, что многие мутации испытывают действие естественного отбора, но почти всеэти мутации вредны и элиминируются их популяции. Второй распространенный класс представлен группой нейтральных мутаций... Кроме того, рассматриваемая теория допускает существование редких благоприятных мутаций, которые закрепляются естественным отбором, поскольку адаптивная эволюция все же происходит» [173, с. 202].
Из сказанного не следует отрицания физического и химического своеобразия биологических систем. Можно лишь, не претендуя на оригинальность, подчеркнуть, что жизнь, живые существа — результат длительного эволюционного процесса. Живые существа, биологические системы — концентрат многих миллиардов лет эволюции. Биология — наука историческая. Невозможно ни выявить, ни, тем более, понять основные свойства биологических систем вне эволюционного подхода. Только такой подход позволит определить ведущие, характерные признаки живых существ и тем самым позволит определить предмет биологии в целом и каждой ее области в отдельности.
Весь процесс биологической эволюции оказывается возможным подразделить на ряд этапов, отличающихся друг от друга видом, качеством ведущих факторов эволюции. Переход от одного фактора эволюции к качественно другому сопряжен с «кризисом», преодоление которого приводит к резкому ускорению преобразования веществ неживой природы в вещества живых организмов.
Диалектический характер процесса биологической эволюции как процесса, идущего от одного качественного этапа (аромор- фоза) к другому, с преодолением при переходе от этапа к этапу кризисов и противоречий стал ясен из работ А. Н. Северцова [255, 256]. Каждый крупный ароморфоз существенно увеличивает темпы прогрессивной эволюции. В принципе каждый ароморфоз можно количественно охарактеризовать, например, увеличением интенсивности преобразования вещества и энергии у организмов, носителей этого ароморфоза, по сравнению с их предками. Ход процесса эволюции от одного крупного ароморфоза к другому можно рационально описать на основании общих физико-химических соображений. Речь идет в данном случае не о темпах перехода от одного ароморфоза к другому, а лишь о дедуктивном описании возможной последовательности ароморфозов при эволюции в данных условиях.
В некотором смысле такой подход аналогичен термодинамическому, при котором существенны лишь начальные и конечные состояния системы. Аналогию с термодинамикой при анализе процесса биологической эволюции нетрудно продолжить. В самом деле, огромное число микроскопических компонент системы (атомов, молекул), не поддающееся описанию разнообразие траекторий движения и взаимодействий друг с другом этих микро- компонент не препятствие, а условие термодинамического, макроскопического подхода, описания состояний макросистемы. Анало-
тичным образом, огромное число и разнообразие объектов биологической эволюции, чрезвычайная сложность их взаимодействий — непрепятствие, а условие описания узловых макроэтапов (ароморфозов) процесса биологической эволюции. На каждом этапе некий существенный в естественном отборе признак, свойство достигает предельного совершенства. Физико-химический и биологический смысл и содержание этого совершенства можно проанализировать без учета времени, необходимого для его достижения (принцип предельного совершенства при анализе эволюционного процесса).
Ясно, однако, что применение принципа предельного совершенства при анализе эволюционного процесса — лишь удобный, а в некоторых случаях и единственно возможный способ первого приближения к действительной картине.
Достаточно полное представление о биологической эволюции может дать рассмотрение не только основных этапов, предельно совершенных по каким-либо факторам, но и темпов эволюции. Далее делается попытка рассмотрения закономерностей, определяющих темпы эволюции, скорость возрастания «биологического прогресса».
Традиционные определения понятий «жизнь, живое существо» были основаны на перечислениях характерных свойств живых организмов. Однако в большинстве случаев удавалось каждое характерное свойство найти в заведомо неживых объектах. Анализ этой ситуации привел к распространенному ныне скептическому отношению к самой возможности таких определений, к убеждению в «бессмысленности» и бесплодности таких попыток {446, 429]. Эту ситуацию четко анализировал Э. Бауэр [13], предложивший, как отмечено выше, в качестве определяющего живое состояние «принцип устойчивого неравновесия».
Затруднения в формулировании необходимых строгих определений снимаются при учете эволюционного происхождения живых организмов: их перечисляемые обычно характерные свойства возникают, формируются в ходе эволюции. В соответствии с этим жизнь — это процесс существования объектов биологической эволюции. Живые существа (организмы) — объекты биологической эволюции [339, 340]. Свойства и характерные черты жизни и жизни существ (организмов) различны на разных стадиях биологической эволюции. Следовательно, единственное, что остается определить, это понятие «биологическая эволюция». Это сделано в следующей главе.
Итогом проведенного далее анализа, по нашему мнению, является доказательство возможности дедуктивного описания важнейших биологических закономерностей и свойств живых организмов посредством рассмотрения процесса эволюции в данных химических и физических условиях. Это значит, что основные этапы биологической эволюции детерминированы физическими и ^химическими факторами окружающей среды.
Глава 2
КИНЕТИЧЕСКИЕ КРИТЕРИИ ЕСТЕСТВЕННОГО ОТБОРА И ФИЗИКО-ХИМИЧЕСКИЕ И БИОЛОГИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ
Осуществление естественного отбора по признаку кинетического совершенства в системе, способной к полиморфной кристаллизации (матричным синтезам). Принципиальная возможность анализа наиболее вероятных путей биологической эволюции на основе оценки величин кинетического, биологического совершенства и эволюционного потенциала.
Для того чтобы пояснить смысл понятия кинетического совершенства, продолжим аналогию между размножением и борьбой за существование живых организмов и кристаллов.
Представим себе, что некая «жидкость» может с одинаковой вероятностью кристаллизоваться в различных кристаллических модификациях, т. е. имеется система, способная к полиморфной кристаллизации.
В нашем абстрактном случае любая возможная кристаллическая форма имеет одну и ту же вероятность появиться в данных условиях. Отсюда следует, что в этих условиях термодинамическая «выгодность» любой формы одинакова.
В достаточно большом, хотя и ограниченном пространстве, судьба системы, вид, форма кристаллов, заполняющих это пространство, определяется, следовательно, не термодинамическими, а кинетическими особенностями разных кристаллических форм: пространство сначала заполнится формой, которая образуется по случайно возникшей затравке быстрее, чем остальные. Затем по прошествии достаточно долгого времени исследуемое пространство окажется занятым всеми кристаллическими формами в равной доле (поскольку термодинамические характеристики всех форм одинаковы). Представим себе теперь, что более «быстрая», т. е. быстрее заполняющая пространство форма, менее стабильна. И в этом случае такая форма некоторое время может «господствовать» в системе, заполняя большую часть ее объема. Теперь допустим (что вполне реально), что у более быстрой, или в общем случае, у какой-то одной из кристаллических форм расположение молекулярных групп на гранях, сторонах кристалла обусловливает их каталитические свойства, которые позволяют кристаллам данной формы ускорять некую химическую реакцию, сопровождающуюся выделением энергии и способствующую процессу кристаллизации *.
Тогда эта кинетически более совершенная, хотя термодинамически и нестабильная форма за счет свободной энергии, выделяющейся из сопряженно катализируемой реакции, будет все более заметно преобладать над другими формами. Теперь сделаем следующий важный шаг — допустим, что наша система, способная к полиморфной кристаллизации, находится в потоке излучения или каких-то иных внешних воздействий, которые могут стимулировать образование новых вариантов кристаллов, увеличивать разнообразие реализованных форм.
Среди таких измененных, мутантных форм могут оказаться и кинетически более совершенные, т. е, способные к более быстрой кристаллизации и (или) к катализу экзэргонических сопряженных реакций. Ясно, что такие мутантные формы имеют «шансы» заполнить большую часть объема. При этом новом условии — изменчивости — наша система будет развиваться в направлении отбора кинетически все более совершенных форм.
Из изложенных выше соображений следует, что при возможности полиморфной кристаллизации, при наличии разных кинетических (в том числе каталитических) свойств у разных кристаллических форм, при возможности изменчивости кристаллических форм в результате тепловых флуктуаций юли иных причин (мутации), осуществляется естественный (!) отбор форм, способных ко все более быстрому размножению, т. е. к кристаллизации по затравке данного вида.
В результате происходит развитие, смена форм в направлении все большего кинетического совершенства. Эти все более совершенные формы могут (и должны быть) термодинамически все менее вероятными и все более сложными.
Я все время подчеркиваю определяющую роль кинетических факторов в процессе эволюции. Ясно, однако, что любая изолированная система будет следовать термодинамическим законам[2]. И арена эволюции данной системы, ее ареал будет заполнен в конце концов не наиболее совершенными кинетически, а наиболее вероятными, и, следовательно, наиболее термодинамически устойчивыми формами.
Однако в открытой системе при наличии внешнего источника свободной энергии процесс образования и роста матриц может осуществляться сопряженно, т. е. за счет свободной энергии поступающей извне, например, в результате сопряженных химических экзэргонических реакций (см. гл. 7). В таком случае в понятие кинетическое совершенство входит и эффективность сопряжения процесса возникновения и роста матриц (кристаллов) с экзэргоническими реакциями и само «качество» этих реакций. В подобных условиях направление процесса эволюции будут определять не термодинамические, а кинетические свойства кристаллов. Мало того, поскольку отбор будет идти по признаку кинетического совершенства (самого по себе мало вероятного), побеждать в нем будут все менее вероятные формы, будет осуществляться выбор удачных флуктуаций, мутаций. Каждый шаг такой эволюции может быть достаточно вероятным, итог многих последовательных шагов, если забыть об истории, о способе достижения данного этапа, будет казаться совершенно невероятным. Многие исследователи оценивали вероятность возникновения определенной последовательности, например, нуклеотидов в ДНК, и получали величину порядка 10~500. Однако такие внеэволюционные оценки нисколько не противоречат высокой вероятности эволюционного возникновения сложных форм жизни [23, 212, 232].
Действительно, вероятность того, что в данный момент существует какая-либо форма, равна 1. Этого достаточно, чтобы начался естественный отбор (при выполнении указанных выше условий).
Таким образом, естественный отбор начинается с неизбежностью, и вопрос состоит в том, как быстро будет возрастать кинетическое совершенство в эволюционирующей системе. Нужны не оценки вероятности возникновения данной формы (данной последовательности нуклеотидов) молекулы ДНК, а оценки скорости лроцесса эволюции, выяснение того, достаточно ли прошедшего времени для достижения данной «величины» биологического прогресса [264]. Естественный отбор полезных флуктуаций в открытой термодинамической системе с матричным воспроизведением направляет процесс эволюции в сторону, противоположную предписаниям термодинамики закрытых систем. Термодинамика отнюдь не нарушается: процесс эволюции «окупается» сопряженными экзэргоническими процессами. Но направление эволюции определяется кинетическими, а не термодинамическими факторами. Мало того, не просто осуществляется процесс эволюции в направлении все менее термодинамически вероятных форм. Сам лроцесс отбора идет со все большей (до некоторого предела) •скоростью — система не просто удаляется от термодинамического равновесия, а удаляется от него все дальше и с возрастающей скоростью, так как в естественном отборе побеждают более совершенные формы, возникающие быстрее, раньше других. В этом отличие эволюционирующей системы от неэволюционирующей термодинамической системы, где в соответствии с теоремой При- гожина, скорость удаления от равновесия, скорость приращения энтропии минимальна. Естественный отбор, процесс эволюции в Силу давления отбора «заставляет» объекты эволюции с предельно возможной скоростью удаляться от положения термодинамического равновесия, отнюдь не нарушая законов термодинамики. Происходит это за счет использования свободной энергии сопряженных экзэргонических реакций и в результате «ограждения» термодинамически маловероятных структур большими кинетическими (потенциальными) барьерами.
Итак, основной чертой процесса биологической эволюции является определяющая роль, доминирование кинетических, а не термодинамических закономерностей. Это «противотермодинами- ческое» развитие системы в направлении все большего кинетического совершенства осуществляется при возможности использования свободной энергии, освобождающейся при каком-либо экзэргоническом процессе. «Очевидный» механизм эволюционного кинетического совершенствования — случайная или закономерная изменчивость объектов эволюции, сохранение кинетически более совершенных и несохранение менее совершенных вариантов в ходе естественного отбора. Сказанное о естественном отборе в системе с полиморфной кристаллизацией верно для кристаллов любой природы. Однако абсолютная величина интенсивности кристаллизации, предельно возможное кинетическое совершенство зависят от природы кристаллов. Кинетические свойства кристаллов определяются природой элементов кристаллической решетки. Скорость кристаллизации, каталитические свойства поверхности кристаллов зависят от физико-химического разнообразия этих элементов и от величины удельной поверхности кристаллов, поскольку матричный механизм основан в принципе на поверхностном взаимодействии.
Важно подчеркнуть, что кинетические свойства становятся критерием естественного отбора в определенных условиях: при ограниченности «реакционного объема» (возможного ареала), при ограниченности количества вещества, могущего быть превращенным в вещества данного вида, при ограниченности интенсивности потока энергии, поступающей в эволюционирующую систему.
В этих условиях кинетическое совершенство /< определяет скорость преобразования веществ среды в вещества i-ro вида посредством матричных механизмов и задается формулой:
It == rhi = dmjdt = Кти
где nii — масса вещества t-ro вида.
Для характеристики направлений процесса эволюции полезно понятие эволюционного потенциала, определяемого как предельное для данного направления кинетическое совершенство 1тг1.
Эволюция в данном /-м направлении идет до тех пор, пока кинетическое совершенство 1[ не сравняется с /4ах- В таком состоянии исчерпания потенциала система может находиться неопределенное время, пока случайные мутации не «толкнут» ее на новое эволюционное направление. Естественный отбор с высокойскоростью будет усиливать казалось бы ничтожные различия в кинетическом совершенстве разных форм.
Представим себе, сильно упрощая картину, два вида А и В ■(кристаллов, матричных молекул, организмов) скорость размножения которых различна. Пусть они, размножаясь, заполняют предельный объем (ареал), вмещающий ограниченное количество «живого вещества» любого вида. При заполнении этого объема масса вещества разных видов будет пропорциональна соответствующим величинам Представим себе теперь, что ареал заполняется организмами двух видов А и В, затем они живут некоторое время, стареют и умирают, оставляя потомство — зародыши, незначительные по массе, численность которых пропорциональна массе родителей. Жизненный цикл от зародышей до смерти назовем поколением. Легко показать, что соотношение масс организмов видов А и В при 1в=1А-\-г через N поколений будет равно: mA/mB=mAJmBoeNl1. Можно оценить, через сколько поколений N при различных величинах е масса организмов вида А превысит массу организмов вида В в определенное число раз а. Приняв mAJmB = 1, получим:
N = \n(mA/mB) /е.
Эти оценки для наглядности представлены в табл. 1.
Таким образом, 1%-ное различие в величинах е уже через 460 поколений приведет к 100-кратному превышению массы вида А над массой вида В (а=102).
Приведенные оценки не учитывают случайных изменений среды. Флуктуации условий существования могут затруднить или даже сделать невозможным «выход на экспоненту» раз-
таблица 1
ЧИСЛО ПОКОЛЕНИЯ, НЕОБХОДИМОЕ ДЛЯ ПРЕВЫШЕНИЯ МАССЫ ВИДА А МАССОЙ ВИДА В в а РАЗ ПРИ РАЗНЫХ ЗНАЧЕНИЯХ е
е
а
Ю
10*
10»
10*
10»
0,1
23
46
69
92
115
0,01
230
4С0
690
920
1150
0,001
2300
4600
6900
9200
1150
множения особей — обладателей малых селективных преимуществ, когда собственно и начинается естественный отбор. Вопрос об условиях действия естественного отбора во флуктуирующей среде очень сложен. Ему, в частности, посвящены работы Ю. В. Чайковского [319, 321]. Общие закономерности эволюционного процесса с учетом колебания численности популяций, дрейфа генов, флуктуаций, условий существованиярассмотрены в ряде специальных книг ,и статей [57, 66, 80, 82, 105, 106, 108, 122, 127, 173, 189, 202, 203, 213, 235, 241, 243, 264, 289, 290, 308, 316, 332, 333, 326, 327, 375, 387, 351, 412, 468].
Таким образом, приведенные выше численные оценки — лишь иллюстрация высокой эффективности естественного отбора после «выхода на экспоненту», когда небольшие различия в итоговой интенсивности размножения — в кинетическом совершенстве — учитываются естественным отбором.
Является ли кинетическое совершенство действительно универсальным критерием естественного отбора в многосложных процессах биологической эволюции?
Нельзя ли в таком случае «приравнять» термины кинетическое совершенство и биологическое совершенство?
Возможны ли измерения этой величины не только для полиморфной кристаллизации, но и для дальнейших этапов биологической эволюции?
Попробуем ответить на эти вопросы. Действительно, при полиморфной кристаллизации в идеализированных условиях критерием отбора одной из многих равновероятных кристаллических форм является скорость кристаллизации. Чтобы измерить кинетическое совершенство разных кристаллических форм, следует измерить скорость заполнения ими данного объема (ареала) при прочих равных условиях. Процедура ее измерения должна состоять во внесении затравок соответствующих кристаллических форм в способную к кристаллизации жидкость и в определении скорости кристаллизации по данной затравке.
Однако в биологических системах сравниваемые формы могут очень сильно различаться. Можно ли количественно сопоставлять кинетическое совершенство бактерии и слона? Очевидно, нет. Формы, способные строить себя за счет разных исходных веществ, не конкурируют друг с другом за материал. Забравшись в недоступную другим экологическую нишу, некий вид может на более или менее длительное время выйти из-под давления естественного отбора и перерабатывать вещества среды в вещества своего вида значительно медленнее, чем виды, принадлежащие другим направлениям эволюции. Таким образом, Ii служит критерием отбора лишь при прочих равных условиях, при одинаковых условиях существования. Эти прочие равные условия соблюдаются только в момент возникновения новых форм, в самом начале дивергенции, в самом начале вытеснения менее совершенного более совершенным. Дивергентная эволюция приводит к заполнению под давлением естественного отбора всех возможных для данного направления эволюции экологических ниш. В каждой нише рано или поздно окажется вид с предельно возможным в данных условиях кинетическим совершенством. Заполнение всех возможных экологических, ниш А. Н. Северцов [255] назвал идиоадатацией.
Однако в процессе естественного отбора могут возникнуть новые «эволюционные идеи», новые принципы жизнедеятельности, допускающие новую серию эволюционных приспособлений, новую волну заполнения всего разнообразия экологических ниш (в том числе и заполненных ранее). Такие новые принципы приспособления, резкие увеличения эволюционного потенциала Северцов назвал ароморфозами. Старые обитатели различных экологических ниш будут вытесняться новыми (как были вытеснены некогда рептилии млекопитающими, папоротники — цветковыми растениями и т. п.) и их вытеснение будет осуществляться снова по критерию /(— в каждой экологической нише будет разыгрываться своя драма: старый хозяин должен будет сойти со сцены, уступить свое место новому [391].
Следовательно, критерий /< действует при заполнении данной экологической ниши в ходе идиоадаптации, все более совершенного приспособления к жизни в данных условиях, т. е. все большей специализации на базе общего ароморфоза и при вытеснении организмов низшего эволюционного уровня представителями более высокого уровня при появлении нового ароморфоза.
Итак, при прочих равных условиях величина /,• является абсолютной мерой биологического совершенства данного вида. Биологическое совершенство организмов, живущих в разных условиях, можно в принципе сопоставлять лишь с учетом особенностей данных условий существования и внесением соответствующих коррекций в наблюдаемую величину U. Такая коррекция представляет собой в высшей степени трудную задачу даже в случае организмов, занимающих сходные экологические ниши. Поэтому вычисление корректированных, приведенных к нормальным условиям величин позволяющих сравнивать биологическое совершенство бактерий и слонов практически не реально. Это не изменяет того обстоятельства, что величины /,-, приведенные к нормальным условиям, в принципе количественно характеризуют биологическое совершенство данного вида (и вообще, таксономических единиц любого ранга).
Как уже отмечено, Северцову принадлежит открытие общего характера эволюционного процесса. Он писал: «Биологический 'прогресс, т. е. выживание и победа в борьбе за существование, достигается четырьмя различными способами, а именно: 1) путем ароморфозов, 2) путем идиоадаптаций, 3) путем ценогенезов и 4) путем общей дегенерации» [255, с. 100]. Эти способы Северцов определял следующим образом: морфофизиологический прогресс, или ароморфозы — приспособительные изменения, при которых общая энергия жизнедеятельности взрослых потомков повышается; идиоадаптации — приспособительные изменения, при которых энергия жизнедеятельности взрослых потомков не повышается, но и не понижается; ценогенезы — приспособительные изменения зародышей (и личинок), при которых общая энергия жизнедеятельности и строение взрослых потомков не
РИС. 1. Зависимость морфофизио- логического прогресса, общей энергии жизнедеятельности, от времени в ходе эволюции (по А. Н. Се- верцову [256])
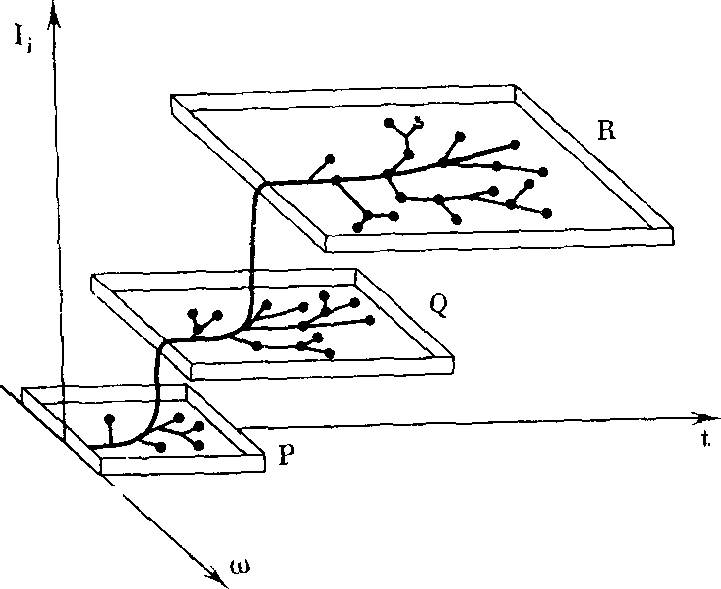
Морфофизиологический прогресс

Биологический прогресс может сопровождаться морфофизиологическим ре* грессом (спуск на плоскость Р) ил» морфофизиологическим прогрессом, аро- морфозом (подъем на плоскости Q и- R). В каждой плоскости происходят приспособления к различным условиям, существования (ось со)— идиоадаптации без изменения общей энергии жизнедеятельности; t — время
РИС. 2. Видоизмененный график Северцова, иллюстрирующий возрастание величины кинетического (биологического) совершенства /> в ходе эволюции
Резкие подъемы, ароморфозы, сменяются приспособлениями к разным условиям существования (ось (о) — идио- адаптациями, в ходе которых также наблюдается некоторый рост /,
РИС. 3. Возрастание кинетического- (биологического) совершенства А организмов, приспособленных к разным условиям существования (ось <о) в ходе эволюции
Жирные горизонтальные отрезки — уровни исходных ароморфозов (Р, Q, R), тонкие горизонтальные — уровни / при идиоадаптациях. Вертикальные линии —- переходы от одного ароморфоза к другому. Существование видов с более высоким I исключает существование в тех же условиях (экологических нишах) менее совершенных видов
РИС. 4. Возрастание кинетического совершенства в ходе эволюции при нормировке всех условий существования

изменяются, но возрастает число потомков; общая дегенерация — приспособительные изменения взрослых потомков, при которых общая энергия жизнедеятельности понижается [255, с. 60].
Ход эволюционного процесса Северцов иллюстрировал ставшим широко известным рисунком (рис. 1). В горизонтальных плоскостях происходят идиоадаптационные приспособления к различным условиям существования. Подъемы с одной плоскости на другую представляют ароморфозы, спуски — дегенерацию. Недостатком приведенной схемы является отсутствие строго определенного смысла понятия «морфофизиологический прогресс», «общая энергия жизнедеятельности».
При характеристике хода эволюционного процесса величиной кинетического (биологического) совершенства h схема Север- цова предстает в измененном виде (рис. 2). Эволюция неуклонно идет в направлении возрастания /Биологический регресс (в отличие от морфофизиологического) невозможен. Общая дегенерация (например, у паразитов) становится в этих координатах одним из способов идиоадаптации.
Если для удобства сравнения абсолютных величин кинетического совершенства в одинаковых условиях пренебречь временем, необходимым для достижения данной величины /(, то получится график (рис. 3). При помощи этого рисунка можно сравнивать кинетическое совершенство организмов, последовательно занимавших одинаковые экологические ниши в процессе эволюции. На рис. 3 видно, что новые ароморфозы сопровождаются резким увеличением кинетического совершенства. После каждого ароморфоза происходят относительно небольшие увеличения /,• в ходе идиоадаптаций (сверхтонкая структура спектра /().
Наконец, при нормировке, приведении всех разнообразных условий существования к нормальным, мы получим график (рис. 4). Это безусловно предельное упрощение.
Ясно сознавая сложность такой нормировки, я вынужден буду анализировать далее основные этапы биологической эволюции, имея в виду именно такую предельно идеализированную схему эволюционного процесса.
Сейчас, конечно, нереально пытаться проследить из общих физико-химических соображений путь и темпы биологической эволюции. Однако анализ узловых этапов, основных ароморфо- зов, представляется мне осуществимым.
Появление ароморфоза означает введение в число механизмов кинетического совершенствования очередного нового принципа. Всякий новый ароморфоз становится необходимым по исчерпании возможностей, т. е. эволюционного потенциала, предыдущего.
Эффективность ароморфозов можно в принципе оценить из общих соображений,— посредством вычисления эволюционного потенциала того или иного физико-химического фактора биологической эволюции. К таким факторам относится весь ряд физико-химических механизмов, определяющих кинетическое совершенство. Так, предельная скорость синтеза лимитируется каталитической активностью некоторых компонент системы, скоростью притока и оттока веществ, эффективностью использования свободной энергии в сопряженных экзэргонических процессах, температурой среды и пр., скорость распада матричных молекул зависит от интенсивности внешних излучений,, температуры среды, наличия специальных замедляющих распад приспособлений и т. д.
Все перечисленные выше физико-химические механизмы можно проанализировать, исходя из общих соображений. Следовательно, можно определить (вычислить) и величину эволюционного потенциала каждого очередного этапа, ароморфоза в биологической эволюции.
Я пока не рискую, однако, проводить количественные оценки. В последующих главах я надеюсь показать возможность качественной оценки эволюционного потенциала различных очередных узловых этапов эволюционного прогресса, а также строгую детерминированность самой последовательности основных ароморфозов в эволюции.
Прослеживая динамику событий в абстрактной системе с полиморфной кристаллизацией, способной к конвариантному воспроизведению, мы пришли к выводу, что в ней происходит естественный отбор по признаку кинетического совершенства. В результате этого некая часть системы, та, в которой воплощаются результаты естественного отбора, развивается, становится все более совершенной (и, следовательно, сложной).
Процесс такого развития мы будем называть эволюцией. Эволюцию, идущую в направлении все большего кинетического совершенства, мы будем называть далее биологической эволюцией.
В общем случае не существенна физико-химическая природа веществ, способных к конвариантной редубликации, приводящей к биологической эволюции. Однако среди многих в принципе возможных систем конвариантной (матричной) редубликацииесть лишь одна в принципе предельно совершенная, обладающая максимальным эволюционным потенциалом. По-видимому, такой системой являются нуклеиновые кислоты и белки. Но в данном контексте это не важно. Независимо от конкретной физико-химической природы конвариантно воспроизводящихся матричных молекул, процесс увеличения их кинетического совершенства в результате естественного отбора есть биологическая эволюция.
УВЕЛИЧЕНИЕ БИОЛОГИЧЕСКОГО (КИНЕТИЧЕСКОГО) СОВЕРШЕНСТВА, НАПРАВЛЕНИЕ И СКОРОСТЬ ЭВОЛЮЦИОННОГО ПРОЦЕССА. ПРИНЦИП ДОМИНАНТЫ
Итак, в конкуренции за вещество, энергию и пространство из нескольких «мутантов» побеждает тот, у которого итоговая скорость заполнения возможного ареала выше. Этот, в сущности, кинетический критерий отбора /,• имеет размерность г-сек-1. В самом деле существенна не численность, а количество вещества, преобразуемого в единицу времени в вещества данного варианта (мутанта, вида).
Приведенная выше (стр. 22) зависимость тi от К и т,- основана на кинетике автокатализа, естественной для систем с матричным (типа кристаллизации) механизмом размножения. Величина /< определяется величиной К, являющейся функцией многих переменных величин. В простейшем случае /С,= ЯКц. В частности, К определяется скоростью синтеза мономеров- компонентов матричных макромолекул, скоростью полимеризации уже имеющихся мономеров с образованием макромолекул, эффективностью использования поступающей в систему энергии, устойчивостью образовавшихся макромолекул (организмов) по отношению к неблагоприятным условиям среды и т. д. Соответственно этому величина h является суммой /у, где каждая /13- равна вкладу в итоговую скорость заполнения пространства, т. е. скорости, обусловленной отбором по данному критерию направлению эволюции Kj, т. е.:
Ki = ^ К и и /,■ = VKijtrii. i
Эволюция все время идет в направлении наиболее быстрого возрастания эволюционирующие системы, медленно заполняющие данное «пространство возможностей» будут вытесняться системами, заполняющими это пространство быстрее. Таким образом анализ направлений процесса эволюции сводится к исследованию закономерностей, определяющих скорость возрастания «кинетического» или, что то же самое, «биологического» совершенства U.
Эволюция идет под давлением естественного отбора. Чем больше это давление в данном направлении, тем с большей
вероятностью и скоростью по нему пойдет траектория эволюционного процесса. Замечательно, что повышение скорости движения по данной эволюционной траектории — более быстрое увеличение /; по /-му направлению — означает (в силу конкуренции) одновременное замедление эволюции по другим направлениям. Происходит активное доминирование (аналогично принципу доминанты А. А. Ухтомского применительно к нервной системе [299]) данного направления над другими. В самом деле, носители небольших преимуществ по другим направлениям будут вытесняться в естественном отборе носителями положительных изменений в доминантном направлении, поскольку селективное преимущество оценивается не по вкладу отдельных направлений, а по суммарной величине /4= 2/^.
В то же время давление отбора тем больше, чем ближе по своим свойствам конкурирующие формы. Следовательно, по мере все большего доминирования данной эволюционной траектории, все более узко направленного эволюционного совершенствования скорость возрастания /,■ будет возрастать. Эволюция в данном /-м направлении не означает отсутствия дивергенции. Однако эта дивергенция свойств лишь по данному Кц±Ьц направлению отбора, где ±6* характеризует вариации Кц в данных условиях. По ходу эволюции происходит все более детальная дифферен- цировка этого очередного направления —«разрешающая сила» критерия все время растет. Итак, однажды начавшись, движение по данной эволюционной траектории, т. е. возрастание /« (по К, критерию) идет с возрастающей скоростью, пропорциональной уже достигнутому уровню Л= 2/3- и обратно пропорциональной широте }-го критерия отбора, т. е. /С,±6. Эволюционное совершенствование, т. е. рост А в данном направлении, т. е. за счет роста /з идет до максимально возможного уровня /,• jmax, ограничиваемого физическими, физико-химическими или чисто биологическими причинами. При приближении I{j к IijmdX, среднее приращение АI в результате мутаций становится сравнимым с /шах—Аз, темп эволюционного совершенствования уменьшается и падает до нуля при 1ц=1ц max- Дальнейшее эволюционное совершенствование становится возможным теперь лишь при увеличении /( за счет возрастания по другому направлению — по новому критерию отбора. В силу уже сказанного эволюционное совершенствование по новому направлению сначала идет медленно, а потом «разгоняется». Ход во времени эволюционного совершенствования по каждому отдельному Kj должен иметь вид графика (рис. 5), а при переходе к другому направлению {Kj-О общий вид процесса эволюции во времени также должен иметь вид графика (см. рис. 5).
Вклад в суммарную величину /,• отдельных Itj может быть резко разным, и ход эволюционного возрастания /, представлен на рис. 5.

РИС. 5. Кинетика возрастания Л под давлением естественного оюора в j-u направлении
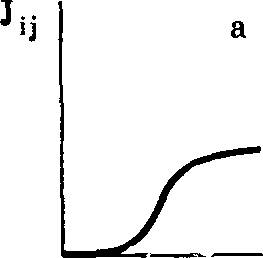
о—«лаг-фаза», фаза «разгона», фаза «замедления темпа роста по /-му напраилеиню. Длительность лаг-фазы определяется временем, необходимым для установления доминирования /-го направления над другими возможными направлениями эволюции. Фаза разгона— возрастание скорости эволюции по мере сужения ее направления при доминировании. Фаза замедления обусловлена исчерпанием эволюционного потенциала данного направления; б — возрастание 1{ последовательно при эволюции по /-му и (/ + 1)-му направлениям; в — последовательная эволюция по трем направлениям. Показано, что величина приращения I^ при этом может быть различной
Описанная кинетика эволюционного процесса соответствует уравнению:
d/(. ■ _ _ /(Д/,(<)
— 11 — ftii —
it 1 + Б6/(.
В начале движения по данной эволюционной траектории разных мутантов велико. Давление отбора мало. Скорость изменения , мала. По мере продвижения по данной траектории 263ч уменьшается. /f растет (т. е. скорость приращения mt растет). Однако при приближении к тах начинает убывать Д/ (стремящаяся к 0), и эволюция замедляется.
Очередность преимущественного движения эволюционирующей системы в направлении то одного, то другого критерия отбора Kj в принципе может быть установлена из общих соображений [339, 340].
ЭВОЛЮЦИОННО-ТАКСОНОМИЧЕСКИЕ ДИАГРАММЫ
Можно представить себе эволюционное возрастание U как движение по радиусу-вектору в Kj-м направлении (К} задается дугой окружности, измеряемой в радианах, отсчитываемой от направления эволюции в начальный момент времени). Каждому Kj соответствует некоторый диапазон К,+6. Пусть, для определенности, поочередно вступающие в действие критерии отбора (факторы эволюции) располагаются друг за другом на окружности при движении по часовой стрелке. Тогда при данных усилиях процесс эволюции может быть представлен серией «эволюционно-таксономических диаграмм» (рис. 6). Процесс эволюционного совершенствования по данному критерию- состоит в движении по радиусу-вектору в секторе /С,±6. Мерой

РИС. 6. Эволюционно-таксономические диаграммы — изменения во времени Kj — факторы, определяющие направления эволюции (т. е. увеличение 1$, расположены по окружности, по часовой стрелке и измеряются отрезком дуги окружности в радианах. Кинетическое (биологическое) совершенство измеряется суммарной площадью диаграммы; а, б, в, г, д — различные этапы эволюции

РИС. 7. Эволюционно-таксономические диаграммы трех (абстрактных) близких таксонов (видов, родов и т. п.)
эволюционного совершенствования в /-м направлении является площадь соответствующего сектора. Полной мерой биологического (кинетического) совершенства является суммарная площадь всех заполненных секторов (заштрихованных на диаграммах). По завершении эволюции по всем направлениям достигаютсямаксимально возможные в данных условиях величины площади соответствующих секторов. (Эти предельные значения могут быть, как уже отмечалось, резко различными для разных Кц).
В таком состоянии темп эволюции резко замедляется, и естественный отбор становится лишь «стабилизирующим» [332]. В других условиях величины /j, шах будут иными — большими или меньшими, чем в данных. Соответственно иной будет и соответствующая диаграмма. Организмы, соответствующие разным эволюционным диаграммам (разным «экологическим нишам»), будут относиться к оазным таксономическим группам (рис. 7)
РИС. 8. Эволюционная диаграмма при нормировке приращений U по разным Kj

Каждый новый виток спирали осуществляется в результате ароморфоза — возникновения возможности нового возрастания Iij по уже пройденным ранее направлениям. 1( измеряется площадью диаграммы
т
в зависимости от ранга отличия Kj к разным видам, разным родам и т. д.
Анализ темпов эволюции соответственно по формуле (1) и диаграммам приводит, таким образом, к выводу о резко неравномерном темпе эволюционного совершенствования — быстром изменении площадей диаграмм до достижения предельных значений Ij замедлением скорости роста 1{ при переходе от Kj-го к Kj+i-му критерию отбора, и, наконец, неопределенно долгой задержке эволюционирующей системы в состоянии U, тах= 2/3-1тах (см. о неравномерности темпов эволюции работу Н. Н. Иорданского [122]).
Это свидетельствует в пользу реальности устойчивых дискретных таксономических единиц видов, родов [105, 189, 201, 202, 203].
Представленная выше кинетика эволюционного процесса основывалась на предположении, что lit ша* неизменны в данных условиях. Однако вполне возможно изменение /3-, тах и при постоянных внешних условиях в результате достижения некоторой степени совершенства по другим Kj+m-направлениям. С этого момента вновь становится возможным эволюционное совершенствование по ранее пройденным траекториям. Если эти изменения затрагивают все пройденные ранее направления, становится возможным «второй виток» эволюционной спирали»—начинает увеличиваться площадь диаграммы U в тех же, в принципе, внешних условиях (рис. 8). Такие изменения Ij по некоторым К, которые делают возможным возрастание /тах по другим /С,, можно назвать «ароморфозы» в соответствии со смыслом этого понятия, предложенного А. Н. Северцовым. [255, 256]. Таким
33
2 С. Э. Шнольобразом, ароморфоз делает возможным новый виток эволюционной спирали — заполнение (в том числе ранее заполненных) экологических ниш организмами с большей величиной 1тах.
ФИЗИКО-ХИМИЧЕСКИЕ И БИОЛОГИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ
Критерии, по которым осуществляется естественный отбор, т. е. различные Kj являются, в сущности, факторами эволюции (от facere — делать — лат.). Они определяют направления эволюции. Как мы видели, действие этих факторов происходит последовательно. Очередность действия этих факторов, как отмечено выше, может быть установлена на основе «принципа предельного совершенства» [339, 340].
Начальные этапы биологической эволюции определяются преимущественно физико-химическими факторами. Затем возникают, становятся критериями естественного отбора и собственно биологические факторы.
Биологическая эволюция начинается с началом естественного отбора матричных макромолекул, способных к конвариантному воспроизведению. Критериями отбора вариантов таких макромолекул являются их каталитическая активность, совершенство сопряженных процессов превращения энергии, скорость и экономичность превращения веществ, совершенство кодирования, записи и считывания наследственной информации, совершенство хранения (сохранения) наследственных текстов. Этот путь, по существу, чисто биохимической эволюции ограничен диффузионным барьером— медленностью диффузии метаболитов и ката- болитов. В связи с этим начинают доминировать факторы, направляющие эволюцию по пути ослабления и преодоления диффузионных ограничений. В результате возникают сложные морфологические структуры и аппараты активного перемещения в пространстве. Эволюция, направляемая отбором по критерию все более быстрого и совершенного перемещения в пространстве, приводит к возникновению мышц, нервов, рецепторов, центральной нервной системы. Несмотря на сложность результатов, например, возникновение нервной системы, критериями отбора, приведшими к ним, являются чисто физико-химические факторы эволюции.
Однако уже на этих этапах в действие вступают и собственно биологические критерии естественного отбора. Характерным свойством собственно биологических факторов биологической эволюции является то, что они сами возникают в результате биологической эволюции. К ним относятся прежде всего критерии, определяющие соответствие организмов в биоценозах. Действительно, предметом внимания естественного отбора оказывается и взаимное соответствие разных организмов в биоценозах. Общеизвестным примером эволюции, направляемой
собственно биологическим критерием отбора, является сопряженная эволюция цветков и опыляющих их насекомых, средств нападения хищников и защитных приспособлений жертв, симбиотических водорослей и грибов в лишайниках и т. п.
Мы видели, что скорость эволюционного совершенствования в данном направлении также находится под давлением естественного отбора. Если возможны несколько траекторий, ведущих к данной величине биологического совершенства, выбирается та, которая приводит к конечному этапу быстрее. Следовательно, наряду с факторами, обеспечивающими максимально высокую итоговую скорость конвариантной редубликации, ферментативного катализа, наибольшую эффективность превращения энергии, наибольшее совершенство морфогенеза, перемещения в пространстве, переработки информации — должны существовать •специфические факторы, определяющие (максимально возможную) скорость эволюционного совершенствования на каждом, этапе.
Факторы, определяющие скорость эволюции, могуть быть внешними и внутренними по отношению к эволюционирующей -системе. Внутренние — это те, которые сами возникают в результате эволюции. Это также собственно биологические факторы ■биологической эволюции. Скорость эволюционного совершенствования, как мы видели, зависит от узости интервала Д/С,- ч величины ДU. Биологические факторы в ходе естественного ■отбора по преимуществу влияют на величину Д/, т. е. на величину приращения U за некую единицу времени. А/ зависит от частоты мутаций, скорости размножения, скорости накопления полезных мутаций, эффективности действия естественного •отбора. Рассмотрим эти зависимости подробнее.
Частота мутаций определяется как внешними факторами, в частности, интенсивностью ультрафиолетовой и ионизирующей радиации, так и активностью биохимической репарационной •системы [61]. Ясно, что максимальная скорость эволюции достигается при поддержании частоты мутаций в некотором диапазоне значений, т. е. что для данных условий должна поддерживаться оптимальная частота мутаций.
Скорость увеличения биологического совершенства, т. е. скорость эволюции, возрастает при переходе от бесполого размножения к половому. Действительно, при бесполом размножении скорость возрастания биологического совершенства •определяется скоростью накопления в геноме полезных мутаций. Этот процесс зависит от вероятности мутирования и времени, необходимого естественному отбору для установления полезности или вредности данной мутации. Большинство «признаков», свойств живых организмов формируется в результате сочетания большого числа мутаций. Мутации происходят независимо друг •от друга. Полезной данная мутация может оказаться обычно лишь при условии предварительного осуществления и отбораряда других. Тем самым становится вероятной такая ситуация. В данном множестве родственных организмов имеется весь комплект случайно распределенных между особями потенциально полезных мутаций. Однако особи-носители большинства этих потенциально полезных мутаций гибнут в естественном отборе. Их полезность не проявляется в отсутствии полного комплекта всех предыдущих. Эволюция идет посредством отбора носителей первой полезной мутации и гибели всех прочих, ожидании появления второй полезной мутации у обладателей первой и т. д. На каждом этапе происходит «безрассудное уничтожение ценностей». Еще ярче это выявляется во вполне вероятной ситуации, когда никакая из мутаций в отдельности, без полного их комплекта, не дает преимуществ их обладателя в естественном отборе. Полный комплект данных мутаций очень полезен — сильно увеличивает кинетическое совершенство. В этом случае можно представить себе неопределенно долгую задержку эволюции на одном месте.
Положение резко изменяется при половой рекомбинации геномов. Теперь становится относительно быстро достижимым сочетание мутаций, резко увеличивающих биологическое совершенство обладателей всего комплекта. Правда, половая рекомбинация обеспечивает как сочетание генов разных особей, так и нарушение уже сложившихся полезных сочетаний. В этом случае для «дела эволюции» полезно чередование полового и бесполого размножения. При следующем за половой рекомбинацией бесполом размножении возникает множество копий, пусть еще незавершенного, по полезного набора генов. Испытание в ходе «борьбы за существование» особей, возникших при бесполом размножении, позволяет существенно увеличить концентрацию полезных сочетаний генов (мутаций). Так илн иначе, но и при половой рекомбинации — процессе перемешивания различных геномов—может потребоваться очень большое время, долгий ряд поколений. Все это время носители потенциально полезных мутаций должны сохраняться в системе, быть участниками общего «обменного фонда» генов. Целостность, сохранение этого обменного фонда на протяжении всего времени перебора сочетаний потенциально полезных мутаций—важное условие возможно быстрого возрастания биологического совершенства соответствующих организмов. Наибольшую опасность для сохранности, целостности данного обменного фонда генов представляет его преждевременное разбавление путем половой рекомбинации с представителями других обменных фондов. Поэтому условием ускорения эволюционного совершенствования оказываются изолирующие механизмы, все механизмы, препятствующие размыванию границ данного обменного фонда генов.
Итак, существование живых организмов в виде дискретных видов является условием ускорения эволюционного прогресса.
Пока мы пренебрегали колебаниями, случайными или периодическими долгопериодными изменениями условий существования. Я^сно, что может быть два способа эволюции в переменных условиях существования: 1-й способ — достижение такой скорости генетического приспособления, при которой эволюционные изменения успевают следовать за колебаниями условий существования; 2-й способ — демпфирование — нереагирование на относительно быстрые колебания условий существования, выработка приспособлений, позволяющих жить в соответственно широком диапазоне условий, т. е. такой темп генетических изменений эволюционного совершенствования, который обеспечивает увеличение биологического совершенства, учитывающее лишь постоянную составляющую изменений внешней среды.
му способу соответствует гаплоидный геном, когда каждая мутация сразу же проявляется в фенотипических свойствах ее носителя, сразу же становится объектом естественного отбора.
му способу соответствует полиплоидный (обычно диплоидный) геном, когда «на всякий случай» в каждом геноме имеются не проявляющиеся без должных сочетаний рецессивные гены.
2-й способ эволюции при наличии рецессивных и доминатив- ных алельных генов возможен, как ясно, лишь при половой рекомбинации, когда в гомозиготных организмах проверяется полезность данного гена, а в гетерозиготных — рецессивные гены «ждут своего часа» (см. [65]).
Можно представить себе, что в зависимости от условий существования наибольшая скорость эволюции может обеспечиваться иногда лишь весьма сложной стратегией: сменой гаплоидных и диплоидных поколений, чередованием полового размножения с неполовым, например, партеногенетическим.
Обсуждаемые в этой главе вопросы — сопоставление скорости эволюции при бесполом и половом размножении, образовании дискретных видов, гаплоидности и диплоидности, определенной очередности полового, бесполого, гаплоидного, диплоидного поколений — все это предмет для математического анализа на ЭВМ. Однако до последнего времени сделаны лишь первые шаги в этом направлении. При этом необходимо отметить, что еще[3] в 1930 и 1932 гг. Р. Фишер [387] и Меллер [432, 433] высказали ряд глубоких соображений о зависимости скорости эволюции от половой рекомбинации. Эти соображения близки приведенным нами выше. Однако строгий математический анализ с моделированием и исследованием посредством ЭВМ этой проблемы только начат. В работах Кроу и Кимуры [372, 373], Мейнарда Смита [424] и Фельзенстейна [386] исследуется вопрос о величине ускорения эволюции в результате половой рекомбинации при заведомо упрощенных предположениях, (см. также [450]). Даже при весьма сильных упрощениях Мейнард Смит [424] и Фельзенстейн [386] пришли к выводу о значительном ускорении эволюции при наличии половой рекомбинации. Мейнард Смит оценивал число поколений, необходимое для накопления примерно 10 полезных мутаций (независимых друг от друга) при бесполом и половом размножении. Он получил, что при размере популяции 109 пользе от каждой такой мутации в 0,01 (относительного ускорения размножения) и вероятности возникновения мутаций 10"9, эволюция ускоряется примерно в 6 раз. Сходные, в принципе, оценки получил Фель- зенстейн. Этот автор, кроме того, провел анализ на ЭВМ высказанного Меллером предположения о весьма быстром ускорении эволюции не столько от увеличения скорости накопления полезных мутаций, сколько от эффективного освобождения генома при половой рекомбинации от вредных мутаций. Проведенный анализ, имеющий явно предварительный характер, подтверждает предположение Меллера.
Можно надеяться на быстрый прогресс в этой области. Необходим математический анализ разных способов биологической эволюции, биологического смысла гаплоидности, диплоид- ности, дискретности видов, смены поколений.
Свидетельством реальности этих надежд можно считать работы В. В. Меншуткина и Б. М. Медникова по математическому, посредством ЭВМ, анализу эволюционных закономерностей и, в частности, процесса дивергентного видообразования [408—410].
Об исследованиях М. Эйгена и задачах книги. Во время работы над рукописью книги были опубликованы исследования М. Эйгена, посвященные проблемам самоорганизации материи и эволюции биологических макромолекул [350] *. Они производят большое впечатление ясностью и смелостью постановки и разработки принципиальных вопросов теоретической биологии. Мне представляется, кроме того, весьма интересным, что в ряде пунктов наши позиции оказываются вполне сходными. Такое сходство, по-видимому, имеет особый смысл: отталкиваясь от разных исходных положений, будучи специалистами в разных областях, основываясь на разных ассоциациях и научной предыстории, мы приходим к ряду аналогичных выводов. Поэтому ниже будут кратко рассмотрены некоторые основные аспекты работы Эйгена и отмечены сходство и различия наших взглядов.
Задача книги Эйгена формулируется им так: «Если мы хотим уничтожить разрыв между физикой и биологией, то необходимо разобраться в том, что такое «отбор» на языке точных молекулярных понятий, которые в конечном счете могут описываться квантово-механической теорией. Мы должны вывести дарвиновский принцип из тех свойств материи, которые нам известны» ([350, с. 21]). Рассмотрев необходимые предпосылки для осуществления отбора, Эйген пишет феноменологическое кинеги-
ческое уравнение для изменения численности каждого носителя информации (в нашей терминологии вида):
Xi = (Ft — Ri) Xt + Yi фахи i*i
где х{ — концентрация i-ro носителя информации, — определяет скорость «самоинструктированного» (в нашей терминологии матричного) его образования, R( определяет скорость уменьшения его концентрации (посредством распада, разбавления и т. д.). Последний член (сумма) уравнения характеризует все другие каналы изменения хи которое происходит в результате неточного копирования других матричных последовательностей, близких к i-той, фи определяет скорость образования таких копий.
Это уравнение описывает типичный автокатализ с увеличением численности л:4 при F^Rt. Величины Ft и Rt можно представить в виде:
Ft = kgAjQf, Ri = k0Di + Фиь
где — константы скорости копирования i-й матрицы, Qi—«фактор качества», показывающий, какая доля образовавшихся матриц представляет собой точные копии i-й, k„D — константы скорости уменьшения концентрации i-й матрицы, cpot имеет смысл, аналогичный ф«. С учетом этого основное уравнение Эйгена принимает вид:
Х{ =• kQ — Д] Xi + ^ ф цХ1 — ФоЛ.
Здесь весьма существенно замечание Эйгена, отмечающее, что это уравнение еще не описывает процесса отбора. Для отбора, для настоящей конкуренции за выживание необходимы внешние селекционные ограничения.
При ограничении суммарной численности всех N носителей информации (в нашей терминологии определенный ареал, ограниченные потоки веществ и энергии) получается следующее. Пусть —Д характеризует избыточную продуктивность;
тогда средняя продуктивность будет равна:
_ .V j N
Е = ^ Ekxk / ^ хк• k—i I k=i
Величину —Д Эйген называет селективной цен
ностью, заимствуя этот термин из работ Фишера, Райта, Холдена. После соответствующих преобразований Эйген получает основное уравнение, характеризующее динамику отбора:
Xi = k0 [Wi — Ё] Xi + >] фaxi.
ш
Теперь будет расти численность только тех носителей информации, _селективная ценность Wt которых оказывается выше порога Е. Здесь, вероятно, наиболее важно то обстоятельство, что средняя продуктивность Ё во всей системе непрерывно растет в ходе эволюции до тех нор, пока не будет достигнут ее оптимум, равный максимальной селективной ценности Wm, возможной для данной системы видов. Приближение Е к Wm совершенно аналогично рассматриваемому мною исчерпанию эволюционного потенциала в данном направлении эволюции.
Состояние с Wm на некотором этапе эволюции Эйген рассматривает как селективное равновесие и указывает, что оно имеет метастабильный характер. Это равновесие сохраняется до тех пор, пока не возникнет селективное преимущество с Wm+l>Wm. Как только появляется новая копия такого рода 4i+1, прежнее равновесие разрушается, и система идет к новому (метастабильному) состоянию равновесия.... ([350, с. 52]). Здесь мы видим явную аналогию с появлением очередного ароморфоза.
Далее Эйген анализирует стохастический подход к отбору, подчеркивая ограничения детерминистической теории отбора. По его мнению, элементарный процесс, ведущий к возникновению какого-либо конкретного мутанта, существенно недетерми- нирован, и автокаталитическое усиление ведет к макроскопическому отображению случайных микроскопических событий. Кроме того, он отмечает, что процесс роста численности сам по себе подвержен статистическим флуктуациям и указывает, что определенные стационарные состояния в отличие от истинно равновесных метастабильны и не могут стабилизироваться, и поэтому для поддержания их в течение длительного времени необходима регуляция [350, с. 75]. С учетом отмеченных ограничений Эйген проводит стохастическое рассмотрение процессов отбора.
Эти вопросы и особенно возможную недетерминированность направлений эволюции в результате автокаталитического усиления случайных событий, я рассматриваю в гл. 13, где на основании данных по биологической конвергенции прихожу к другим выводам.
Затем Эйген дает очень интересный анализ возможности эволюционного совершенствования, основанного на комплементарном узнавании только нуклеиновых кислот, и повторяет его для чисто белковых каталитических систем. Он приходит к выводу, что ни нуклеиновые кислоты, ни белки-ферменты не обеспечивают осуществления естественного отбора. Эйген рассматривает также систему, образованную нуклеиновыми кислотами и белками, в которой сочетается комплементарное инструктирование (матричное конвариантное воспроизведение в нашей терминологии) с каталитической связью. Эта система — само- воспроизводящийся гиперцикл — может эволюционировать,т. е. совершенствоваться, путем использования селективных преимуществ. Замечательно, что для гиперциклов с тремя и более членами теоретический анализ предсказывает колебания [350, ,с. 151] что, как отмечено мною, является 'необходимым условием ускорения эволюции.
Далее весьма кратко Эйген отмечает необходимость «ком- партментализации», т. е. образования дискретных особей, в качестве неизбежного следствия эволюции гиперциклов. Более детальный физико-химический анализ этого вопроса изложен в гл. 6. В своей книге он, как и я, рассматривает опыты С. Спигелмана в качестве иллюстрации верности теоретических критериев естественного отбора.
Эйген следующим образом подводит итоги всему сказанному: «Эволюция представляется неизбежным событием, если задано присутствие определенного вещества с определенными авто- каталитическими свойствами и если поддерживается такая величина потока (свободной) энергии, которая необходима для компенсирования стационарного производства энтропии» [350, с. 185]. «Тот факт, что «отбор» и «эволюция»—по аналогии с «равновесием» в термодинамике — можно охарактеризовать экстремальными принципами, позволяет физически обосновать и количественно сформулировать дарвиновскую теорию. В такой форме теория Дарвина уже не просто описывает некий исторический путь, но представляет собой выводимый из физики закон, управляющий общими процессами самоорганизации материи» [350, с. 197]. «Процессы автокаталитического отбора отфильтровывают и усиливают мутантов, обладающих высокой селективной ценностью, снижая тем самым недетерминированность, однако недетерминированность все-таки сохраняется в отношении выбора индивидуальных копий и затем отображается на макроскопическом уровне. Вследствие этого невозможно с достаточной точностью ни проследить прошлую, ни предсказать будущую эволюцию за пределами известных временных границ...» [350, с. 199].
При сопоставлении наших взглядов для дальнейшего изложения, вероятно, наиболее существенно сравнение понятий селективного преимущества и кинетического (биологического) совершенства. Смысл этих понятий очень близок. И и /; служат критериями в процессах естественного отбора. Величина Wm, введенная Эйгеном, аналогична понятию эволюционный потенциал данного направления эволюции, введенному выше.
Есть, однако, и существенное различие. Величина кинетического совершенства U характеризует (при прочих равных условиях) скорость возрастания массы веществ данного видя, a W{— приращение численности (особей, молекул) данного вида. Это различие становится важным при сопоставлении биологического совершенства дальних родственников, например, мелкихс крупными. Wi является непосредственным критерием отбора, U — как непосредственным критерием отбора, так и абсолютной характеристикой биологического совершенства данного вида.
Эйген ограничивает свою задачу физико-химическим анализом лишь первых шагов биологической эволюции. Основной проводимой здесь мною мыслью является применимость одного и того же критерия естественного отбора — кинетического (биологического) совершенства на всем пути биологической эволюции от «самого начала» до человека.
Мне не кажется правильным писать в настоящее время уравнения динамики эволюционного процесса для поздних его этапов. В сущности вся сложность динамики этого процесса скрывается в параметре К. в уравнении типа:
ihi = Kimi.
Однако К зависит от многих переменных — совершенства матричного конвариантного воспроизведения, совершенства катализаторов, совершенства процессов использования энергии, совершенства аппаратов перемещения в пространстве и т. д. Все эти переменные и являются факторами биологической эволюции‘. По существу их можно охарактеризовать в физико-химических терминах. Поэтому в книге и рассматриваются физико-химические факторы биологической эволюции. Все эти факторы поддаются количественной оценке.
В самом деле, т,- в конце концов определяется скоростью синтеза полимерных молекул, скоростью притока и оттока метаболитов, эффективностью использования энергии, длительностью нативного существования матричных молекул и других компонентов организмов.
Прослеживание детальных путей возрастания т( в зависимости от того или иного физико-химического фактора нереально. Зато вполне реальна оценка предельного по данному фактору значения т(. Полагая время, отводимое на эволюционное совершенствование в данном направлении достаточно большим, можно оценить предельно возможную, ограничиваемую лишь физическими законами, скорость синтеза полимерных молекул, предельно возможную эффективность преобразования энергии, предельно возможную скорость притока и оттока метаболитов, предельно возможную скорость перемещения организмов в пространстве и т. д. Такой «принцип предельного совершенства»,
как метод дед^кхивного анализа траекторий и динамики эволюционного проц'есса> оказывается, по-видимому, единственно возможным применительно к столь сложному предмету, каким является биолс)ГИЧеСкий эволюционный процесс.
Используя этот метод для прослеживания процесса эволюции «с самого начала», можно убедиться в том, что физико-химические факторы биологической эволюции выступают на эволюционную арену последовательно. Сначала возрастание величины тп{ определяется, например, скоростью синтеза полимерных цепей из предсуществующих, возникших «абиотически», мономеров. Потом оказывается, что рост определяется уже ростом концентрации этих мономеров, зависящим от скорости их биосинтеза. Потом оказывг^тся, что при некоторых величинах скорости синтеза полимеров и скорости (био) синтеза мономеров дальнейшее увеличение определяет «транспорт»—скорость притока
«пищи» и оттока «катаболитов». Так, одно узкое место, ограничивающее рост rhi, сменяет другое, один физико-химический фактор биологической эволюции перестает быть критерием естественного отбора и сменяется в этом качестве другим, очередным фактором. В силу этого процесс эволюции идет ступенчато: периоды быстрого увеличения при движении к физическому пределу по данному фактору эволюции сменяются периодами поиска новых принципов увеличения ть по новому критерию естественного отбора, соответственно очередному фактору эволюции.
Дальнейшая задача этой книги и состоит в выявлении и характеристике роли различных факторов эволюции на разных этапах биологического эволюционного процесса.
Глава 3
ЕСТЕСТВЕННЫЙ ОТБОР МАТРИЧНЫХ МАКРОМОЛЕКУЛ, СПОСОБНЫХ К КОНВАРИАНТНОМУ ВОСПРОИЗВЕДЕНИЮ
Спонтанное возникновение в планетных условиях полипептидов и полинуклеотидов, вовлекаемых в естественный отбор.
Необходимость однозначного соответствия полипептидных и полинуклеотидных полимерных цепей. Проблемы генетического кеда, прямого структурного соответствия полинуклеотидов и полипептидов
( «узнавание»).
Белки и нуклеиновые кислоты в дальнейшей эволюции. Разделение функций.
Фенотип и генотип.
Из многих в принципе возможных систем матричного конвари- антного воспроизведения, обладающих сходными физико-химическими свойствами, рано или поздно остается («побеждает») лишь одна с наибольшим эволюционным потенциалом. Таким образом, существенным этапом эволюции оказывается отбор эволюционирующей системы. Рассмотрим критерии выбора такой системы.
Эволюционный потенциал, естественно, сильно зависит от физико-химической природы матричных молекул.
Из простых соображений ясно, что /,• будет тем больше, чем больше удельная поверхность матричного вещества. Предельно большой поверхностью обладает одномерный кристалл — нить. Эволюционный потенциал зависит от разнообразия элементов матричной нити. Необходим набор элементов, обеспечивающий все возможные каталитические и матричные свойства.
Таким образом, максимально возможным эволюционным потенциалом будет обладать одномерная последовательность различных элементов кристаллической решетки — «букв, образующих строки». Следуя принятому нами при описании эволюционного процесса принципу предельного совершенства, мы можем отнести все сказанное о естественном отборе в полиморфной кристаллической системе к одномерному апериодическому кристаллу [344], т. е. к гетерополимерной макромолекулярной нити, состоящей из набора разных мономеров, порядок чередования которых в полимерной цепи может быть разным.
Необходимо лишь, чтобы такая нить обладала матричными свойствами, чтобы любое чередование мономеров в цепи было в принципе термодинамически равновероятным, но отличалось другот друга по кинетическим свойствам *. Этим общим условиям соответствуют нуклеиновые кислоты.
Однако прежде чем перейти к анализу возможных этапов эволюции в конкретных биохимических системах, рассмотрим условия, необходимые для начала биологической эволюции.
Условия неизбежности начала биологической эволюции. Эволюция становится неизбежной при выполнении основных условий: существования механизмов матричного, конвариантного воспроизведения и различия в скоростях воспроизведения разных форм полиморфных матриц.
Таким условиям удовлетворяет одномерная полимерная нить, для которой скорость матричного размножения, скорость синтеза матричных копий зависит от природы и последовательности мономеров.
При выполнении этих двух условий в ограниченном пространстве—ареале (третье условие)—при неравновесности самой системы или при наличии внешнего источника свободной энергии в среде, содержащей все необходимые компоненты для спонтанного возникновения линейных гетерополимеров (четвертая группа условий) начнется эволюционный процесс, и будет осуществляться естественный отбор по критерию наибольшей скорости образования полимера данного вида. Слово «вид» употреблено здесь вполне сознательно, хотя это еще совсем не такой же вид, как «ящерица прыткая» или «овес посевной» (см. гл. 2, стр. 36).
Однако реализация эволюционного потенциала системы, удовлетворяющей только этим условиям, может оказаться очень затрудненной. Ареал может быть заполнен формой, полимеризу- ющейся не с наибольшей скоростью. Затравка этой формы возникает случайно, в результате тепловых флуктуаций, и вероятность того, что случайно возникшая затравка соответствует наиболее кинетически совершенной форме очень мала. Вместе с тем заполнившая ареал полимерная форма будет препятствовать образованию более совершенных форм — материала для их построения не останется, а тот, который уже использован для построения заполнивших ареал матриц, защищен кинетическими потенциальными барьерами от посягательств пусть и способных к более быстрому воспроизведению зародышей, но возникающих позже затравок новых матриц. Речь идет, следовательно, о том, что одна даже самая удачная мутантная форма способная обеспечить наибольшую скорость матричного воспроизведения «ничего не сможет сделать» с уже заполнившей ареал менее совершенной массой полимерных молекул.
Нужно «открыть путь» для реализации полезных мутаций, реализации эволюционного потенциала. Этого можно достичь
;I Для этого энергии связей между мономерами в полимерной цепи должны быть одинаковыми в разных последовательных сочетаниях мономеров.
лишь одним способом: время жизни каждой матричной полимерной молекулы должно быть ограниченным, необходима закономерно наступающая смерть, т. е. распад матричных молекул по истечении некоторого времени жизни. Иными словами, условием длительного осуществления эволюции оказывается смена процессов синтеза процессами распада.
Каков в общем виде способ реализации вновь возникших мутаций в условиях заполненности ареала ранее образовавшимися видами?
Наверное, таких способов, по крайней мере, два. Вновь возникшие виды должны быть в состоянии «поедать» виды, образовавшиеся ранее, и использовать их материал для построения своих матричных молекул, или ранее появившиеся виды должны сами распадаться по истечении некоторого времени жизни. Первый способ (столь популярный на поздних стадиях эволюции), по- видимому, нереален на ранних интересующих нас сейчас стадиях. Представим себе даже, что вновь возникшие затравки — матрицы — обладают каталитическими свойствами, позволяющими им ускорить распад образовавшихся ранее. И что же? Эти каталитические свойства приведут к разрушению как ранее возникших, так и вновь возникающих. Невероятно спонтанное в результате однократной мутации появление в полимерной цепи такой последовательности мономеров, которая катализировала бы распад всех остальных последовательностей, а свою бы не разрушала. Для избирательного поедания представителей лишь чужого вида необходим длительный процесс эволюционного совершенствования, выработки механизмов различения «свое — чужое».
Для обеспечения распада матричных молекул нет необходимости придумывать сложные регуляторные механизмы — типа бомбы замедленного действия, взрывающей матричную полимерную молекулу после некоторого времени ее существования, ее жизни. Эволюция осуществляется в открытой системе, пронизываемой потоками энергии разного типа, которые обеспечивают эволюцию энергетически и служат причиной мутаций. Таким образом, в результате мутаций возникают не только затравки новых видов. Тепловые флуктуации, ультрафиолетовое и более жесткое излучения повреждают уже заполимеризованные матричные молекулы. Накопление возникших повреждений и приводит через некоторое время (время жизни) к разрушению полимерных молекул.
Однако на самих примитивных стадиях эволюционного совершенствования может преобладать весьма простой механизм, ограничивающий время жизни уже образовавшихся матричных форм.
В нашей абстрактной модели с полиморфной кристаллизацией время жизни данной г'-й формы тем меньше, чем быстрее она образуется. Действительно, одинаковая термодинамическая стабильность всех форм означает равенство всех констант равновесия Кг для всех i реакций кристаллизации (полимеризации) из мономеров в полимер i-го вида. Так как:
Kt=ki'/ki",
где k{ и к/' — константы скорости соответственно процессов образования и растворения (плавления) кристаллов, отношения всех к/ и к" равны для всех i: чем быстрее образуется кристалл {чем больше k/), тем быстрее он растворяется (тем больше к/').
В этой системе сначала все пространство заполняется преимущественно видом кристаллов, образующимся быстрее всего, а затем появляются имедленно растущие кристаллы. Через неограниченно большое время весь объем заполнится всеми возможными формами в соответствующих долях.
Все приведенное выше рассуждение относилось к закрытой системе. При условии притока энергии картина может оказаться совершенно иной; возможны кинетические «приспособления» — «барьеры», замедляющие распад даже .быстрее всего возникающей формы.
В этом случае жизнь полимерных матричных молекул слагается из двух качественно различных стадий, или периодов, или фаз: периода роста — относительно быстрой (и, быть может, ускоряющейся) полимеризации по возникшей затравке, т. е. периода матричного размножения, и периода относительно медленного умирания, т. е. накопления разрушений и, по-видимому, быстрого распада.
Эволюция нуклеиновых кислот — первый этап биологической эволюции. Главный кризис — необходимость возникновения полипептидных катализаторов-ферментов. Возможно, что перечисленным выше необходимым условиям начала биологической эволюции отвечает большое число различных по химической природе веществ — гетерополимеров. Реальное осуществление эволюционного процесса, выбор той или иной системы матричного воспроизведения зависит прежде всего от возможности спонтанного, доэволюционного образования соответствующих этим условиям веществ. Иными словами, общее направление эволюционного процесса с самого начала полностью детерминировано химическими и физическими свойствами среды.
Как следует из множества данных последних лет, в планетных условиях, т. е. при температурах и давлениях, соответствующих существованию полимерных химических веществ, с наибольшей вероятностью под влиянием ультрафиолетового и другого излучения из воды, углекислоты, аммиака, цианидов образуются аминокислоты, нуклеиновые основания, а также углеводы. Кроме того, спонтанно образуются нуклеотиды, полинуклеотиды и полипептиды (см. [1, 16, 23, 24, 53, 126, 127, 128, 132, 207, 228, 240, 243, 250, 261, 303, 306, 309а, 385, 389, 390, 408., 447, 448]).
Итак, аминокислоты, нуклеотиды и их полимеры — весьма вероятный исходный материал для естественного отбора. Намследует попытаться решить, какие же системы — полипептиды или полинуклеотиды — больше соответствуют указанным выше условиям, с неизбежностью приводящим к эволюционному совершенствованию. Необходимо, чтобы полимер синтезировался матричным образом с конвариантным сохранением вариантов- мутаций, чтобы скорости этого синтеза были различными для разных последовательностей мономеров в полимерной цепи и чтобы время жизни уже синтезированных матричных копий было не очень большим.
Для выбора возможного исходного объекта эволюции необходимо тщательное сопоставление реальных свойств полинуклеотидов и полипептидов.
«Матричность» полинуклеотидов бесспорна. Вопрос «матрич- ности» полипептидов должен еще стать предметом непредвзятого исследования (см. [421]). Можно предполагать, что скорости спонтанного синтеза и матричной полимеризации для разных последовательностей мономеров неодинаковы и для полипептидов, и для полинуклеотидов. Однако и этот вопрос требует тщательного исследования (литературного, теоретического и экспериментального) .
Старение — ограниченность времени сохранения нативности— свойственна и белкам, и нуклеиновым кислотам. Длительность сохранения нативности зависит от реальных условий.
Мы еще вернемся к этим вопросам, в частности к возможной «матричности» полипептидов. Пока же поступим следующим образом. Примем сначала, что матрицами являются только полинуклеотиды, что только они соответствуют условиям неизбежного начала эволюционного процесса и посмотрим, как при таком допущении должен идти эволюционный процесс. Затем примем другую версию — предположим, что матрицами являются и полипептиды и попытаемся понять как в таком случае должен осуществляться эволюционный процесс. А потом сопоставим полученное с современными биохимическими данными.
Представим себе первичную полинуклеотидную эволюционную систему. В среде спонтанно, в результате флуктуаций, возникают разные последовательности нуклеотидов в небольших полимерных цепях — образуются низкомолекулярные нуклеиновые кислоты со случайной последовательностью мономеров. По этим возникающим в результате флуктуаций затравкам идет кристаллизация (матричное воспроизведение, размножение), на что расходуются мономеры из среды. В конкуренции за «пищу» —мономерный материал для полимеризации — происходит естественный отбор определенных последовательностей нуклеотидов по признаку наибольшей скорости матричного синтеза.
Полимерные молекулы стареют и разрушаются. С некоторого момента темп эволюции, темп естественного отбора будет определяться именно скоростью разрушения возникших ранее полимерных молекул. Этот момент наступит тогда, когда скорость.
интенсивность, матричных синтезов сравняется и превзойдет интенсивность спонтанного образования в среде новых мономерных молекул нуклеотидов.
Так мы подошли к рассмотрению первого кризиса в биологической эволюции — дальнейшее совершенствование, т. е. дальнейшее ускорение матричного воспроизведения посредством уже существующих механизмов оказывается невозможным. Здесь эволюционирующая система может задержаться неопределенно долго до тех пор, пока не возникнет принципиально новый механизм ускорения кинетического совершенствования объектов эволюции. Есть два неисключающих друг друга пути выхода из этого самого острого в биологической эволюции кризиса: первый — выработка механизмов ускорения синтезов мономеров и второй — выработка механизмов ускорения разрушения «старых» полинуклеотидов. Избирательное ускорение определенных химических реакций есть катализ. Следовательно, основное содержание второго этапа биологической эволюции — возникновение в ходе естественного отбора предельно совершенных катализаторов-ферментов. Самое естественное начальное предположение состоит в допущении каталитических свойств у самих матричных полимерных молекул. В одном отношении такое допущение тривиально — матрицы избирательно катализируют синтез своих копий ‘.
Однако рассматриваемый нами кризис обусловлен не недостаточной скоростью синтеза на матрице полимеров из мономеров, а недостаточно высокой концентрацией именно мономеров. Было бы невероятно счастливым совпадением наличие нужных нам каталитических функций у самих матричных молекул, их способность ускорять процессы синтеза мономеров или разрушения старых полимерных молекул.
Более того, молекулы, способные к конвариантиому матричному воспроизведению, должны обладать (для обеспечения совершенства в этой функции) возможно меньшей химической активностью, возможно меньше «ввязываться» в разные химические процессы; их назначение—сохранение уникальных возникших в результате естественного отбора последовательностей мономеров в полимерной цепи. Иными словами, они обеспечивают наследственность и являются основой изменчивости. Каталитические функции должны быть отделены от них.
Тем не менее возникновение таких катализаторов-ферментов в ходе естественного отбора возможно лишь при условии зависимости синтеза этих катализаторов от последовательности мономеров в полимерной полинуклеотидной цепи. Иными словами, данная последовательность нуклеотидов должна определять свойства катализатора, возникающего на полинуклеотидной матрице.
В силу основного механизма эволюционного совершенствования катализаторы-ферменты по своему происхождению должны представлять собой одномерные нитчатые полимеры. Иначе невозможен постепенный процесс совершенствования катализаторов в результате отбора полезных мутаций, сопряженный с совершенствованием полинуклеотидных матриц.
Химические реакции, избирательный катализ которых необходим для обеспечения должной скорости синтеза матричных копий данного вида, сложны и разнообразны. Соответственно сложными и разнообразными должны быть функциональные свойства ферментов. Требованиям относительно большой реакционной способности, возможности существования в виде полимерных нитчатых молекул отвечают аминокислоты и их полимеры — белки. К тому же, как мы видели, аминокислоты и полипептиды легко возникают спонтанно в планетных условиях. Удовлетворимся земным опытом и будем считать разнообразие из 20 аминокислот достаточным для обеспечения всех свойств ферментов. Главным для нас в данном контексте является вопрос о способе сопряжения синтезов двух полимерных систем — полинуклеотидной и полиаминокислотной (полипептидной). Необходимо, чтобы последовательность аминокислот в полипептидной цепи определялась последовательностью нуклеотидов в полинуклеотидной цепи. Если наличие образующего в результате такого соответствия фермента способствует большей скорости матричного воспроизведения нуклеотидной последовательности данного вида, то и количество этого фермента тоже увеличивается быстрее.
Следовательно, на начальных стадиях эволюции, когда еще не возникли сложные механизмы обеспечения опосредованного соответствия полинуклеотидного и полипептидных синтезов, должна существовать прямая связь между последовательностью нуклеотидов и синтезом полипептидных цепей. Отсюда следует, что полинуклеотидная цепь должна непосредственно влиять на последовательность аминокислот и скорость синтеза полипептидной цепи *.
Итак, исходя из общих эволюционных соображений, следует,, что такая связь — непосредственный, избирательный катализ образования полипептидной цепи на полинуклеотидной цепи — должна была иметь место. Иначе не может начаться эволюционное совершенствование белков, не способных к самостоятельному матричному воспроизведению и, значит, к самостоятельному участию в процессе эволюции.
Получается, следовательно, такая картина. Нуклеотидная цепь ускоряет синтез полипептидных цепей. Образуются полипептиды, в которых последовательность аминокислот соответствует последовательности нуклеотидов. Некоторые последовательности аминокислот в полипептидной цепи оказываются способными ускорять синтез мононуклеотидов или ускорять распад уже существующих полинуклеотидов, ускоряя тем самым эволюционный процесс. Острейший кризис начального этапа биологической эволюции преодолевается — возобновляется естественный отбор полинуклеотидных матриц, причем теперь уже по признаку их способности обеспечить синтез все более каталитически совершенных полипептидных цепей.
Как ясно из сказанного, условием преодоления этого кризиса, условием продолжения биологической эволюции должно быть прямое, контактное соответствие нуклеотидных матриц по- липептидным отпечаткам. Посмотрим, в какой мере последнее- требование реализуется в действительности.
При первоначальном знакомстве с современной биохимией кажется, что такого соответствия нет. Последовательность аминокислот в полипептидных цепях определяется не непосредственным контактом аминокислот и нуклеотидов матричной цепи нуклеиновых кислот, а посредством адаптора — тРНК[4]. Необходимость адаптора в свое время предсказал Крик на основании геометрических соображений. Каждая аминокислота кодируется- тремя нуклеотидами. Нуклеотидный триплет много больше аминокислотного радикала и строгое геометрическое соответствие- триплета аминокислоте невозможно. Все верно. Однако известны весьма важные биохимические процессы, в которых осуществляется непосредственное, строго специфическое взаимодействие аминокислот и нуклеотидов. Так, сам процесс соединения аминокислотного ацильного остатка с гРНК основан на таком непосредственном взаимодействии, обеспечиваемом строгим специфическим соответствием аминокислотной последовательности в полипептидной цепи и нуклеотидной последовательности в полинуклеотидной цепи. Речь идет о процессе узнавания специфической для данной аминокислоты тРНК специфической же молекулой фермента—аминоацил-тРНК-синтетазой. Напомним' здесь высказывания В. А. Энгельгардта: «Зашифровку амино
кислот, осуществляемую действием фермента аминоацил-тРНК- синтетазы, можно считать узловым пунктом всего механизма биосинтеза белков. Здесь впервые скрещиваются и неразрывно переплетается биохимические функции обеих главенствующих групп биологических полимеров — белков и нуклеиновых кислот» [133, 353]. Итак, это «узнавание» происходит при строго специфическом непосредственном взаимодействии аминокислотных и нуклеотидных радикалов — белка-фермента и нуклеиновой кислоты. Можно было бы привести и другие примеры специфического взаимодействия аминокислотных и нуклеотидных последовательностей.
Механизм узнавания интенсивно изучается последние годы (см. [453]).
Представляют интерес взгляды на этот вопрос В. И. Брускова, изложенные им на нашем семинаре еще осенью 1966 г. Брусков полагал, что строгое однозначное соответствие нуклеотидных триплетов одной из 20 аминокислот может быть достигнуто при наличии соответствующего комплементарного словаря из трипептидных слов, т. е. при однозначном соответствии трех аминокислот трем нуклеотидам. Брусков шел в своей гипотезе дальше: он предположил, что четырехбуквенному нуклеотидному словарю мог бы соответствовать четырехбуквенный аминокислотный словарь и что этими аминокислотами как раз и являются три ароматических аминокислоты и одна неароматическа-л, я именно, триптофан, тирозин, фенилаланин и производное имидазола гистидин,
В основе этой гипотезы лежало старое предположение о возможной специфичности заряд-флуктуациониых взаимодействий в биологических системах. Хотя исследования последних лет, действительно показали, что наиболее сильные взаимодействия в водных растворах наблюдаются между ароматическими аминокислотами и нуклеотидами, заметной специфичности при этом выявить не удалось (см., например, [32]). Поэтому в настоящее время специфичность узнавания на такой основе представляется весьма сомнительной.
В недавней работе Брусков [39] существенно изменил и развил представления о возможных механизмах специфического нуклеотид-аминокислотного узнавания. Анализ проблемы в предположении о существовании аминокислот-, но-нуклеотидного кода показал, что наиболее вероятными кандидатами на роль аминокислот, участвующих в узнавании нуклеотидов, являются аминокислотные радикалы, способные к одновременному образованию двух (или большего числа) водородных связей. С помощью молекулярных моделей Брусков исследовал и показал стереохимическую возможность существования комплементарных к азотистым основаниям аминокислот, ди- и трипептидон. При этом в коде узнавания однонитчатых нуклеиновых кислот достаточно участия не более семи аминокислот (аргинина, аспарагина, аспарагиновой кис« лоты, глутамина, глутаминовой кислоты, серина и треонина), соответствующих четырем типам различных химических групп, образующих водородные связи, а именно, гуанидиновой, амидной, карбоксильной и гидроксильной группам. Легко видеть, что минимальное необходимое число химических групп, участвующих в узнавании четырех нуклеотидов, равно двум. Из опытов с моделями ^сделан также вывод о вырожденности и конформационном характере кодаузнавания однонитчатых полинуклеотидов. Именно для специфичности оказы- ;ается необходимой строго определенная стабилизация в пространстве боко- ых радикалов аминокислот и пептидных групп, что, возможно, осуществляет- я в белках за счет стерических ограничений или дополнительных связей.
В работе [38] рассмотрены возможные физико-химические механизмы, при омощи которых матричные ферменты — полимеразы могут влиять на степепь эчности матричных синтезов и, следовательно, регулировать ее. В частности, редполагается, что ферменты синтеза и репарации нуклеиновых кислот содер! •;ат «узнающий участок», определяющий правильность комплементного спаря- ания (в результате образования дополнительных водородных связей) по инвариантным для комплементарных пар атомам N = 3 пуринов и 0 = 2 пирими- динов. Такой узнающий участок (центр) фермента играет роль дополнительной белково-нуклеиновой матрицы, способной «усиливать» отбор правильных и выбраковывать неправильные пары.
Вернемся, однако, к начальным стадиям эволюции [388]. На начальных этапах эволюции вовсе не требуется строгого однозначного соответствия нуклеотидов и аминокислот. Конфигурации белковых макромолекул грубо определяются не строго однозначной аминокислотной последовательностью, а лишь порядком чередования в полипептидной цепи полярных и неполярных аминокислотных радикалов. Исходя из этого, все аминокислоты можно разделить на два класса — полярные и неполярные. Может быть, полярные аминокислоты следует в свою очередь разделить на отрицательно и положительно заряженные в водных растворах — тогда будет три класса. Таким образом, в начале эволюции было бы достаточно, чтобы одни нуклеотидные радикалы в полинуклеотидной цепи непосредственно кодировали связывание полярных аминокислот, а другие — неполярных. Здесь следует отметить работу М. В. Волькенштейна [55], обнаружившего корреляцию между нуклеотидным составом кодирующих триплетов (кодонов) и полярностью кодируемых ими аминокислот. Волькенштейн обратил внимание на то, что во всех случаях, когда второй нуклеотид в кодоне — аденин, кодируемый .аминокислотный остаток полярен, во всех случаях, когда второй нуклеотид — уридин, аминокислотный остаток неполярен. Я думаю, что мы имеем здесь дело с корреляцией, обусловленной физико-химическими особенностями непосредственного взаимодействия аминокислот и нуклеотидов, действовавшей в древнейшие времена, когда современный перевод нуклеотидного языка в аминокислотный еще не сформировался. Сам Волькенштейн рассматривает эту корреляцию как приспособление, повышающее «помехоустойчивость» кода: если в результате мутации изменится кодон, то велика вероятность того, что вместо одной, например, неполярной аминокислоты в кодируемом белке появится другая, но также неполярная. Конфигурация макромолекулы от этого изменится не очень сильно, и мутант не погибнет. Мне же кажется, что в ходе эволюции такая корреляция могла возник-
нуть лишь в том случае, если по своим физическим свойствам аденин лучше «приспособлен» к взаимодействию с полярными аминокислотами, а уридин —• с неполярными.
Следует сказать, что и Волькенштейн допускает мысль о непосредственном взаимодействии аминокислот с полинуклеотидной цепью на ранних стадиях эволюции. «Возможно, матричный синтез белка на заре жизни осуществлялся иначе, скажем, без участия тРНК, а посредством прямой сборки белковой цепи на полинуклеотиде. Вероятность такой сборки, однако, невелика.. Скорее следует думать, что современный код является результатом селективного давления естественного отбора, минимизирующего эффект мутаций» [55].
Анализируя проблему возникновения жизни, Д. С. и Н. М. Чер- навские [322] выдвинули гипотезу о возможном механизме установления соответствия между последовательностями нуклеотидов и аминокислот. Они предположили, что двойная полину- клеотидная спираль служит гетерогенным катализатором, ускоряющим синтез пептидных связей между аминокислотами, адсорбированными на полинуклеотиде. Так образуется белковый чехол, предохраняющий полинуклеотидную двойную спираль от разрушений, который сам может обладать каталитическими свойствами, способствующими тем или иным путем образованию' полинуклеотидных цепей. Для того, чтобы такой механизм играл биологическую, т. е. эволюционную, роль, последовательность, аминокислот, образующих белковый чехол, должна зависеть от последовательности нуклеотидов. Гипотеза ценна тем, что ее можно проверить экспериментально (см. также [371]).
Наличие даже слабой корреляции между последовательностями нуклеотидов и аминокислот в синтезируемой на полинуклеотидной матрице полипептидной цепи позволяет продолжиться естественному отбору. В конкуренции за «пищу и пространство» будут побеждать такие полинуклеотидные последовательности, на которых синтезируются полипептиды, способствующие более быстрому размножению матриц своего вида. Критерием отбора на этом этапе служит каталитическое совершенство образующихся белков-ферментов, которое в свою очередь зависит от степени совершенства перевода последовательности нуклеотидов в последовательность аминокислот в полипептидной цепи. Таким образом, необходимость совершенствования каталитических свойств белков-ферментов, вызванная давлением естественного отбора, обусловливает направление эволюции в сторону совершенствования механизма перевода нуклеотидного языка на аминокислотный. Предел совершенства этого механизма — полная детерминированность последовательности всех аминокислот, необходимых для полноценного функционирования белков, последовательностью нуклеотидов в матрице — нуклеиновой кислоте. Мы приходим, следовательно, к необходимости кодирования примерно 20 аминокислот четырьмя нуклеотидами, т. е. к механизму,.
в котором каждая аминокислота должна соответствовать определенному сочетанию по крайней мере трех нуклеотидов (кодону). Недостаточность простого соответствия аминокислота — нуклеотид, необходимость трех нуклеотидов для кодирования каждой аминокислоты обусловливают обязательное существование адаптора при синтезе белка на нуклеиновой матрице. Однако преодоление этого очередного кризиса представляется мне более легким, чем предыдущего — хоть какие-то уже ферменты есть. Ключевым пунктом при этом оказывается аминоацил-тРНК-син- тетаза (ее древние предшественники), т. е. фермент, катализирующий соединение аминокислоты с адаптером — полинуклеотидом, содержащим антикодон, комплементарный специфическому для данной аминокислоты кодону. Аминокислоты, «подвешенные» посредством эфирной связи к кислороду гидроксильной группы рибозного кольца молекулы адаптора, в должной последовательности соединяются в полипептидную цепь. Прямо же код, основанный на непосредственном физико-химическом соответствии аминокислот и нуклеотидов (см. выше), продолжает использоваться в механизме узнавания при функционировании аминоцил- тРНК-синтетазы, а также в процессах регулирования считывания генетической информации.
Мне хочется еще раз подчеркнуть «автоматический», «циклический», характер эволюционного совершенствования после преодоления первого кризиса: ферменты, образующиеся в соответствии с нуклеотидным текстом, возникающим в результате отбора мутаций, будут тем совершеннее, чем «совершеннее сам текст». В понятие совершенства текста входит качество инструкций о механизме его перевода на аминокислотный язык, и, следовательно, о свойствах синтезируемых ферментов. Получается эффективная система (цикл) положительных обратных связей, приводящая ко все более эффективной, все более быстрой эволюции в направлении все большего кинетического совершенства конвариантно размножающихся матричных полимерных систем *.
В мою задачу не входит детальное описание биохимических механизмов синтеза белка в ныне живущих организмах. Это прекрасно сделано в ряде оригинальных книг. Мне важно подчеркнуть физико-химическую детерминированность эволюционного возникновения связи, корреляции синтезов полинуклеотидов и полипептидов, а также основных механизмов этих синтезов. Естественный отбор в планетных условиях рано или поздно с неизбежностью должен привести к формированию предельно совершенного механизма синтеза белка и нуклеиновых кислот, аналогичного механизму, работающему в системе рибосома — ферменты.
Возможность начала эволюции с полипептидов. Теперь посмотрим, каким должен быть процесс эволюции, начинающийся
1 М. Эйген называет эту систему «гиперциклом» [350]; см. гл. 2, с. 40.
с отбора все более кинетически совершенных полипептидов. Условия неизбежного начала эволюции остаются те же — матричное конвариантное воспроизведение, различия в скоростях воспроизведения (синтезов) разных форм матриц, конечное время жизни каждой (и, разумеется, ограниченность ареала). Наиболее важен здесь ответ на первый вопрос — способны ли полипептиды к матричному конвариантному воспроизведению?
Тут уместно отметить, что для полинуклеотидов мы полагали положительный ответ очевидным. Однако, как это ни парадоксально, взаимно комплементарные матричные свойства нуклеиновых оснований в водных растворах не проявляются. Их удается выявить лишь в «вакууме»: вода, конкурируя за водородные связи, препятствует спариванию комплементарных нуклеиновых оснований. Подобные «вакуумные условия» возникают в поли- нуклеотпдной цепи при ее спирализации: внутрь спирали вода не проникает, и поэтому ничто не мешает комплементарному спариванию. Таким образом, эволюционный процесс мог бы начаться лишь после спонтанного возникновения биспиральных поли- нуклеотидных молекул со случайной последовательностью нуклеотидов (при условии разных кинетических свойств различных по- линуклеотидных вариантов). Как мы видели, однажды начавшийся процесс естественного отбора «чистых» полинуклеотидных матриц очень быстро (т. е. через небольшое число актов эволюционного совершенствования) прекращается — эволюционный потенциал исчерпывается. Для выхода из этого кризиса необходимо осуществление сопряженного синтеза катализаторов с достаточно разнообразными кинетическими свойствами, т. е. белков.
Тем более важно выяснить, не могут ли полипептиды вступать на путь естественного отбора независимо от нуклеиновых кислот.
Да, могут, если они способны к конвариантному матричному воспроизведению. Н. К- Кольцов предполагал, что полипептид- ные цепи обладают этой способностью и, быть может, такая его позиция кажущаяся неверной с точки зрения современной нам биохимии, не вовсе ошибочна.
Способность белков, полипептидных цепей к точнейшему, строго специфичному различению молекул (что служит необходимым условием матричной полимеризации), общеизвестна. Прославленная специфичность ферментов — один из примеров этой способности.
Известно много разных ферментов, строго специфичных к той или иной аминокислоте. Можно назвать все те же амино- ацил-тРНК-синтетазы, каждая из которых специфична для одной данной аминокислоты, протеолитические ферменты, разрывающие быстрее всего пептидную связь около определенной, специфичной для данного фермента аминокислоты (ее радикала) и т. д. Однако кажется вероятным, что во всех перечисленных выше случаях специфичность по отношению к данной аминокислоте обусловлена специфической укладкой полипептидной цепи фермента, в результате чего в активном центре (точнее, в центре связывания аминокислоты на ферменте) достигается необходимое пространственное взаиморасположение нескЬ^ьких (двух, трех) аминокислотных радикалов. Впрочем, точно это еще неизвестно. Быть может, сама специфичность связывания ферментом данной аминокислоты обусловлена наличием лишь одного такого же или комплементарного ему аминокислотного радикала полипептидной цепи фермента. Остальные аминокислотные радикалы в центре связывания выполняют другие функции — собственно каталитические, а именно обеспечивают поляризацию связанной молекулы, ее деформацию и т. п.
Может показаться, что я ломлюсь в открытую дверь — способность аминокислот к кристаллизации — чего еще желать для иллюстрации матричных свойств аминокислот? Однако возможность кристаллизации аминокислот в значительной мере определяется геометрией их заряженных групп (отрицательного карбоксила и положительно заряженной аминогруппы). Геометрия этих ионизированных групп у всех аминокислот практически одинакова. Поэтому вряд ли можно ожидать избирательной кристаллизации аминокислот из их смеси. Впрочем, возможна ли такая избирательная кристаллизация, я не знаю (но очень хотел ■был бы узнать). Вместе с тем общеизвестна способность белков, полипептидов образовывать кристаллы. К сожалению, это их свойство не очень проясняет картину. Во-первых, не известно, возможна ли избирательная кристаллизация белков, во-вторых, она определяется не столько детальным соответствием полипептидных цепей друг другу, сколько сходством геометрии целых белковых глобул. Кроме того, нас интересует способность полипептидных цепей служить матрицами при синтезе новых полипептидных цепей из аминокислот. Это значит, что нас интересует специфическое связывание аминокислот на полипептидной цепи.
Насколько мне известно, возможность такого специфического связывания не исследована. Много лет назад я занимался изучением связывания меченных (метками служили радиоактивные изотопы углерода или серы) аминокислот белками [334]. Белки действительно связывают аминокислоты, причем для разных аминокислот это происходит неодинаковым образом. Однако специфичность такого связывания, его обусловленность аминокислотным составом белка должным образом не изучены. Всего- то речь идет об опыте, в котором... нужно измерить количество разных аминокислот, неотмываемых от полипептидов разного ■состава.
Особый интерес в связи с обсуждаемыми вопросами представляют взгляды Н. Г. Есиповой, изложенные ею по моей просьбе ниже. «Еще раз напомним, что Крик выдвинул адаптерную гипотезу, в которой соответствие между аминокислотой и трину-клеотидом устанавливается специальной молекулой-адаптером (транспортной РНК), только потому, что он не усмотрел кон- формационного соответствия между тринуклеотидом в развернутой конформации и аминокислотой. Важная черта адаптер- ной гипотезы состоит также в том, что длина кодона становится унифицированной, не зависящей от размера аминокислоты. Адаптер является по существу элементом логической машины и с его помощью могут исправляться ошибки, которые были бы фатальны при прямом конформационном соответствии аминокислот и нуклеотидов. Весьма привлекательна, однако, мысль, что на ранних этапах эволюции между полинуклеотидами и аминокислотами существовало такое непосредственное соответствие. Подобный механизм обладает очевидным преимуществом и очень прост, кроме того он обеспечивает обратное влияние аминокислотной последовательности на полинуклеотидную, а подобного рода обратная связь, быть может, и создает то недостающее ускорение эволюции, которое позволило начальным ее этапам уложиться в известные нам сроки.
Вначале рассмотрим возможное непосредственное соответствие аминокислоты и кодирующего ее триплета. Анализ стереохимии аминокислот и триплетов, входящих в состав ДНК, проведенный в двух лабораториях почти аналогичными методами показывает, что всегда можно заметить специфическое соответствие боковых радикалов аминокислоты и триплета из нуклеотидов, если аминокислота расположена в малой бороздке двойной спирали ДНК. Структурное подобие описанного типа наглядно определяет степень вырожденности кода — короткие аминокислоты (гли, ала, сер) кодируются фактически одним нуклеотидом, средние — двумя и, наконец, наиболее длинные — арг, лиз, три— тремя нуклеотидами. Наличие такого соответствия можно считать одним из факторов эволюции сопряженного синтеза нуклеиновых кислот и белков.
Одно из проявлений соответствия между нуклеотидами и белковыми (аминокислотными) структурами следует усматривать в супрессии бессмысленных мутаций у бактерий. Механизм супрессии состоит в нивелировке мутаций, приводящих к вклиниванию бессмысленного кодона в структурный ген. В результате мутаций тРНК-адаптор, содержащий антикодон с бессмысленным триплетом, начинает транспортировать нужную аминокислоту в нужное место при синтезе белка. Казалось бы ошибка в: адапторе исправляется на уровне адаптора же. Но на пути трансляции аминокислоты имеется специфический фермент тРНК-синтетаза—именно в этом месте, видимо, работает механизм непосредственной связи между нуклеотидной и аминокислотной последовательностью».
Однозначное соответствие последовательности нуклеотидов- в синтезируемом полинуклеотиде последовательности аминокислот в уже существующем полипептиде, т. е. «обратный переносинформации» кажется в настоящее время большинству ученых чистой крамолой. Установление такого соответствия\означало бы, что «центральная догма» молекулярной биологии — не догма, что знаменитая схема ДНК.-*-РНК->белок, уже нарушенная открытием обратной транскриптазы (в присутствии которой оказалось возможным направление переноса информации РНК->- —>ДНК), нарушается и в следующем звене.
Как ясно из сказанного, об узнавании белком определенной последовательности нуклеотидов и из исследований Н. Г. Есиповой и В. Г. Туманяна, а также Томас [466], физико-химическое соответствие полинуклеотндных и полиаминокислотных последовательностей вполне реально. Очевидно, однако, что глобулярные белки уступают по своим матричным свойствам нуклеиновым кислотам вследствие своей упаковки (в общем случае вследствие своей сложной третичной структуры). Естественна мысль, что фибриллярные белки, не свернутые в глобулу полипептидные цепи, могут в принципе служить вполне «хорошими» матрицами (так думал Н. К. Кольцов!) как для саморепликации, так и для снятия с них полинуклеотндных комплементарных копий. Эта мысль развита в очень интересной статье Картера и Краута [371] (о возможности матричного воспроизведения на белках см. также [306]).
Картер и Краут показали, что весьма распространенная вытянутая конформация полипептидных цепей (р-складчатая структура по Полингу и Кори) соответствует стабильной правозакру- ■ченной двойной спирали, т. е. существует двойная полипептидная спираль. Ее шаг и радиус примерно такие же, как и у двойной спирали нуклеиновых кислот. Авторы весьма тщательно построили пространственные молекулярные модели и увидели, что полипептидная двойная спираль в точности комплементарна двойной спирали РНК, причем полипептидная двойная спираль точно входит в малую бороздку двойной спирали РНК. В результате ■образуются водородные связи между 2'-ОН-группами рибозы (что, по-видимому, и объясняет особые свойства РНК, у ДНК нет этого гидроксила) и кислородом карбонила пептидной связи. Авторы отмечают, что такую точную комплементарность стабильных конформаций РНК и полипептидов вряд ли можно считать простым совпадением. Они предположили, что обе эти двойные спирали взаимно катализировали сборку друг друга из активированных предшественников на ранних стадиях эволюции. В самом деле, комплементарное соответствие двух двойных спиралей представляется весьма важным, но оно обеспечивает ускорение синтеза лишь основных каркасов — полипептидного и полинуклеотидного, тогда как для преодоления обсуждаемого нами кризиса на начальной стадии биологической эволюции необходимо установление полного однозначного соответствия определенных аминокислотных и нуклеотидных радикалов. Для этого нужно, чтобы «взаимная» полимеразная активность поли-
нуклеотидной и полипептидной двойных спиралей зависела от последовательностей нуклеотидов и аминокислот соответственно. Существование такой зависимости может показать лишь эксперимент.
В октябре 1975 г. опубликована статья Г. В. Гурского,,
В. Г. Туманяна, А. С. Заседателева, А. Л. Жузе, С. Л. Гроховского и Б. П. Готтиха. «Код, управляющий специфическим связыванием регуляторных белков с ДНК, и структура стереоспецифиче- ских участков регуляторных белков» [79]. Как ясно из заглавия статьи, в .ней расшифрован второй биологический код, выяснен механизм однозначного соответствия полинуклеотидных и полипептидных цепей в процессах узнавания. Механизм узнавания основан на специфическом взаимодействии двух двойных спиралей— нуклеотидной и полипептидной [371]. Регуляторные белки узнают последовательность оснований в двойной спирали ДНК; не расплетая ее. Узнавание основано на пространственном соответствии контактных, связывающих друг с другом группировок в полипептидных и полинуклеотидных спиралях. Оно аналогично специфическому совпадению отверстий в двух налагаемых друг на друга перфокартах. Такой способ установления однозначного соответствия авторы называют решеточным принципом узнавания. В качестве контактных групп в нуклеотидных цепях функционируют или гуанин, или цитозин, тимин и аденин, а в полипептидных цепях — только атомы азота амидных групп полипептидного остова. Амидные азоты связываются посредством водородных связей с контактными группами полинуклеотидных цепей. С гуанином способны образовывать водородные связи атомы амидного азота полипептидной цепи; их конформация рассчитана авторами и она оказалась отличной от конформации полипептидной цепи, способной взаимодействовать с тимином. Такие конформационно различные полипептидные цепи называются соответственно g- и /-цепями. Оказалось, что эти две анти- параллельные полипептидные цепи, находящиеся в g- и /-конформациях, могут соединяться в полипептидную двойную спираль водородными связями, образующими между амидными группами двух цепей, не взаимодействующими с основаниями ДНК.
Специфичность связывания белков и нуклеиновых кислот объясняется тем, что боковые радикалы некоторых аминокислот в /-цепи (названные авторами фундаментальными аминокислотами) могут образовывать водородные связи с карбонильными группами g-цепи, разрывая или резко ослабляя водородные связи между g'-цепью и гуаниновыми основаниями ДНК Таких фундаментальных аминокислот шесть: серии, треонин, аспарагин, ци- стеин, гистидин и глутамин. Таким образом, код вырожден в отношении аминокислотных последовательностей в стереоспецифи- ческом участке белка. Он требуетприсутствия аминокислот — «выключателей» в определенных местах последовательности. Авторы отмечают, что высокая степень вырожденности кода по-
зволяет совершенствовать трехмерную структуру и каталитические центры регуляторных белков в ходе эволюции, оставляя неизменной их способность находить правильные места"связывания на ДНК.
Предложенный авторами код узнавания проверен ими путем исследования соответствия последовательности нуклеотидов в. 1ас-операторе и последовательности аминокислот в 1ас-репрессо- ре Е. coli. Они показали наличие единственного участка полипептидной цепи (от треонина 19 до валина 30), находящегося согласно принятому механизму узнавания в полном соответствии с последовательностью нуклеотидов в ДНК.
Если бы даже эволюция началась с возникновения полипептидных матриц, все равно произошел бы кризис. Сохранение- матриц трудно совместимо с относительно высокой химической активностью полипептидов. Две противоположные тенденции—«консерватизм» и высокую каталитическую активность, нельзя с должным совершенством объединить в одном веществе. Поэтому, начнись дело с белков, все равно наступил бы кризис— побеждали бы матричные полипептиды, катализирующие синтез химически малоактивных «информационных» полимерных молекул (т. е. тех же нуклеиновых кислот) и в данном случае кризис удалось бе преодолеть лишь посредством выработки однозначного соответствия последовательности аминокислот в полипептиде последовательности нуклеиновых оснований в полинуклеотиде.
Это чисто диалектическое противоречие. С одной стороны, необходимость возможно более совершенного сохранения накопленного в поколениях запаса полезных мутаций, сохранение наследственных текстов, и с другой — обеспечение возможно более интенсивной жизнедеятельности и достижение темпа мутаций, достаточного для обеспечения возможно большей скорости эволюционного совершенствования.
Первая консервативная задача решается переходом функции сохранения наследственных текстов к специализированным молекулам ДНК- Почему именно ДНК? Вероятно из-за отсутствия гидроксила в положении 2 углеводного кольца дезоксирибозы, что делает ДНК химически более инертной, чем РНК.
Наличие двойной спирали ДНК существенно повышает сохранность наследственных текстов — повреждение одной из двух нитей в двойной спирали легко восстанавливается: уцелевшая нить служит матрицей для восстановления правильной последовательности мономеров в поврежденной.
Н. Г. Есипова и В. Г. Туманян следующим образом объясняют физико-химические причины функциональных различий ДНК и РНК:
«...Химические различия РНК и ДНК невелики: ОН группа во 2-м положении рибозы и урацил вместо тимима. Однако эти небольшие химические различия обусловливают существенные-различия их конформаций. Конформация двуспиральной РНК обычно A-форма, конформация ДНК — 5-форма. 5-форма характеризуется увеличенной по сравнению с А формой проекцией нуклеотидов на ось опирали: 13,4 А вместо 2,8 А. Однако главное стереохимическое отличие 5-формы от A-формы заключается в том, что в первой расстояния между любой соседней парой фосфатов отличаются друг от друга меньше, чем в A-форме. Расчет потенциальной энергии различных конформаций ДНК, проведенный методом атом-атомных потенциалов показал [120], что единственным устойчивым с точки зрения возможности достижения минимума потенциальной энергии типом структуры ДНК является 5-форма. A-форма сама по себе нестабильна: она дестабилизируется электростатическими взаимодействиями. A-форма существует лишь при условии компенсации роста свободной энергии электростатических взаимодействий возрастанием энтропии в системе полинуклеотид — вода.
*~^СИСТ ’ *5ДНК “Ь 5раств.
Увеличение 5СИСТ возможно либо за счет 5ДНК — например, если линейная молекула ДНК свертывается в макроструктуру, либо за счет SpaCTB — например при разупорядочении воды. Последнее возможно при добавлении в раствор сильного и структурно подвижного дегидратанта. Наличие полимерного дегидра- танта увеличивает вероятность перехода молекулы нуклеиновой кислоты в квазиглобулярное состояние, но не в А-форму.
Таким образом конформация A-типа возникает как частный случай конформационного перехода с увеличением энтропии системы ДНК-растворитель. Энтропийная стабилизация А-конфор- мации делает ее неустойчивой по отношению к внешней полимерной среде. Напротив, 5-форма устойчива как в такой среде, так и при комплексировании с полимерами. Это определяет основной тип взаимодействия ДНК с белкам,и и лолимерными лигандами — взаимодействие типа решетка — решетка.
Из рассмотрения физико-химических особенностей ДНК и РНК следует возможная причина уникальной роли ДНК как хранителя информации в сложных биологических системах. РНК как структура неравновесная, не может быть основой для иерархического упорядочения при наличии в результирующей системе нескольких уровней иерархии, так как организация следующего уровня сложности в системе требует устойчивости предыдущего уровня, являющегося «строительным кирпичом» для следующего. В то же /время в процессах считывания информации важно, чтобы полимер с одной стороны удовлетворял условиям комплементарное™, а с другой был бы легко разъединяем с матрицей — носителем информации. Этим требованиям удовлетворяет РНК, образующая с ДНК энтропийно или кинетически стабильную А-форму.
Таким образом, данные о потенциальной энергии для различных структур ДНК и РНК могут служить объяснением, почему два типа нуклеиновых кислот, идентичных по многим физикохимическим свойствам, различны по своим функциям в процессах жизнедеятельности».
Вторая задача, в частности достижение темпа мутаций, достаточного для возможно большей скорости эволюции, решается не так просто. Темп мутаций должен быть «соразмерным» — не слишком быстрым и не слишком медленным. Таким образом, вторая задача решается выработкой механизмов регулирования: частоты мутаций. В последние годы была обнаружена неожиданно сложная и совершенная биохимическая система регулирования’ темпа мутаций. Один из существенных ее механизмов — процесс ферментативной репарации поврежденных молекул ДНК, катализируемый так называемыми репаразами (см. [61]).
Обеспечение возможно более интенсивной жизнедеятельности решается передачей соответствующих функций белкам. Однаю> интенсивная работа «дорого стоит» макромолекулам белков. Эти молекулы все время повреждаются и ломаются. Взамен синтезируются новые. Таким образом, чем интенсивнее функция белков, тем интенсивнее они обновляются. Это постоянное их обновление стало очевидным после первых же опытов Р. Шонхей- мера с изотопной меткой (см. [244]). По существу, после возникновения механизма синтеза белка на полинуклеотидных матрицах, дальнейшая эволюция состоит в совершенствовании белков, полипептидных цепей. Кинетические свойства самих матричных полинуклеотидных молекул перестают быть факторами эволюции. Фенотип, т. е. результат реализации наследственных свойств в жизнедеятельности (кинетические свойства) отделяется от генотипа, т. е. совокупности наследственных текстов (информационно-термодинамические свойства), т. е. белковый фенотип отделяется от нуклеотидного генотипа. Такое разделение кинетики и термодинамики, фенотипа и генотипа, не нужно понимать слишком буквально. Речь идет лишь о ведущих критериях естественного отбора, физико-химических факторах эволюции.
Дарвиновские эксперименты Спигелмана. Естественный отбор в растворе макромолекул. Справедливость представлений о приложимости дарвинизма к процессам на молекулярном уровне, возможность естественного отбора в растворе макромолекул, доказана Спигелманом [460]. Более того, из его опытов следует и правильность вывода о кинетическом совершенстве как критерии естественного отбора.
Спигелман исследовал репликацию молекул РНК, катализируемую выделенным из Qd-фага очищенным ферментом, Qp-pe- пликазой. Этот фермент использует в качестве матриц молекулы фаговой РНК и катализирует образование на матрице новых молекул РНК из нуклеотидов. При проведении синтеза рибонуклеиновой кислоты в эксперименте было обнаружено возник
новение «мутантов» — молекул РНК, отличающихся от исходных матричных молекул большей скоростью репликации. Такие мутанты размножались при посредстве того же фермента Qp- репликазы. Получение клонов этих мутантов достигалось последовательными «пересевами» (молекул!) — переносом небольшой части инкубационной смеси, в которой процесс репликации завершился, в новую инкубационную смесь, содержащую фермент и трифосфорибонуклеотиды УТФ, ЦТФ, АТФ, ГТФ. Оказалось, что достаточно попадания в новую среду одной молекулы мутанта для образования нового клона популяции таких же молекул. В работе [460] было показано, что после 74 переносов можно выделить мутант, у которого скорость репликации в 15 раз превышает скорость репликации исходной, «родительской» РНК Qp-фага. Такое увеличение этой скорости, по-видимому, обусловлено уменьшением длины молекулы РНК — вместо 3600 нуклеотидных остатков в исходной РНК в мутанте их оказалось лишь 550. (В мутантных молекулах РНК для обеспечения высокой скорости размножения не требуется большинства последовательностей нуклеотидов, «осмысленных» предшествующей эволюцией фага).
Затем авторы цитируемой выше работы исследовали возможность возникновения в условиях молекулярного дарвиновского эксперимента молекул-мутантов с определенными новыми свойствами (т. е. молекул, отличающихся не только скоростью репликации). Был получен мутант, способный быстрее реплицироваться при меньших концентрациях субстратов (например, ЦТФ). Нуклеотидный состав и размер этой молекулы-мутанта примерно такие же, как и у исходной формы. Следовательно, отбор привел к изменению последовательности нуклеотидов в РНК • Как оказалась, ее изменение происходит при меньших концентрациях не только ЦТФ, но и других нуклеотидтрифосфатов. Быть может в результате изменения последовательности нуклеотидов изменилась вторичная структура мутантных молекул РНК, что привело либо к взаимодействию последних с ферментом, либо к увеличению активности фермента посредством алло- стерических механизмов. Затем были получены мутанты, способные к репликации в присутствии веществ, нарушающих репликацию «нормальной» РНК, — сначала авторы обнаружили мутант, чувствительный к концентрации АТФ, (мутант У-8), а затем осуществили серию переносов (пересевов) в среды, содержащие ингибиторный аналог АТФ (туберцидинтрифосфат (ТуТФ), содержащий в положении 7 пуринового кольца вместо азота углерод). В результате возник мутант У-9 вдвое быстрее реплицирующийся в присутствии ТуТФ, чем У-8.
Таким образом, в этих опытах показана решающая роль кинетических факторов в эволюции — побеждают в отборе те мутанты, скорость размножения которых .в данных условиях выше.
Глава 4
ФЕРМЕНТАТИВНЫЙ КАТАЛИЗ
Конечный результат биохимического этапа биологической эволюции — создание предельно совершенных катализаторов — ферментов. Обеспечение ферментами прохождения процесса по наиболее «быстрому» маршруту, путем отбора конформационно-лабильных полипептидных молекул. Возможность синхронизации отдельных конформационных циклов макромолекул ферментов. '
Ускорение диффузии и механо-химические преобразования энергии как результат этой синхронизации.
Итак, в естественном отборе выживают матричные полимерные макромолекулы, способные к более быстрому результирующему конвариантному воспроизведению. В этом состоит кинетический принцип, критерий естественного отбора. Как подчеркивалось выше, кинетическое совершенство — сложное понятие, и на степень такого совершенства влияет не только скорость матричного воспроизведения, но и скорость синтезов мономеров, а также кинетические («барьерные») механизмы сохранения уже синтезировании х макромолекул, кинетические механизмы осуществления реакций, обеспечивающих энергией «противотермодинамиче- ский» ход биологической эволюции.
Все аспекты кинетического совершенствования в ходе эволюции определяются эволюционным совершенствованием белков, полипептидов, т. е. полимеров, состоящих из достаточно разнообразных в химическом отношении мономеров (аминокислот).
Та или иная биохимическая реакция достигает в ходе эволюции необходимой скорости, целесообразно (в соответствии с нуждами данного этапа существования) регулируемой различными факторами, только при эволюционном совершенствовании соответствующего фермента.
Следовательно, эволюция аппарата матричного воспроизведения, процессов энергетического обеспечения (фотосинтез, гликолиз) и аппарата синтеза белка — есть эволюция ферментов.
Рассмотрим поэтому, пользуясь, как и раньше, методом анализа предельно совершенного этапа эволюции, общие свойства ферментов. При этом мы будем иметь в виду множество книг и обзоров, посвященных частным и общим вопросам ферментативного катализа [25, 29, 54, 56, 89, 90, 116, 176, 223, 238, 297, 302, 329, 395, 405, 436, 445]. Наш эволюционно-дедуктивный подход может способствовать выявлению нерешенных вопросов.
Для этого сформулируем эволюционные задачи, из которых следует необходимость высоко совершенных ферментов; учтем физические и химические свойства материалов, образующие биологические системы; попробуем представить себе, что получилось бы при неограниченно большом времени совершенствования ферментов, и, наконец, сравним полученную нами картину с действительной.
Эволюционные задачи мы уже неоднократно формулировали. Кратко их можно определить так: достижение предельно возможного биологического (кинетического) совершенства. В качестве материала, подлежащего эволюционному совершенствованию, мы уже выбрали полипептиды. Итак, необходимы белковые катализаторы, способные катализировать множество различных химических процессов и изменять свою каталитическую активность при взаимодействии с веществами-регуляторами, причем по принятому условию обе эти способности должны быть предельно совершенными. Все менее совершенные считаются погибшими в ходе эволюции.
Уточним сначала смысл основных понятий. Что значит «катализировать»? Меня не смушают при постановке этого наивного вопроса великие имена Я. Берцелиуса или В. Оствальда. Вовсе не лишнее раз в 50—100 лет возвращаться к таким вопросам. Катализ — это ускорение одной из в принципе многих возможных реакций, в которых опять же в принципе способны участвовать реагенты. В закрытых системах катализатор ускоряет как прямую, так и обратную реакции. В открытых системах катализатор определяет направление превращения веществ, которые в отсутствие катализатора вообще могут не взаимодействовать друг с другом с заметной скоростью. В этих системах катализатор может ускорять превращение лишь в одну сторону. Такая роль катализаторов следует из концепции маршрутов реакций Хориути [317, 401] и М. И. Темкина [286]. Вещество А может превратиться через целый ряд промежуточных реакций в вещество X — термодинамически наиболее стабильный конечный продукт. В достаточно сложной системе, содержащей множество различных веществ, превращение А в X может идти множеством путей— маршрутов. В принципе, только один маршрут «самый быстрый». Однако высокая итоговая скорость прохождения по маршруту отнюдь не обязательно означает высокую скорость на всех этапах пути. Вполне вероятно медленное, затрудненное прохождение по начальным этапам маршрута, оказывающегося в конце концов самым быстрым. Для некатализируемых реакций верен принцип Хиншелвуда, а именно, спонтанное прохождение их в конечном итоге по самому быстрому маршруту. Однако принцип Хиншелвуда отнюдь не обеспечивает высокой скорости реакции, не гарантирует самого быстрого из вообше возможных маршрутов (имеется в виду система без катализатора). Внесение в систему катализатора увеличивает разнообразие возможных маршрутов, среди которых оказывается и наиболее быстрый. На самом же деле, задачи ферментов и сложнее — речь идет не только об ускорении превращения А в X, но и о накоплении определенного промежуточного продукта Р. Такой продукт не обязательно «лежит» на самом быстром маршруте. Следовательно, катализатор — вещество, так изменяющее возможные маршруты реакций, что среди них оказывается маршрут, проходимый быстрее всех, возможных в отсутствие катализатора. А дальше работает, по существу, статистический принцип Хиншелвуда. Такое понимание роли катализаторов означает, что если бы и в отсутствие катализатора можно было обеспечить маршрут реакций, идентичный самому быстрому в его присутствии, то мы полностью имитировали бы каталитический процесс. Система реакций, в которых осуществляется наиболее быстрый маршрут, называется конгруентной. В соответствии с таким представлением о катализаторе последний лишь обеспечивает необходимый набор конгруентных промежуточных реакций.
Это понимание роли фермента является чисто химическим и не требует наличия каких-либо специфических физических механизмов ускорения катализируемых процессов. Для допущения необходимости подобных механизмов нужны серьезные, в том числе эволюционные обоснования. Посмотрим далее, найдутся ли такие обоснования, а пока попробуем конкретизировать «маршрутную» роль ферментов.
Что значит сделать возможным новый маршрут? Это значит обеспечить прохождение процесса через ряд новых промежуточных реакций. Новые промежуточные стадии не осуществляются без катализатора, и поэтому соответственные промежуточные вещества (состояния) нестабильны вне комплекса с катализатором. Отсюда следует весьма важный вывод — сколько-нибудь совершенный катализ возможен лишь при условии непрерывного комплексообразования реагирующих веществ с катализатором (ферментом). Это значит, что в пределе ни на одной из промежуточных стадий катализируемого процесса промежуточные вещества не должны выходить из комплекса с ферментом. Отсюда формулируется первое условие в «техническом задании» на эволюционное изготовление ферментов. Молекулы ферментов должны образовывать комплексы с исходными веществами и всеми промежуточными формами реагирующих веществ. Для появления комплексов с последовательно возникающими промежуточными веществами в катализируемом процессе макромолекула белка-фермента должна обладать способностью к последовательному изменению своей конформации. Конформационная лабильность служит условием последовательного изменения «реакционного клубка», т. е. образования в активном центре необходимого очередного сочетания химически активных боковых радикалов определенных аминокислот. Так и обеспечивается осуществление особо быстро проходимого маршрута в присутствии фермента.
Как отмечалось, по происхождению и вследствие единственно возможного способа эволюционного совершенствования посредством мутаций ферменты представляют собой белки, полипептиды. Следовательно, именно макромолекулы ферментов должны быть конформационно лабильными. Конформационной лабильностью могут обладать полипептидные цепи, не сшитые жесткими (например, дисульфидными) связями. Макромолекула фермента должна состоять из нескольких консолидированных частей, подвижно (на шарнирах) связанных друг с другом.
Такую макромолекулу можно макроскопически моделировать, например, системой шаров, соединенных шарнирами и пружинами. Следовательно, для надлежащей функции она должна представлять собой машину - -устройство с весьма ограниченным целесообразным набором степеней свободы — направлений перемещений частей друг относительно друга [318].
Чем замечательна такая машина? Все ее своеобразие — в молекулярных размерах[5].
Мы часто в последнее время произносим словосочетание «молекулярная машина», не осознавая его экстравагантности. «Нормальная» машина — устройство, в котором тепловое движение составляющих ее атомов (деталей) не играет никакой роли. Машина обычно вполне макроскопична. Молекулярная машина существует в оглушительном тепловом шуме, «целесообразные» движения ее деталей происходят среди теплового беспорядка и являются статистическим итогом разнонаправленного «броуни- рования» [136—138]. Почему же мы говорим о макромолекуле белка как о машине? Потому, что в силу структурных ограничений большая часть взаимных перемещений «кусков» макромолекулы друг относительно друга невозможна и сама она совершает броуновское движение как целое. Лишь в некоторых функционально значимых направлениях тепловые флуктуации приводят к изменениям конформации, изменениям взаимного расположения частей макромолекулы. В макромолекуле фермента, не соединенной с субстратом, эти движения равновероятны в двух направлениях — «туда» и «обратно» (они представляют собой флуктуационные конформационные колебания), тогда как в макромолекуле, связанной с превращаемым субстратом, движения «туда» и «обратно» неравноценны. Например, при движении какой-либо функциональной группы полипептидной цепи «туда» осуществляется реакция, сопровождающаяся необратимым изменением субстрата (его свободная энергия уменьшается и выделяется тепло), а при движении «обратно» реакция не идет (без сопряженного подвода энергии).
Так осуществляется выбор нужного направления флуктуаций, использование тепловой энергии для направленного превращения. Нас обычно преследует демонобоязнь — боимся мы демонов Максвелла!
Действительно, по первоначальному замыслу Максвелла, демоны должны были открывать дверцу в перегородке для молекул газа, летящих с большей, чем средняя, скоростью, имеющих избыток кинетической энергии, и закрывать эту дверцу перед медленными молекулами. Так демоны призваны были разделять газ, находящийся в тепловом равновесии на две части — холодную н горячую. Давно уже Сциллард и Бриллюэн показали принципиальную невозможность сушествования таких демонов. Затраты свободной энергии на различение молекул в точности равны ожидаемому возрастанию свободной энергии (за счет уменьшения энтропии системы) при сортировке молекул.
Нужно поэтому особо подчеркнуть, что в данном случае мы имеем дело с неравновесной системой, идет превращение вещества, при котором выделяется энергия, причем выделяется она необратимо. Следовательно, принципиальных запретов для окупаемого затратами энергии направленного использования флуктуаций нет.
Молекулярные демоны—анизотропные структуры — в принципе могут работать за счет энергии, освобождаемой в катализируемой реакции.
Механизм нашего демона также вполне рационален — отобранные в ходе длительной эволюции, синтезируемые с затратой свободной энергии из сопряженных процессов, структуры макромолекул таковы, что конформационные движения в них резко и целесообразно ограниченны. Таким образом, предполагаемое своеобразие макромолекулы фермента в том, что она представляет собой машину, работа которой невозможна без тепловых флуктуаций, хотя энергия, обеспечивающая весь ее рабочий цикл, поступает из нетепловых источников — тратится химический потенциал превращаемых веществ (см. [328, 337, 445]).
Впрочем, для приведенной выше картины работы макромоле- кулярной машины не требуется каких-либо механизмов накопления флуктуаций; не предполагается и ускорение катализируемого процесса за счет энергии, накопленной тем или иным способом в макромолекуле фермента ‘. Например, в результате «рекуперации» энергии, выделяющейся в ферментативной реакции [136—138]. Любая конгруентная реакция, т. е. превращение веществ без участия фермента (если бы ее удалось осуществить) шла бы с такой же скоростью, как и при помощи фермента. Медленные реакции и там и тут медленные. Просто без фермента эти процессы в данной системе вообще не идут таким маршрутом. Итак, ферменты могут не ускорять отдельные элементарные
* Кажется поивлекательной возможность «собирания флуктуаций» со многих степеней свободы в активном центре фермента в качестве механизма ускорения катализируемых реакций [400]. Нам не удалось показать реальность этой гипотезы [328].
реакции, а «просто» позволяют осуществляться таким промежуточным элементарным стадиям, которые невозможны без них, они ускоряют образование конечных продуктов, делая возможным осуществление новых маршрутов. Это звучит, конечно, парадоксально, и тем не менее, вероятно, соответствует действительности.
Здесь уместно отметить, что представление о ферментативных реакциях как об очень быстрых — привычное заблуждение. Ферментативные реакции (правильнее, процессы) обычно гораздо быстрее неферментативных. Но неферментативные, как было показано, нельзя сравнивать с ферментативными — они идут по другим маршрутам. Абсолютные скорости ферментативных процессов (как и конгруентных им неферментативных) вовсе не велики.
В самом деле, ферментативные циклы — превращения субстрата в продукт — совершаются за 1 —10_3 сек, т. е. идут очень медленно. За это время в элементарном акте взаимодействия молекул, атомов, радикалов происходит 1013—1010 соударений и только одно из 1013—1010 эффективно — где уже тут говорить о высокой скорости![6]
Поэтому актуальны «парадоксальные» вопросы — в чем причина столь медленной работы ферментов? Почему столь малы числа оборотов большинства ферментов? Что ограничивает эти числа? Скорее всего — малая частота больших тепловых флуктуаций. В самом деле, частота появления флуктуаций величиной 30 кТ на каждую степень свободы составляет примерно 1 сек-1, 25 кТ — 100 сек-1, 20 кТ — 104 сек-1. А такие флуктуации и нужны для преодоления потенциальных барьеров при взаимодействии реагентов. Таким образом, ферментативные процессы протекают медленно из-за малой частоты больших тепловых флуктуаций.
Подведем итог. Белки — ферменты должны претерпевать последовательные изменения конформации, что обеспечивает прохождение катализируемого процесса по наиболее быстрому маршруту. Эти последовательные изменения конформации осуществляются за счет флуктуации тепловой энергии и энергии, выделяющейся в катализируемом процессе. Общее для всех ферментов «узкое место» — малая частота больших флуктуаций.
В табл. 2 приведены данные о числе оборотов, т. е. о числе полных циклов ферментативного процесса за 1 сек для разных ферментов. Как мы видели, для большинства ферментов числа оборотов очень малы — от нескольких единиц до тысяч единиц в 1 сек (и это при 1013—1010 элементарных актов-столкновений в 1 сек).
ТАБЛИЦА 2
КАТАЛИТИЧЕСКАЯ АКТИВНОСТЬ ФЕРМЕНТОВ В ЧИСЛЕ ОБОРОТОВ
Фермент
Источник получения
Субстрат
Число оборо- тов, сек~*
«Медленные» ферменты
Пепсин
Желудочный сок
Ацетил-/-фенилаланил-
2-10-'
свиньи
дийодтирозии
То же
То же
Ацетил-/-феиилаланшЯ-
1,19-10-2
фенилалаиин
»
»
Карбобензокси-1 - г лутамил-
1,09-10-8
/-тирозин
Т рипсин
Поджелудочная же
Этиловый эфир бензоил-/-
22
леза
аргинииа
То же
То же
Бензоил-/-аргинин
►Ч
1
О
ОО
•^н
Химотрипсии
»
Этиловый эфир ацетил
7,МО"1
триптофана
То же
»
Метиловый эфир бензол-/-
1,1-Ю-1
лейцина
Тромбин
Кровь человека
/7ара-нитрофеииловый
2,2-10-1
эфир N-карбобензокси-/-
тпрозина
То же
То же
Лара-нитрофеииловый
7,2-Ю'1
эфир N-карбобензокси
/-лизина
«Средние» ферменты
Алкогольдег идро-
Дрожжи
Этаиол
250
геназа
То же
То же
Ацетальдегид
416
Г лкжозо-6-фосфат-
Глюкозо-6-фосфат
1500
дегидрогеназа
Сукцинатдегидроге-
Митохондрии серд
Сукцннат
184
наза
ца быка
Алаиинаминот ранс-
Сердце свиньи
Аланнн
680
фераза
Г ексокиназа
Дрожжи
Глюкоза
1300
Фосфофруктокииаза
Мышцы кролика
Фруктозо-6-фосфат
1800
а-амилаза
Поджелудочная
Амилоза
1035
железа
Альдолаза
Животные, высшие
Фрукт озо-1,6-дифосфат
63
растения
Ппруватдекарбокси-
Дрожжи
Пируват
150
лаза
Карбокснпептидаза
Поджелудочная же
Г 1:ппурил-/-лизин
220
В
леза
То же
То же
Г иппурил-/-аргинин
105
Фермент
Источник получения
Субстрат
Число оборотов, сек~1
«Быстрые» ферменты
Каталаза
Печень быка
н202
9-104
Ацетилхолинэстера
Электрический
Ацетилхолин
за
угорь
Угольная ангидра
Эритроциты чело
Н2СОа
2,3 ■ 104
за Ь
века
Угольная ангидра
—
—
3,6•105
за с
Уреаза
Бобы капавалии
Мочевина
1,7-10^
Инвертаза
Дрожжи
Сахароза
1,3-Ю4
Т риозофогфз тизо-
Мышцы кролика
3-фосфоглицериновый аль
8,5-Ю3
мераза
дегид
Такую картину нельзя объяснить биологической целесообразностью— тем, что большие скорости и не были нужны в эволюции. Давление естественного отбора не терпит пустоты. Совершенство ферментов, в силу сказанного ранее, должно быть предельно возможным. И если их числа оборотов столь малы, значит, они ограничены физическими причинами.
Мы видим, однако, в табл. 2 и несколько ферментов, у которых числа оборотов на несколько порядков выше, чем у остальных— это ацетилхолинэстераза, каталаза, угольная ангидраза. Они выполняют функцию быстрого разрушения небольших молекул— ацетилхолина, перекиси водорода, угольной кислоты. Из сказанного выше мы должны вывести, что эти ферменты отличаются от основной группы принципом работы. Возможно, что катализируемые ими реакции происходят в один акт, и сами быстрые ферменты не должны претерпевать значительных кон- формационных изменений в каталитическом цикле. Такие ферменты, по-видимому, наиболее близки к небиологическим гетерогенным катализаторам.
Протеолитические ферменты (см. общие сведения [217]). В табл. 2 приведены также сверхмедленные ферменты — протеа- зы, которые гидролизуют белки. Иные из них осуществляют всего один оборот за 103 сек. Чем это можно объяснить? Сложностью катализируемых процессов? Вряд ли. Процессы, катализируемые синтетазами, например, синтетазой жирных кислот, по числу отдельных стадий, необходимости взаимодействия большого числа молекул, согласованности действия разных субъединиц макромолекулы фермента нисколько не проще, а идут быстрее.
Можно предложить несколько объяснений этого странного явления.
Возможно, что причина медленности протеолиза — особые
свойства субстратов-белков. Нативная конформация белковых макромолекул «устроена» так, что возникающие в полипептидной цепи флуктуации энергии отводятся по «хребту» главных валентностей на поверхность глобулы и там рассеиваются. Такая канализированная теплопроводность обеспечивает особую устойчивость нативной макромолекулы белка. Протеолитический фермент, атакуя пептидную связь в нативной макромолекуле, должен дожидаться для ее разрыва значительно больших флуктуаций энергии, чем для разрыва такой же связи в денатурированной макромолекуле с нарушенной внутримолекулярной теплопроводностью. Существенно легче, быстрее должны разрушаться пептидные связи и в низкомолекулярных пептидах. Этого, как видно из таблицы, нет. Однако эффект резкого ускорения скорости протеолиза после денатурации белков широко известен. На этом эффекте основан метод характеристики степени нативности конформаций макромолекул белков. На этом же, в сущности, основана и кулинарная обработка мяса. Однако, несмотря на огромный исторический опыт человечества в денатурации белков для ускорения их протеолиза, достоверный механизм этого эффекта не выяснен. Принято думать, что ускорение протеолиза после денатурации белков-субстратов вполне объясняется увеличением доступности для ферментов, скрытых в нативном белке пептидных связей, гидролизуемых данной протеазой. Мне не кажется, что такое объяснение вполне обосновано. Необходимы тщательные кинетические исследования.
Более правдоподобно объяснение особой медленности — гидролиза пептидных связей, в том числе и низкомолекулярных пептидов, эволюционными причинами. В эволюции не было необходимым возникновение высокоактивных протеаз, поскольку их активность лимитирована медленностью диффузии их естественных субстратов — белков (см. главу 5, с. 81).
Общая картина функционирования ферментов представляется такой. «Голодные» макромолекулы фермента в растворе без субстратов (или встроенные в какую-либо структуру) подвергаются действию тепловых флуктуаций — испытывают случайные изменения конформации, «щелкают челюстями», т. е. иными словами, в результате анизотропии теплопроводности макромолекул тепловые удары молекул среды по всей поверхности макромолекулы фермента преобразуются в высоко амплитудные конфор- мационные движения ее частей друг относительно друга.
При взаимодействии фермента с молекулами субстрата образуются фермент-субстратные комплексы. Теперь анизотропия теплопроводности макромолекул обусловливает подведение флуктуаций энергии в активный центр, где эта энергия тратится на преодоление потенциальных барьеров, препятствующих осуществлению реакций. Преодоление этих барьеров осуществляется за счет высоко амплитудных конформационных движений (колебаний) макромолекулы фермента, которая перекусывает,разгрызает, или наоборот слепляет молекулы субстрата. Реакции могут состоять из многих элементарных стадий. Каждой стадии соответствует своя конформация макромолекулы, свое время ожидания должной по величине флуктуации энергии, свой вид конформационных движений. По завершении всех промежуточных превращений возникает конечный продукт, не образующий достаточно прочного комплекса с ферментом и этот комплекс диссоциирует. Длительность существования так®ге фермент- субстратного комплекса определяется в основном суммарным (по всем этапам) временем ожидания нужных флуктуаций. Время это велико, чем и объясняется непреодолимая в эволюции медленность ферментативных процессов. Конформационные движения макромолекул ферментов при осуществлении ими каталитических реакций должны приводить кувеличению скорости диффузии макромолекул, «работающих» ферментов по сравнению с неработающими.
Возможная синхронность конформационных движений макромолекул в ансамблях и ее следствия. Представим себе теперь большое число таких макромолекул фермента в растворе, содержащем соответствующий субстрат. По существу речь здесь идет
множестве генераторов механических колебаний, распределенных в среде, в которой они совершают циклические изменения конформации. Поскольку эти генераторы полностью идентичны, обязательно должно происходить их взаимодействие, приводящее к образованию упорядоченных в пространстве синхронно колеблющихся ассоциаций ферментных макромолекул. В результате в растворе должны образоваться структуры типа жидких кристаллов отличающиеся от обычных своей эфемерностью ■— они представляют собой ансамбли лишь работающих макромолекул фермента. Мне представляется появление таких ассоциаций обязательным следствием способа функционирования ферментов. Иными словами они образуются не потому что это эво- люционно необходимо а потому что не могут не возникнуть (см. гл. 8). Однако их формирование, обусловленное физико-химическими механизмами, может стать причиной ряда биологических следствий. Существование таких ансамблей макромолекул, синхронно циклически изменяющих свою конформацию, должно приводить к некоторым макроскопическим эффектам. Среди них наиболее вероятным представляется не только резкое ускорение диффузии, но и возникновение около таких ансамблей более или менее регулярных потоков жидкости, возбуждаемых и поддерживаемых синхронными движениями ансамблей макромолекул. (Нужно помнить, что в результате осуществления катализируемых ферментами реакций выделяется энергия, окупающая все расходы и достаточная для совершения разного рода работы[7].)
Эквивалентным эффектом является перемещение самих макромолекул, которое в силу синхронизации и синфазности конфор- мационных циклов отдельных макромолекул должно значительно превышать перемещение последних. Так, перемещение лодки- восьмерки с синхронно работающими гребцами, значительно превосходит перемещение отдельных лодок-одиночек, движущихся в разных направлениях и мешающих друг другу. Такой эффект— ускорение диффузии и перемещение в пространстве каталитических ансамблей — имеет огромное принципиальное значение и создает новое направление кинетического совершенствования в процессе эволюции.
Из возможных следствий взаимодействия макромолекул ферментов, осуществляющих конформационные колебания, или в общем случае, циклические изменения конформации, рассмотрим пульсации давления — звуковые волны. Диапазон чисел оборотов большинства ферментов соответствует слышимым звуковым частотам. Некую, пока еще фантастическую картину «музыкальных взаимодействий» биохимических систем, клеток, органов, и возможную роль этих взаимодействий в жизнедеятельности мы рассмотрим подробнее в главе 8 (см. [253, 337]). Возможно, наиболее важным следствием функционирования макромолекул ферментов посредством изменения своей конформации являются механо-химические преобразования энергии. Способность к таким преобразованиям не может остаться вне поля зрения естественного отбора. Начинается эволюционное совершенствование, образование специализированных, высокоэффективных систем биологической подвижности (см. гл. 9, с. 174).
Теплоустойчивость белковых макромолекул и анизотропия теплопроводности. Анизотропия теплопроводности макромолекулы белка делает эту молекулу особо устойчивой к разрушающему влиянию тепловых флуктуаций — они отводятся по «хребту» главных валентностей и разряжаются на концевых группах, способных совершать высоко амплитудные конформационные движения, или же на дефектах структуры. Можно представить себе такой ход тепловой денатурации белка, при котором первоначальное медленное из-за анизотропной теплопроводности нарушение нативной структуры, требующее преодоления очень высокого барьера, сменяется быстрым легко осуществляемым ее нарушением. Такая пороговая зависимость тепловой денатурации от температуры и будет восприниматься в опыте как проявление очень больших величин предэкспоненциальных множителей и больших энергий активации в уравнении Аррениуса для скорости денатурации.
Как скажется на теплоустойчивости макромолекулы фермента взаимодействие с субстратом? В разных случаях по-разному. В свободной макромолекуле фермента больше всего должны
быть подвержены тепловому разрушению связи, близкие к концевым группам, шарнирные группы и т. п., т. е. все области, где разряжаются тепловые флуктуации. При взаимодействии с субстратом большие тепловые флуктуации обеспечивают катализируемые превращения, и тогда можно ожидать повышения теплоустойчивости работающего фермента по сравнению с безработным. Вместе с тем в ходе каталитического акта может выделяться энергия, создающая дополнительную нагрузку на структуру фермента. В этом случае можно ожидать неоднозначной зависимости теплоустойчивости ферментов от концентрации субстрата — при малых концентрациях субстрата теплоустойчивость может падать, а при больших — расти.
Возможные способы регулирования ферментативной активности. В понятие совершенства функционирования катализаторов входит и совершенство изменения скоростей биохимических процессов. Поскольку их скорости определяются в основном наличием и активностью соответствующих ферментов, регулирование биохимических процессов может идти двумя путями: регулированием синтеза ферментов, т. е. регулированием преобразования генетической информации, записанной в полинуклеотидных цепях, в последовательность аминокислот в полипептидных цепях белков — ферментов и регулированием активности уже синтезированных ферментов. Оба эти пути — предмет многочисленных исследований и им посвящены фундаментальные работы и обзоры [11а, 25, 33, 42, 56, 89, 90, 176, 211, 245, 257, 258, 302, 310].
В связи со сказанным ранее, здесь все же следует подчеркнуть, что конформационная лабильность, машинный принцип функционирования макромолекулы фермента создают возможность для тонкой регуляции ферментативной активности.
Резюмируем сказанное в этой главе. Биологические катализаторы по своему происхождению и способу эволюционного совершенствования с необходимостью должны быть полипептидами, белками. Их каталитические свойства определяются строго специфическим соединением как с исходным субстратом, так и с промежуточными продуктами его превращения. Это достигается в большинстве случаев посредством закономерных обратимых конформационных изменений макромолекул ферментов. Работающие молекулы ферментов, возможно, образуют многомолекулярные ансамбли с синхронными конформациониыми движениями всех его членов. Следствием таких движений может быть активное перемешивание реакционной смеси, эквивалентное существенному ускорению диффузии. Эти ансамбли могут в ходе дальнейшей эволюции явиться началом формирования специализированных аппаратов активного перемещения в пространстве. Анизотропная теплопроводность может быть причиной как повышенной теплоустойчивости, так и повышенной устойчивости к протеолизу нативных молекул белка.
Глава 5
КИНЕТИЧЕСКИЙ СМЫСЛ И ВОЗМОЖНЫЕ СПОСОБЫ СТРУКТУРНОЙ ОРГАНИЗАЦИИ БИОХИМИЧЕСКИХ ПРОЦЕССОВ
Принципиальная осуществимость всех биохимичскш процессов в гомогенных растворах.
Необходимость структурной организации для обеспечения высокой скорости переключений многоэтапных биохимических превратный и для осуществления этих превраще-ий посредством наименьших количеств ферментов.
Обеспечение полиферментными комплексами t органеллах, окруженных избирательно проницаемыми мтб ранами, при активном, противоградиентном накачивании и.хосных веществ наивысшей возможной суммарной скорости м-огсэтапных биохимических превращений и наибольшей сксоости изменения их режима.
В предыдущей главе мы рассматривали общие механизмы функционирования ферментов, не обращая внимания на диффузные ограничения обеспечения ферментов субстратами. На самом деле дальнейшее эволюционное совершенствование каталитической активности ферментов становится с некоторого момента бесполезным и, следовательно, не происходит из-за диффузионных ограничений (см. [208]).
В самом ли деле диффузия ограничивает скорости биохимических процессов? Убедиться в этом можно лишь посредством количественных оценок. Такая оценка проведена при содействии
С. Э. Шноля нами [343].
В этой статье оценена интенсивность дифсузяонных потоков субстратов в биохимических процессах, идусих без активного перемешивания для двух случаев моделей гои диффузии субстрата к каталитически активной поверхностна 2\ при диффузии субстрата в растворе катализатора-фермента. Первый случай соответствует расположению фермента, на какой-либо внутриклеточной мембране или адсорбции фермента на наружной поверхности клетки (например, в случае так называемого «пристеночного пищеварения» [294]). Второй случай еоогветствует процессам типа гликолиза.
Модель 1. Пусть на одном конце трубки длиной I поддерживается постоянная концентрация S0 данного субстрата. На другом конце представим себе тонкий слой молек л фермента, понижающих S0 до Si. Максимально возможный диффузионный поток получится при Si = 0. Он равен:
/’max ^ DSq — ,где D — коэффициент диффузии, 0 — площадь поперечного сечения трубки. Для D~ 10~6 см2/сек, 50 — 103 М (10~в моль/см3) получим /т.ах в зависимости от I и а = а2, приведенную в табл. 3.
ТАБЛИЦА 3
МАКСИМАЛЬНЫЙ ДИФФУЗИОННЫЙ ПОТОК СУБСТРАТА /тах В МОЛЬ/СЕК К ФЕРМЕНТАТИВНО-АКТИВНОИ ПОВЕРХНОСТИ
1
а
1
ю->
Ю-»
ю-*
Ю-‘
1
10-12
10-U
10-16
Ю-!8
10-20
10-*
кг11
Ю-is
10-!5
10~17
10-19
1(Г2
10-10
Ю-12
1(Г14
10-16
Ю-18
10-3
1<Г9
10-11
10-13
10-15
Ю-17
10-4
10“8
Ю-ю
Ю'12
Ю-м
10-!в
Посмотрим теперь, какой каталитической активностью должен обладать фермент, чтобы обеспечивать переработку всего субстрата, поступающего с диффузионным потоком.
Пусть молекулы фермента расположены на площадке а в один слой. При концентрации фермента Е0 моль/см3 на площадке в
см2 расположится n=(E0N)*!* молекул фермента (N—число Авогадро).
Таким образом, каждая молекула фермента должна перерабатывать j-N/(E0N)% молекул субстрата в секунду, чтобы обеспечивать максимальный диффузионный поток. Следовательно:
DSBaN
kn = 7Г ■
“ ■ I(E„N) h
В зависимости от диффузионного пути И, концентрации фермента Е0 получаются величины кг (числа оборотов в сек-1 см. гл. 4, с. 70), необходимые для переработки в одном слое фермента всего приносимого диффузионным потоком субстрата к каталитически активной поверхности, приведенные в табл. 4.
ТАБЛИЦА 4
ВЕЛИЧИНА КАТАЛИТИЧЕСКОИ АКТИВНОСТИ АДСОРБИРОВАННЫХ НА МЕМБРАНЕ
ФЕРМЕНТОВ-Kj В СЕК-1, ОГРАНИЧИВАЕМЫЕ ДИФФУЗИОННЫМ ПОТОКОМ
СУБСТРАТА В ЗАВИСИМОСТИ ОТ I И Е0 (ПРИ Д~ 10-8 CM2/CEK, S„~ 10-3 М
(10—5 МОЛЬ/СМ3)
1, см
£0, м
1 ю-1
ю-*
ю-*
10-*
10~5
2-101
2-102
2 -103
2-104
2-105
10-7
4-102
4-103
4-104
4-105
4.10е
Если субстратом являются большие молекулы, например, белка в случае протеолиза, то D~10-7 см2/сек и S0 —10-5 М (10-8 моль/см3) предельные величины k2 будут соответственно ниже (табл. 5).
ТАБЛИЦА 5
ВЕЛИЧИНЫ КАТАЛИТИЧЕСКОЙ АКТИВНОСТИ АДСОРБИРОВАННЫХ НА МЕМБРАНЕ ФЕРМЕНТОВ, ТИПА ПРОТЕИНАЗ, ОГРАНИЧИВАЕМЫЕ ДИФФУЗИОННЫМ ПОТОКОМ СУБСТРАТА В ЗАВИСИМОСТИ ОТ I И Е„
I, см
10, м
1
ю-»
ю-*
ю-«
ю-<
10-7
4-10-'
4
4-101
4-102
4-103
1и~5
2-ID-2
2-Ю-1
2
2-10
2 • 102
Мы видим, что процессы, аналогичные внеклеточному пищеварению, очень сильно лимитируются диффузией. Действительно, к ферменту — протеиназе, адсорбированному на какой-либо поверхности при реально достижимых концентрациях белков при пищеварении диффузионный поток приносит одну молекулу субстрата один раз на протяжение времени порядка секунд. Нет никакого смысла в эволюционном совершенствовании протеиназ сверх этого лимита — более активные ферменты — бесполезны и, следовательно, не могут возникнуть в ходе естественного отбора. Так что наши рассуждения ib главе 4 о возможной причине парадоксально низкой каталитической активности протеиназ, /наверно, действительно должны быть сведены к объяснению диффузионными ограничениями.
Рассмотрим теперь возможные диффузионные ограничения биохимических процессов, идущих при диффузии субстратов в растворах ферментоз.
Модель 2. Представим себе клетку в виде трубы длиной I, сечением а, заполненную раствором фермента концентрации Е0. Пусть на одном конце трубки поддерживается постоянная концентрация субстрата S0, а на втором конце концентрация субстрата S,. Примем, как и раньше, что скорость превращения субстрата в продукт задается формулой Михаэлиса — Ментен:
k2E S
v = т—г~т . (Кт — константа Михаэлиса)
т г *
В стационарных условиях диффузионный поток субстрата уравновешивается в каждой точке объема ферментативным процессом, т. е.:
d2S k*SE,
РИС. 9. Изменение концентра ции субстрата при его диффу зии в растворе фермента

Ясно, что коэффициент диффузии D и величина k2 связаны друг с другом. Установить связь между ними прямо из условия S(x) > >0 не удастся, т. к. нельзя аналитически решить уравнение с граничными условиями S(0)=S0, dS/dx|*=i=0. Попытаемся хотя бы грубо оценить необходимую величину кг через D. Если предположить, что S{x)^Km, т. е. молекулы фермента в достаточной мере обеспечены субстратом, то кривая S(x) на рис. 9 пройдет ниже кривой Si(a:) и выше кривой S2(x), которые удовлетворяют
d2Si , /0 k2E0 d2S2 k2E0 соответственно уравнениям и — —и т
же граничным условиям, что и S (х).
Требование, чтобы концентрация S(x) была больше 0, с необходимостью влечет S^xJX). Отсюда несложными выкладками получаем неравенство: k <_40S^_ l2E0
Важно подчеркнуть, что в отличие от случая с диффузией к каталитической поверхности (с. 78), величина &2 оказывается обратнопропорциональной квадрату диффузного пути I.
Теперь мы можем провести численную оценку. Примем D~ ~ 10_6 см2/сек S0 — 10—4 М (10-7 моль/см3) и вычислим величины &2, лимитируемые диффузией субстратов для растворенных ферментов.
ТАБЛИЦА 6
ВЕЛИЧИНЫ КАТАЛИТИЧЕСКОЙ АКТИВНОСТИ ВНУТРИКЛЕТОЧНЫХ ФЕРМЕНТОВ К2 В СЕК-1, ОГРАНИЧИВАЕМЫЕ ДИФФУЗИОННЫМ ПОТОКОМ СУБСТРАТА В ЗАВИСИМОСТИ ОТ I И Е,
Ео, м
1, см
ю-1
10“г
10-*
I0-*
10-5
4-10-3
4-10-1
4-101
4-103
10-0
м
1
О
4
4-102
4-104
10-7
4-10-1
4-101
4-103
4-105
Полагая характерными размеры клетки порядка 10-3 см (10 |я), концентрации субстрата порядка 10-4 М, концентрации ферментов 10-5—10-7 М мы получим для kz величины порядка 101—Ю3. Именно этому диапазону соответствует каталитическая активность ферментов, функционирующих во внутриклеточных растворах (типа ферментов гликолиза). Это «средние ферменты» в нашей классификации (см. табл. 2, гл. 4, с. 71).
В то же время для внутриклеточного процесса типа протео- лиза (все это в отсутствие активного перемешивания) реальна концентрация субстрата 10~6 м, а фермента 10_в—10-7 ми D~ ~ 10-7 см2/сек) диффузионно-лимитируемая величина оказывается равной 10-1—1 сек-1.
Как видно при сравнении табл. 5 и 7, при характерных размерах клетки порядка 10-4 см различия в диффузионно-лимити- руемой каталитической активности ферментов, адсорбированных на мембране и свободно растворенных в «протоплазме», нивелируются. Следовательно, для мелких клеток типа микробных адсорбция ферментов не дает значительных преимуществ. Возможно, этим и определяются сами эти характерные размеры микробных клеток.
Может возникнуть сомнение в правильности приведенных оценок — там были заданы в качестве самых низких концентраций ферментов порядка 10~7 М. Казалось бы можно совершенствовать работу каждой молекулы фермента, увеличивать kz, еще сильнее уменьшая концентрацию фермента. Оказывается — нельзя. Предельно низкая концентрация ферментов определяется малыми размерами клеток. В самом деле в клетке не может быть меньше одной молекулы данного фермента. Одной молекулы, конечно, мало — ненадежно. Легко вычислить, что при характерных размерах клетки 10~4—10-3 см и наличии в ней 10‘— 102 молекул данного фермента, концентрации ферментов не могут быть ниже 10~6—10"8 М.
Так или иначе, эти, как ясно, весьма приближенные оценки иллюстрируют диффузионные ограничения даже одноферментных биохимических реакций.
Эти ограничения становятся особенно существенными в многоэтапных, многоферментных биохимических превращениях. Действительно, в тех многоэтапных превращениях, в которых важен лишь конечный продукт, чем меньше времени расходуется на промежуточные стадии и чем меньше необходимое количество ферментов, тем лучше. Однако, если промежуточные превращения происходят в бесструктурном гомогенном растворе, путь каждой данной молекулы по всей цепи превращений становится очень долгим.
В многоэтапных превращениях, в которых важен лишь конечный продукт, диффузия промежуточных продуктов в объем (клетки) ухудшает кинетические характеристики системы в целом. Диффузия, как и в одноферментных системах, лимитирует
скорости промежуточных реакций. Кроме того, для обеспечения должной скорости промежуточных реакций, т. е. создания соответственно высоких концентраций промежуточных субстратов, оказываются необходимыми большие количества исходных субстратов. На это может расходоваться значительная часть исходного субстрата. И, возможно, самое главное, резко возрастает инерционность системы, время, необходимое для переключения с одного режима на другой. На каждом этапе продукты, возникшие на предыдущем ферменте, диффундируют в среду. Для того, чтобы дальнейшее превращение пошло со значительной скоростью, концентрация образовавшегося продукта, т. е. субстрата очередного фермента, должна повыситься до некоторой величины. В зависимости от объема системы на это нужно большее или меньшее время. Такая задержка происходит на каждом этапе. Суммарная задержка пропорциональна числу промежуточных стадий. Ясно, что подобная биохимическая система будет очень инерционной и малопригодной, например, для энергетического обеспечения быстрых физиологических процессов.
Другой существенный недостаток гомогенной системы — неэкономное расходование ценного ферментативного вещества. Для обеспечения необходимой скорости превращения веществ нужна большая концентрация соответствующих ферментов во всем реакционном объеме.
Таким образом, давление естественного отбора должно привести к выработке приспособлений, уменьшающих задержку на промежуточных стадиях и обеспечивающих более экономное использование ферментов. Эти приспособления — надмолекулярные комплексы, структуры, в которых катализаторы промежуточных превращений находятся на малых расстояниях или даже определенным образом ориентированы друг относительно друга.
Осуществление полиферментных превращений посредством надмолекулярных полиферментных комплексов, расположенных внутри органелл, отграниченных от окружающей их среды избирательно проницаемыми мембранами, обеспечивает дополнительные кинетические преимущества. Условием эффективного функционирования таких органелл является относительно незатрудненное проникновение в них исходных веществ и свободный выход конечных продуктов при малой проницаемости их поверхностной мембраны для промежуточных продуктов.
В этом случае скорость промежуточных превращений зависит от концентрации первого субстрата, поступающего из среды.
Ясно, что эффективным способом ускорения промежуточных превращений может служить увеличение концентрации превращаемых веществ внутри органеллы посредством активного транспорта, т. е. противоградиентной накачки веществ при сопряженной трате энергии, например, выделяющейся при гидролизе АТФ.
Таким образом, активный транспорт, сопряженный гидролиз АТФ, обычно анализируемые с чисто термодинамических позиций, обеспечивают большую скорость многоэтапных биохимических превращений, увеличивают их кинетическое совершенство. В этом, вероятно, и состоит основной биологический смысл процессов активного транспорта.
Мне хочется специально подчеркнуть последнее обстоятельство, так как обычно полагают, что связь между кинетическими и термодинамическими характеристиками химических и биохимических систем отсутствует. Здесь мы можем даже вывести количественную связь между затратами свободной энергии при активном транспорте и скоростью превращения вещества внутри органеллы.
Пусть v — скорость превращения субстрата в продукт, a AF — изменение (затрата) свободной энергии при противоградиентном переносе данного субстрата к месту его дальнейшего превращения; тогда, из уравнения Михаэлиса (см. с. 79) и соотношения:
S = Su exp (Д F/RT) получим:
k2S0E exp (AF/RT)
V = S0 exp (AF/RT) + Ku ’
где E — концентрация фермента, Км— константа Михаэлисаг равная половине концентрации субстрата, обеспечивающей максимальную скорость, S0 — концентрация субстрата вне органеллы, 5 — его концентрация внутри органеллы.
Наивысшая скорость многоэтапных превращений при наименьших количествах промежуточных продуктов достигается при переходе к полиферментным комплексам с «правильным», соответствующим маршруту превращений, расположением ферментов в комплексе и наиболее интенсивным противоградиент- ным переносом исходного субстрата внутрь органеллы и конечного продукта из нее. Суммарная скорость потока продукта при таком многоэтапном превращении лимитируется лишь собственно каталитической активностью наиболее медленного фермента в полиферментном комплексе и скоростью отвода (быть может, той же диффузии или активной откачки) образующегося конечного продукта.
До появления аппаратов перемещения в пространстве, обеспечивающих радикальное преодоление диффузионных ограничений, давление естественного отбора должно было обусловить йаиболее быструю в данной системе дуффузию.
Итак, биологический смысл структурной организации многоэтапных биохимических процессов, биологический смысл образования митохондрий и хлоропластов — предельно возможное ускорение превращений веществ и наивысшая скорость регуляции этих превращений.
Второе, возможно столь же важное, биологическое преимущество структурно организованных систем по сравнению с гомогенными растворами — резкая экономия каталитического вещества. Действительно, относительно небольшое число митохондрий обслуживает очень большие объемы протоплазмы. Если бы удалось так гомогенизировать митохондрии, чтобы все содержащиеся в них ферменты в неповрежденном состоянии перешли в раствор, то итоговая скорость превращения веществ (даже при избыточной их концентрации) была бы очень низкой. Для обслуживания такой гомогенной системы потребовалось бы значительно больше митохондриального вещества (то же касается вещества хлоропластов и т. п.).
Более или менее правильное пространственное расположение дискретных порций малого количества ферментов позволяет обеспечить высокую скорость превращения веществ, достигаемую в гомогенном растворе лишь при относительно очень высоких концентрациях ферментов.
Биологические преимущества более или менее правильной пространственной организации надмолекулярных полифермент- ных комплексов основаны на «игре на градиенте». Расположение небольшого числа митохондрий на некотором расстоянии друг от друга позволяет им использовать зависимость скорости диффузии от величины градиента концентрации диффундирующих веществ (см. рис. 10).
Представим себе, что поток вещества из внешней среды в клетку перерабатывается в конечном итоге в какой-либо полезный продукт.
Пусть для конкретности речь идет о диффузии кислорода и поглощении его митохондриями, вырабатывающими АТФ. В принципе можно рассчитать, сколько митохондрий необходимо для оптимальной работы и как они должны быть расположены (на каких расстояниях друг от друга, от поверхности клетки, от потребителей АТФ, например, миофибрилл (рис. 10). Для дедуктивного построения было бы желательно получить аналитическое выражение, связывающее коэффициент диффузии, каталитическую активность митохондрий, расстояние от границы клетки и т. п. с необходимым числом митохондрий и расстояниями между ними. Задача эта, однако, очень сложна. Можно не сомневаться, что именно такие оптимальные решения были найдены во всех конкретных случаях под давлением естественного отбора.
Теперь вполне уместно сказать, что изложенные выше соображения о биологическом смысле замены сплошной каталитической поверхности дискретными пространственно отдаленными друг от друга структурами (клеточными органеллами) являются результатом применения -системы аналогичных представлений, давно развитых в другой области.
Много лет назад (в 1947 г.) на меня произвело очень сильное впечатление изложение вопроса об устьичной регулировке транспирации листьев выдающимся лектором, профессором био-
РИС. 10. Переработка малым числом дискретных каталитиче- ски-активных структур потоков субстрата с большой поверхности за счет краевых эффектов диффузии

So — концентрация субстрата у наружной поверхности; Р — концентрация продукта
логического факультета МГУ Д. А. Сабининым. Несколько менее красочно материал этой лекции опубликован в книге [251]. Слова Сабинина имеют прямое отношение к рассматриваемому нами вопросу, и я приведу их полностью: «...Еще в 1800 г. Брауном и Эскомбом [370] было проведено изучение диффузии углекислоты через целлюлозные пластинки с отверстиями диаметром 0,38 мм. Это исследование показало, что при общей площади пор указанного диаметра, составляющей лишь 11% от площади пластины, через нее проходило столько же углекислоты, как и через свободное сечение трубки, иными словами, перегородка з данном случае переставала быть препятствием, снижавшим скорость диффузии. Браун и Эскомб предположили, что при порах еще меньшего диаметра, равного ширине устичных щелей эпидермиса, равенство скорости диффузионного тока при наличии и в отсутствии перегородки с порами будет достигнуто, если общая площадь пор составит лишь 1—3% площади пластинки. Такое соотношение площади устьичных щелей и всей поверхности эпидермиса является обычным у большинства растений.
Предположение Брауна и Эскомба примерно через четверть века могло быть подвергнуто экспериментальной проверке, когда фирмой Цейсс были изготовлены тонкие медные пластинки с порами диаметром около 10 мкм. Эта величина пор примерно равна диаметру открытых устьичных щелей. На 1 ммг площади медного листка толщиной 20 мкм наносилось до 100 пор. Для невооруженного глаза такая пластинка, пронизанная микроскопически мелкими порами, представлялась совершенно сплошной. Но при изучении испарения воды с поверхности небольших сосудиков, накрытых этими пластинками или оставленных совершенно непокрытыми, оказывалось, что в обоих случаях скорость испарения воды была примерно одинаковой. Этот удивительный результат явился блестящим подтверждением теоретических представлений о значении градиента дефицита насыщения как фактора скорости испарения.
При диффузии водяного пара через отверстия, равные по диаметру ширине открытых устьичных щелей, краевое испарение играет столь большую роль, что скорость испарения с единицы поверхности поры почти в 80 раз превышает соответствующуювеличину для свободной поверхности. В работах Губера [403] имеется обширный экспериментальный материал, позволяющий установить зависимость испарения от диаметра пор и величины общего сечения всех пор по отношению ко всей площади пластинки» [251, с. 132].
Было бы несправедливо не сказать, что за 20 лет до Брауна и Эскомба Стефан [461] теоретически и экспериментально исследовал испарение и диффузию жидкостей из ограниченных поверхностей и отверстий. При этом он исходил из работ Гельмгольца по прохождению звуковой волны через трубы и отверстия (органные трубы) и своих собственных аналогий между концентрационными (диффузионными) и электростатическими градиентами.
Стефан показал, в частности, что в силу краевых эффектов скорость диффузии через круглое отверстие пропорциональна его радиусу, а не площади, т. е.
v=4 DRp,
где v — скорость диффузии, D — коэффициент диффузии, р — величина градиента концентрации, R — радиус отверстия.
Таким образом, при уменьшении площади круглого отверстия в п раз скорость диффузии через него убывает лишь в У/г раз. Этим и объясняются устьичные эффекты. Их количественный анализ с учетом формы и взаимного влияния соседних отверстий сложен и требует применения ЭВМ.
Из изложенного выше ясно, что роль пространственных факторов, «игра на градиенте» давно осознана в физиологии растений ‘. Тем удивительнее, что в весьма близкой к физиологии растений области — биохимии — кинетическая роль структуры до сих пор осталась неосознанной. Насколько мне известно, до сих пор аналогия между пластинкой с малым числом дырок и, например, клеткой с митохондриями не проводилась. Отчасти это объясняется тем особым значением, которое придавалось каждому успешному осуществлению того или иного биохимического процесса в бесструктурной системе, в гомогенном растворе. В самом деле, лишь реализация ферментативного процесса в бесструктурной системе много десятилетий считалась обязательным условием изучения механизма такого процесса. Это обстоятельство было очень четко подчеркнуто В. А. Энгельгардтом в 1962 г. Он писал:
«В заключение подчеркнем, что все сказанное не означает, разумеется, недооценки значения структуры. Но мы предпочли придерживаться области тех вопросов, которые могут быть выражены химическими уравнениями и конкретными физическими представлениями.
К сожалению, вопросы структуры опутаны весьма больши' ореолом неопределенности, в них можно вкладывать все, что угодно, от структуры молекулы до структуры любого вещества, существа и даже до структуры популяций.
Повторяем, структура важна, ею пренебрегать нельзя. Но опыт биохимии дает нам примеры иного порядка. Был длительный период, когда все, что не удавалось описать химическими уравнениями, относилось за счет структуры. В период, когда ученые не могли ничего сказать о механизме дыхательного процесса, Варбург с восторгом говорил о роли структуры для клеточного дыхания. Но что осталось от роли структуры для клеточного дыхания? (его интенсивность (!)—С.Ш.). В настоящее время можно весь процесс клеточного дыхания воспроизвести в гомогенном растворе структуры. От структуры, которая фигурировала как основное звено Варбурга, не осталось и следа. Опыт учит, что термин «структура» обычно бывает ширмой, за которой скрывается наше незнание... Если мы можем воспроизвести кардинальные проявления жизнедеятельности в бесструктурной системе, нам нечего желать лучшего. Мы можем в бесструктурной системе воспроизвести редупликацию молекул ДНК, мы можем в бесструктурной системе воспроизвести такой коренной биологический процесс, как явление гибридизации». [352, с. 47].
Энгельгардт совершенно прав, говоря о возможности в принципе воспроизвести даже самые сложные биохимические процессы в бесструктурных системах. Строгая, высоко совершенная стереоспецифичность молекул белков-ферментов, связывающих в активном центре лишь молекулы своего субстрата, их способность различать например, окисленное и восстановленное состояние молекул субстратов (мы знаем это на примере НАД- дегидрогеназ)вполне может обеспечить нужную последовательность реагирования в многоэтапных процессах [471]. Для правильного, безошибочного осуществления многоступенчатых процессов превращения веществ, освобождения и усвоения энер- тии структура не нужна. Но смысл структуры не в этом. Структура обеспечивает большее биохимическое совершенство — в структурированных системах процессы идут быстрее, чем в бесструктурных. Структура нужна для ускорения многоэтапных превращений и их более совершенной регуляции-, она увеличивает экономичность системы, т. е. обеспечивает переработку наи- -большей массы субстратов наименее «материалоемкими» системами катализаторов-ферментов[8].
Глава 6
МЕМБРАНЫ, ИОННАЯ АСИММЕТРИЯ
Живое вещество должно быть дискретным, разделенным на организмы.
Возникновение дискретных организмов сопряжено с решением в ходе эволюции многих проблем.
Необходимость сохранения целости организма приводит к возникновению сигнальной системы, реагирующей на повреждение, а затем и на возможное повреждение (раздражение) клетки.
Асимметричное распределение ионов в системе клетка — среда является эволюционным результатом и способом осуществления возникновения раздражимости и возбудимости клеток.
Причина повышенной концентрации внутри клетки ионов калия, а снаружи ионов натрия заключена в геологической истории Земли.
В предыдущей главе мы пришли к необходимости эволюционного возникновения структурной организации для преодоления диффузионных ограничений биохимических процессов.
Структурная организация оказывается необходимой и по другим не менее важным эволюционным соображениям.
Ранее (гл. 2) мы подчеркивали, что «биологическое (кинетическое) совершенство» зависит от скорости увеличения массы вещества данного вида. Казалось бы, если не принимать в расчет диффузионные ограничения, вещества данного вида — матричные молекулы и соответствующие ферменты могли бы существовать в виде неразделенной смеси — «живого киселя», по выражению Н. В. Тимофеева-Ресовского.
Однако в этом случае естественный отбор становится неэффективным. Условие эволюции, способ естественного отбора— гибель, смерть (менее приспособленных. Гибель должна быть строго избирательной — лишь менее совершенных. Это возможно лишь при условии существования «живого вещества» в воде дискретных порций, содержащих необходимый я достаточный, комплект матричных макромолекул, ферментов и всего прочего. Чрезмерная избыточность понижает селективные преимущества.
Оптимальный по размеру комплект матричных и каталитических полимеров я низкомолекулярных веществ необходимый и достаточный для жизни — это элементарный организм, клетка. Такой организм должен обладать специфической поверхностью, отделяющей его от внешней среды и препятствующей вымыванию веществ, нарушению необходимого их комплекта.
Следовательно, биологическая целесообразность, т. е. предельно совершенная форма гибели, смерти достигается при разделении «вещества» данного вида на в некотором смысле независимо погибающие организмы. Размеры этих первичных организмов— клеток должны удовлетворять двум противоположным требованиям: с одной стороны, они должны быть достаточно большими, чтобы в них мог поместиться весь необходимый для автономного существования набор матриц, ферментов и прочих молекул, а с другой,— достаточно малыми для достижения наиболее интенсивного процесса естественного отбора наиболее экономным образом, а также для обеспечения требуемой поверхности взаимодействия с внешней средой.
Теперь на очередь становится задача рассмотрения биохимических механизмов отграничения от внешней среды, специфической биохимии клеточных поверхностей, клеточной оболочки.
Наиболее естественным механизмом построения оболочек клеток, образования дискретных порций живого вещества в водной среде, является создание гидрофобных границ раздела. Поскольку речь идет об отграничении гидрофильных биохимических систем (ферменты, матричные молекулы и другие основные биохимические компоненты по необходимости гидрофильны) от окружающей водной среды, первичная гидрофобная граница могла образовываться лишь детергентами, т. е. веществами-пиб- ридами, содержащими гидрофильные группы, обращенные внутрь клетки, и гидрофобные группы, обращенные наружу. Таким образом, появление дискретных форм жизни — особей, клеток сопряжено с возникновением системы синтеза биодетергентов (например, фосфолипидов). Замечательным свойством детергентов является их способность образовывать дискретные структуры (пленки, мицеллы, коацерватные капли, пузыри, пену [94, 261, 422]. Эта способность обусловлена взаимодействием сравнительно небольших молекул детергентов друг с другом и с молекулами среды посредством ван-дер-ваальсовых и электростатических сил. Эволюционная необходимость детергентов и липидов, возможная роль коацерватных структур, появляющихся в смесях детергентов и белков в процессе возникновения жизни, рассмотрены А. И. Опариным и сотрудниками [94, 261].
Ключевая роль детергентов в формировании дискретных особей (в том числе и прекрасных) вполне соответствует представлению о возникновении Венеры (Афродиты) из пены морской.
Возникновение клетки представляется следующим образом. В ходе эволюции появляются ферментные системы синтеза детергентов (например, фосфолипидов). Слой таких детергентов обволакивает комплект матричных молекул, молекул ферментов и низкомолекулярных метаболитов, образуя поверхностную мембрану. Синтез компонент мембраны строго регулируется — их количество должно соответствовать поверхности клетки.
Тут возникает непростая проблема: детергенты должны быть ориентированы, по-видимому, своими полярными группами внутрь клетки, а неполярными — наружу. Как достигается различение внутреннего и наружного;' Как предотвратить «нечаянное» выворачивание мембраны наизнанку? Это, по-видимому, возможно лишь посредством более или менее специфического взаимодействия с содержимым клетки, которое само не должно вытекать из нее и в отсутствии мембраны. Иными словами, должен существовать твердый каркас, на котором закрепляется мембрана. Этот каркас (наиболее вероятно — белковый гель) представляет собой трехмерную структуру, состоящую из фибриллярных макромолекул белка типа коллагена, тубулина или актина.
С другой, наружной, стороны не вся поверхность мембраны клетки должна быть гидрофобной — в противном случае произошло бы слипание клеток и стало бы невозможным необходимое для жизнедеятельности взаимодействие со средой. Следовательно, поверхностная мембрана должна обладать сложной структурой — ее обращенная внутрь сторона должна содержать химические группы, специфически соединяющиеся с внутренним каркасом, с внутриклеточными белками, а ее наружная сторона не должна быть однородно гидрофобной. Каркас, на который натягивается, к которому прикрепляется поверхностная мембрана, необходим, как мне кажется, и для того, чтобы эта мембрана не отделялась от клетки. В самом деле, детергентный слой вполне может образовать автономную структуру, и не обволакивающую клетку. Поэтому нужно, чтобы взаимодействие мембранных детергентов с каркасом было бы более сильным и специфичным, чем взаимодействие одной молекулы детергента с другой. Ясно, что такое обволакивание мембраной внутриклеточного содержимого нельзя обеспечить лишь за счет неспецифических электростатических или гидрофобных взаимодействий.
Много лет назад Н. К- Кольцов обнаружил, что форма клеток определяется формой каркаса, состоящего, как он считал, из коллагеновых волокон [144]. В последние годы весьма интенсивно развиваются исследования фибриллярных структур, определяющих форму клеток. Эти структуры состоят из так называемых микрофиламентов. Основная компонента микрофила- ментов — актин, т. е. один из двух главных белков мышц [392, 393]. Микрофиламенты расположены непосредственно под мембраной клетки. Однако, насколько мне известно, вопрос об их взаимодействии с мембраной не изучен. Неясно также, следует ли исключить предположение о коллагеновом каркасе. Возможность участия коллагена в таком каркасе следует из опытов по созданию на нем искусственных возбудимых липопротеидных мембран [369].
Итак, по указанным выше причинам предельно совершенные биохимические системы оказываются представленными в виде дискретных порций живого вещества. И эта дискретность осуществляется посредством биосинтеза специализированных де- тергентных молекул, по-видимому, спонтанно образующих граничные и прочие мембраны клеток. Мы видим, что и в этом случае все необходимое разнообразие свойств поверхностей клеток можно получить «наборным» методом — посредством должного сочетания относительно небольшого числа универсальных компонент — букв (фосфолипидных и прочих), составляющих слова «мембранного языка». Однако в отличие от полипеп- тидного и полинуклеотидного языков в мембранном языке не нужен строго детерминированный порядок слов и даже букв в слове, а достаточно, чтобы в данной структуре слова и буквы были представлены в должных соотношениях. Поэтому при наличии специфического белкового каркаса нет необходимости в соединении отдельных молекул в мембранных структурах валентными или строго определенными водородными связями — вполне удовлетворительные результаты достигаются и при соединении молекул ван-дер-ваальсовыми и электростатическими связями. Действительно замечательным свойством системы липиды— детергенты — вода является способность к автоматической самосборке мембран [307а, 422]. Мембраны в этом смысле вполне детерминированы физическими и химическими свойствами компонент самой системы. Особенности строения возникающей мембраны зависят почти исключительно от того, какие липиды, детергенты, электролиты и неэлектролиты содержатся в системе и в каком количестве. Следовательно, свойства мембраны определяются характером и интенсивностью биосинтетических процессов в протоплазме. Так, число и качество дефектов в мембране, мест, где плотная упаковка молекул (например, лецитина) нарушена, зависит от концентрации в системе других детергентных молекул. В конечном итоге, свойства мембраны записаны в соответствующих матричных текстах, определяю- ющих синтез в нужном месте и в нужное время необходимых ферментов, и задаются оперативным изменением активности уже синтезированных ферментов.
Попробуем представить себе возможные механизмы функционирования поверхностных клеточных мембран. Их функция состоит в избирательном пропускании одних веществ и не пропускании других, причем должно осуществляться как движение веществ по термодинамическому градиенту (из места с более высокой концентрацией в места с меньшей концентрацией), так и противоградиентное движение.
Ясно, что противоградиентное движение веществ может осуществляться лишь при использовании свободной энергии в каком-либо сопряженном экзэргоническом процессе.
Один из мыслимых способов противоградиентного перемещения состоит в уничтожении нормального градиента концентрации переносимого вещества. Это может осуществляться посредством связывания поступившего вещества в кинетически (осмотически) неактивный комплекс или же его вовлечения в какой-либо химический процесс. Так, глюкоза может исчезатьв процессе образования из нее гликогена, аминокислоты — в процессах синтеза белка и т. д.
В этих случаях не возникает принципиальных трудностей в представлении исходных шагов эволюционного совершенствования. Задача такой мембраны — обеспечение лишь грубой избирательности, наличие в ней дырок, пор, позволяющих нужным веществам двигаться (диффундировать) по «искусственному» градиенту и препятствующих вытеканию протоплазмы. Я подчеркиваю слово вытекание, так как именно в препятствии макроскопическим потокам жидкости и состоит первоначальная функция мембран. Множество микродырок не мешает макроскопической функции мембраны. Так, пористые, ячеистые например, вязанные свитеры и даже крупноячеистые вуали, ограничивая макроскопические потоки воздуха, сохраняют прикожный нагретый слой воздуха, и поэтому «греют» людей.
Регулирование свойств такой мембраны также может быть очень грубым — нет нужды в специальной подгонке диаметра пор к размеру проходящих через мембрану молекул.
Однако задача усложняется тем, что в биохимических процессах с необходимостью должны участвовать и низкомолекулярные промежуточные продукты, не все время пребывающие в комплексах с макромолекулами.
Таким образом, сведение избирательности переноса веществ к биохимической специфичности — к .избирательному вовлечению веществ в соответствующие биохимические процессы, лишь первый шаг эволюционного совершенствования трансмембранной системы переноса веществ.
Какие пути увеличения избирательности проницаемости мембран могут существовать в процессе эволюции?
Конечно, можно представить себе такое состояние протоплазмы, при котором все низкомолекулярные компоненты биохимических процессов оказываются в специфических комплексах с макромолекулами. Для его реализации нужно, чтобы образование этих комплексов было термодинамически выгодным. Величина выигрыша энергии при образовании комплекса низкомолекулярного вещества с макромолекулой и определит возможность «противоградиентного» переноса данного вещества. Слово «противоградиентный» взято в кавычки, поскольку в этом случае никакой противоградиентности нет — связывание вещества уменьшает его осмотическую, кинетическую активность. В месте образования комплекса создается активность (концентрация), меньшая, чем в других местах. Остается позаботиться
о том, чтобы не вытекали сами макромолекулярные комплексы, а это обеспечивается образованием наружной дырявой, сетчатой мембраны или же соединением всех или большей части макромолекул в трехмерную сеть — гель. Такие желеобразные дискретные организмы в принципе удовлетворяют условиям дискретности и стабильности оптимальных по размерам комплектов матричных молекул, ферментов и метаболитов.
Однако скорости биохимических процессов в таких организмах будут далеко не предельно возможными. Скорости транспорта субстратов и продуктов в биохимических системах, где все участники связаны с макромолекулами геля, очень малы. Условием быстрого протекания биохимических процессов является свободное, неограниченное соединением с макромолекулами движение низкомолекулярных веществ в протоплазме. Эволюция должна обеспечить максимум скорости узловых биохимических процессов без специальных уже нехимических приспособлений и это достигается при совершенно незатрудненной диффузии.
Следовательно, среда, в которой протекают наиболее важные биохимические процессы, должна быть жидкой *. В этой маловязкой бесструктурной жидкой среде могут находиться относительно небольшие компактные структурные образования — митохондрии, хлоропласты и пр., но основная среда должна быть жидкой и основные метаболиты в ней не должны быть связаны с гелеобразным комплексом. Большинство низкомолекулярных метаболитов (субстратов, ионов металлов, анионов, кофермен- тов и пр.) в соответствии с требованием максимально возможной скорости протекания узловых биохимических процессов должно свободно диффундировать в маловязкой жидкой протоплазме. Следовательно, крупно-ячеистая мембрана служит недостаточно совершенной границей раздела организма и внешней среды, так как она не будет препятствовать утечке метаболитов. Мембрана должна обладать избирательной проницаемостью.
Приведенное рассуждение, т. е. вывод о невозможности обеспечения должной интенсивности биохимических процессов в клетке при осуществлении специфического переноса веществ посредством специфического связывания в толще протоплазмы, имеет прямое отношение к недавней острой дискуссии. Д. Н. Насонов и его последователи [220, 221] полагали, что именно специфическое связывание в протоплазме лежит в основе механизма избирательного противоградиентного переноса. Д. Л. Рубинштейн [246, 247] наиболее ярко представлял противоположную точку зрения, а именно необходимость существования избирательно проницаемых клеточных мембран. За прошедшие после дискуссии годы наши знания о механизмах специфического переноса веществ в биологических системах чрезвычайно расширились и углубились. Ключевая роль мембран давно уже не вызывает сомнений. Однако механизм специфического связывания отнюдь нельзя считать полностью отвергнутым. Из общих соображений, приведенных выше, следует лишь, что внутриклеточное связывание низкомолекулярных веществ как способ проти- воградиентного транспорта не совместимо с высокой скоростью биохимических процессов. Однако в ряде специальных случаев внутриклеточное связывание может оказаться очень эффективным.
Мы видим, следовательно, что в ходе эволюции возникли несовместимые требования—с одной стороны, необходимо жидкое состояние значительной части внутриклеточного содержимого, несвязанное состояние внутриклеточных низкомолекулярных веществ (пока они не включаются непосредственно в ферментативные процессы), свободная их диффузия, а с другой, необходим строго специфичный противоградиентный перенос через клеточную мембрану. Выход из этого положения может состоять в постулировании аппарата специфического связывания и переноса веществ, сосредоточенного в мембране или примембранных слоях, и в сопряжении процессов переноса с расходом макро- эргических соединений (или в более общей формулировке — сопряжении эндэргонического переноса с каким-либо экзэргониче- ским процессом).
Истинные механизмы специфического трансмембранного транспорта все еще не вполне выяснены. Этим вопросам посвящен ряд фундаментальных исследований (см. [5, 33, 48, 62, 158, 171, 172, 177, 178, 180, 197, 225, 226, 285, 314, 365, 383, 434, 435, 451, 459]), к которым и должен обратиться читатель. Тем не менее, в общем виде ответ известен. Противоградиентный перенос осуществляется или за счет свободной энергии гидролиза пирофосфатной связи АТФ или за счет распада ее макроэрги- ческого предшественника. Специфичность транспорта веществ через мембрану достигается либо путем связывания этих веществ со специфическими молекулами белков-переносчиков, либо посредством калиброванных пор [399].
Целостность поверхностной мембраны — условие существования клетки. Раздражимость и возбудимость. Ясно, что повреждение поверхностной оболочки (клеточной мембраны) должно приводить к гибели клетки. Нарушение целостности мембраны сопряжено с потерей внутриклеточного содержимого и вхождением в клетку вредных компонент из внешней среды. Условием дальнейшего эволюционного прогресса оказывается теперь выработка быстрой реакции на повреждение. Эта реакция должна быть сопряжена с включением (и выключением) определенных биосинтетических систем, например, с включением системы интенсивного синтеза фосфолипидов и белков. Таким образом, очередная задача эволюции состоит в выборе предельно совершенного вида сигналов о целостности и нарушении целостности клеток. Кажется разумным предположить, что эти сигнальные функции могла бы выполнять относительно безвредная компонента внешней среды, всегда присутствующая в достаточных количествах в первичной среде обитания древних организмов. Для выбора такой компоненты необходимо выяснить состав первичной среды обитания живых организмов.
При таком анализе становятся ясными следующие выводы. Жизнь возникла в море. Химический состав морской воды определен составом земной коры, физическими и химическими свойствами ее компонент, а следовательно, химическим составом планеты. Высокая вероятность возникновения жизни именно в море подчеркивалась почти всеми. Мне также кажется это почти бесспорным. Море замечательно своей стабильностью. Химический состав морской воды определился условиями ее возникновения я существования. Первичная вулканическая вода — «рассол»—содержала все растворимые компоненты коры в соотношениях, определяемых как концентрацией этих компонент в коре, так и их относительной растворимостью. Однако по мере фильтрации извергаемой вулканами воды через образующиеся и уже образовавшиеся осадочные породы — глины (алюмосиликаты с разным содержанием железа и алюминия) из первичного рассола избирательно извлекался калий (по причинам, хорошо изложенным Г. Г. Маленковым), а в воде, стекающей в море, оказывалась повышенной концентрация натрия. Избирательное сродство сэндвичевых алюмосиликатов (глин) к калию настолько велико, что уже в первых слоях глины, соприкасавшихся с первичным рассолом задерживалось основное количество калия, и состав профильтрованного раствора приближался к составу морской воды. Свойство глин избирательно накапливать калий делает их в этом отношении похожими на «живое 1вещество» (и оправдывает распространенную ранее гипотезу об изготовлении первого человека из глины...).
Ниже приводятся округленные данные о составе морской воды (табл. 7).
Солевой состав плазмы крови различных животных, характерный для земных организмов, приведен в табл. 8, 9 [68].
Из приведенных данных видно сходство (а иногда даже совпадение) соотношения концентраций ионов в плазме крови животных и морской воде. Однако по абсолютным величинам содержание солей в морской воде отличается от солености плазмы крови — в морской воде почти в 3 раза больше натрия, в несколько раз меньше калия и почти в 12 раз больше магния. Сторонники идеи о том, 4i(j в наших жилах течет, в сущности, мор-таблица 7
КОНЦЕНТРАЦИЯ ОСНОВНЫХ КОМПОНЕНТОВ МОРСКОЙ ВОДЫ *
Вещество
%
м
Вещество
%
м
Натрий
10,56
0,44
Стронций
0,01
1,3-10-4
Калий
0,38
0,0095
Хлор
18,98
0,535
Магний
1,27
0,047
Сульфаты
2,65
0,028
Кальций
0,40
0,0096
Карбонаты
0,14
0,0023
* Данные заимствованы из 2-го издания БСЭ.
ТАБЛИЦА 8
СООТНОШЕНИЕ КОНЦЕНТРАЦИЙ ИОНОВ В ПЛАЗМЕ КРОВИ ПОЗВОНОЧНЫХ И В МОРСКОЙ ВОДЕ (%) [68]
Позвоночные
Калий
Кальций
Магний
Млекопитающие
3,6
1.9
0,8
Птицы
3,2
—
Пресмыкающиеся
4,1
—
Амфибии
5,8
1,9
1,1
Рыбы
5,3
Морская вода
3,6
3,9
12,1
Примечание. Концентрация натрия принята равной 100%.
ТАБЛИЦА 9
КОНЦЕНТРАЦИИ ИОНОВ В ПЛАЗМЕ КРОВИ (ММОЛ/Л) [68]
Позвоночные
Натрий
Калий
Кальций
Магний
Человек
145
5,12
2,49
1,23
Курица
141
4,2
Черепаха
122
5,1
Лягушка
102
5,9
2.0
1,2
Окунь
88
5,1
ская вода, не смущаются различием в абсолютных величинах содержания катионов. На протяжении сотен миллионов лет геологической эволюции солевой состав моря изменялся: осадочные породы поглощали все больше калия, при эрозии коры выщелачивались все большие количества натрия и т. д. Таким образом, если учитывать все эти изменения, то кровь (вернее, плазма крови) современных животных аналогична по своему солевому составу воде древних морей. Выйдя в давние времена из древ
них морей, животные унесли в своих кровеносных сосудах привычную и необходимую для существования их клеток среду, ставшую теперь внутренней средой организма.
Поддержание необходимого состава этой внутренней среды — одно из главных направлений физиологической работы многоклеточных организмов — их прото- или мезо- или метанефридии (почки) расходуют огромное количество энергии, сопоставимое с количеством энергии, затрачиваемым сердцем и скелетными мышцами (см. [436]).
Эти траты необходимы для обеспечения строго определенных условий существования клеток. Солевой состав, концентрация катионов и анионов в клетке и их соотношение совсем иные, чем во внешней по отношению к клеткам среде — в морской воде и в жидкости межклеточных пространств (табл. 10).
ТАБЛИЦА 10
КОНЦЕНТРАЦИЯ Ионов В ТКАНЯХ И ЖИДКОСТЯХ ТЕЛА (в мМ) [68]
Животное
Ткань или жидкость тела
Натрий
Калий
Кальций
Магний
Хлор
Крыса
Плазма крови
145
6,2
3,1
1,6
116
Мышца
27
101
1,5
11,0
16
Лягушка
Плазма крови
104
2,5
2,0
1,2
74
Мышца
24
85
2,5
11,3
10
Беззубка
Кровь
15,4
0,38
5,3
0,35
10,5
Мышца
5,2
10,5
5,4
2,46
10,6
Осминог
Кровь
525
12,2
11,6
57,2
480
Мышца
81
101
3,7
12,7
93
Голотурия
Жидкость тела
460
11,8
10,7
50
523
Мышца
191
139
89
39
277
Не имея сейчас возможности отвлекаться на рассмотрение своеобразия солевого состава клеток и внутренней среды разных животных, подчеркнем лишь одно. У большинства живых существ на Земле все полноценные живые клетки содержат натрий в концентрациях, значительно меньших, чем в морской воде или во внутренней среде организмов. Наоборот — концентрация калия в клетках во много раз выше, чем во внеклеточной среде.
Итак, мы забежали вперед — вместо дедуктивного построения предельно совершенного дискретного организма, клетки, мы сразу заглянули в ответ — посмотрели, что же получилось на Земле. Мы и дальше будем так иногда поступать, поскольку вполне «честное» решение эволюционных задач в ряде случаев еще невозможно. Зато мы можем с удовлетворением считать, что нам удалось выяснить, решению какой эволюционной задачи соответствует известный нам ответ. Итак, ответ гласит: калия в клетке больше, а натрия меньше, чем в среде, ее окружающей. Мы знаем (зернее думаем, что знаем) в чем здесь дело, каков эволюционный смысл такого соотношения концентраций.
Поддержание градиента концентрации такого химически вполне безобидного иона, как ион натрия (или калия), позволяет клетке быть в постоянной готовности к реакции на случай повреждения (а в дальнейшем, при совершенствовании этого механизма,—уже л,ишь при угрозе повреждения). Изменение градиента концентрации служит сигналом для принятия защитных мер, прежде всего для включения механизмов синтеза пограничного вещества — липидов и белков клеточной мембраны. Необходима тонкая регулировка процесса биосинтеза элементов мембраны (как и других биосинтетических процессов — синтезов ферментов и матриц). В самом деле, избыточный синтез делает жизнь невозможной — клетка переполняется веществом, на избыточный синтез напрасно расходуется вещество и энергия. Таким образок, возникает необходимость непрерывного (пока существует опасность повреждения) поддержания готовности к реагированию, к раздражению и возбуждению. Для этого приходится все время поддерживать искусственный градиент концентрации химически безвредных сигнальных веществ — ионов натрия и калия. Постоянная боевая готовность клетки обычно сопряжена с непрерывным расходом энергии. В неблагоприятных солевых условиях расходы на поддержание необходимого градиента могут превысить возможности клетки. Тогда становится «дешевле похоронить, чем прокормить».... На этом основано использование в качестве консервирующего средства поваренной соли —в сильно соленой среде истощаются и гибнут нежелательные нам микроорганизмы. Поэтому безжизненны резко соленые водоемы и столь мало пригодны для жизни солончаки (см., однако, стр. 141).
Необходимость непрерывного поддержания асимметрии распределения нонов натрия и калия в системе клетка — внешняя среда вовсе не означает, что этот механизм должен все время функционировать.
Было бы нерационально, бессмысленно непрерывно откачивать воду из дырявой лодки. Во всяком случае, ни о каком совершенстве тут говорить нечего. Необходимый градиент можно сохранить, закрыв пробоины. Конечно, умеют это делать далеко не все органязмы, а лишь специально приспособленные к жизни в необычных условиях чрезмерной солености или пресности. Физиология и биология приспособлений к жизни водных организмов при разной степени солености рассмотрена [311].
Итак, первичный механизм реагирования клеток на повреждение (раздражение) был, по-видимому, основан на изменении внутриклеточной концентрации ионов калия и натрия.
Мерой градиента концентрации ионов является электрохи
мический потенциал. Для реагирования на внешние воздействия вовсе не обязательно доводить дело до грубого повреждения клетки — функции сигнала может взять на себя изменение электрохимического потенциала. Ясно, что обязательным компонентом такого механизма должен быть процесс резкого активирования биосинтеза биодетергентов и белков. Следовательно, процессы считывания текста нуклеотидных матриц и синтезы необходимых ферментов, а также, быть может, активность уже готовых молекул ферментов должны быть чувствительны к уровню концентрации одновалентных катионов.
Так возникает в процессе эволюции электрофизиологический механизм, детальное рассмотрение которого выходит за пределы задач этой главы.
Нам остается, однако, обсудить здесь еще один важный вопрос: почему в процессе эволюции на Земле возникла именно такая ионная асимметрия.
Мы 'Считали, что ионная асимметрия, т. е. «противоестественный» градиент концентраций возник как эволюционное следствие необходимости клетки реагировать на повреждение. Однако мы уже видели, что в «простых» глинах происходит резко избирательное накопление калия, и, следовательно, возникает ионная асимметрия, а именно, натрий остается вне глины, калий проникает внутрь. Строго говоря, в глине никакого противогра- диентного накопления калия нет. Напротив, строго по предписаниям термодинамики калий заполняет пустоты в структуре сэндвичевых глин, в результате чего свободная энергия системы понижается [192]. В глиняной модели живого организма можно предусмотреть и реагирование на повреждение. В самом деле, всякое нарушение целостности сопряжено с механическим воздействием. А глина, как и другие гелеобразующие материалы, тиксотропна, т. е. разжижается при механическом воздействии (вот почему в мокрой глине ноги вязнут тем сильнее, чем интенсивнее попытки выбраться). Нарушение структуры тиксотроп- ных гелей — вполне удобный в принципе механизм раздражения, который может включить необходимые для восстановления целостности клетки процессы биосинтеза.
Таким образом, даже изменение ионной асимметрии — выход калия ,и вход натрия в «глиняную клетку» — вполне можно считать следствием, а не условием раздражения и возбуждения клетки. Для этого необходимо только, чтобы при тиксотропном разрушении структуры глины изменялось бы ее сродство к калию. И в самом деле, почему бы не считать такой тиксотропный механизм, механизм перехода гель — золь вполне соответствующим эволюционным задачам? Почему бы не считать ионную асимметрию лишь следствием структурных особенностей прото- плазматических структур?
Выбор правильного решения здесь очень не прост. Ведь мы должны найти доводы, основанные на убедительных общих соображениях, а не только на сведениях о механизмах жизнедеятельности земных организмов.
Я уже отмечал, что еще очень недавно исследователи были разделены на два непримиримых лагеря — сторонников сорбционной теории, возглавляемых Д. Н. Насоновым, и сторонников мембранной теории во главе с Д. Л. Рубинштейном. Темпераментные, не всегда академические споры завершились во втором поколении противников победой мембранной теории. Она восторжествовала благодаря замечательным достижениям в изучении биоэлектрических явлений на гигантских аксонах кальмаров и других удобных объектах. В самом деле, можно перфу- зировать, промыть гигантский аксон, удалить из него всю протоплазму— а способность к раздражению, к генерации нервного импульса останется. Более того, можно получить искусственную- возбудимую мембрану, обладающую всеми основными свойствами естественной [434, 435]. Следовательно, функция аксона и. в самом деле обусловлена лишь его мембраной. Следовательно, ионная асимметрия и в самом деле создается без участия избирательной сорбции ионов в толще внутриклеточной протоплазмы.
Мы уже очень много знаем о природе избирательных ионных насосов, локализованных в клеточной мембране. Их функцию' выполняют ферменты, аналогичные асимметрично расположенной в мембране К, Na-эктивируемый АТФазе (см. [33, 180, 459]), в сочетании с избирательно проницаемыми каналами.
Мы все больше узнаем о свойствах и природе веществ, модифицирующих проницаемость био-мембран. Все это так. Однако еще раз подчеркиваю — нам нужно понять, в силу каких причин в процессе эволюции возникли именно такие механизмы? Один из возможных ответов гласит: данное приспособление возникло как неизбежное следствие физических и (или) химических свойств веществ, вовлеченных в процесс эволюции. В духе такого ответа можно сказать, что ионные градиенты, противотер- модинамлческая асимметрия ионов в системе клетка — внешняя среда возникли как неизбежное следствие образования биоде- тергентных мембран. Если в силу эволюционной необходимости в дискретности живого вещества появились мембраны, отграничивающие клетки от внешней среды, то вследствие физических и химических свойств этих мембран, их избирательной проницаемости возникла и ионная асимметрия. Ионная асимметрия оказалась весьма удачным показателем целостности клетки и в ходе дальнейшей эволюции смогла обеспечить реагирование клетки на внешние воздействия.
Очень возможно, что такой ход рассуждений верен. Однако тиксотропия, превращение геля в золь — важнейший физико-хи- мический процесс в протоплазме как современных, так и древних земных организмов. Мало того, этот процесс представляет неспецифический универсальный ответ клетки на различные воздействия. Я думаю, однако, что процесс превращения гель — золь не был выбран в ходе эволюции как универсальный ответ на нарушение целостности клетки по вполне простой причине — отнюдь не любые повреждения клетки сопровождаются механическими воздействиями, вполне вероятны и повреждения в результате химических воздействий. А «тихое» растворение мембраны клетки не менее опасно, чем ее разрыв в результате свирепого укуса.
И все же, почему натрий снаружи и калий внутри? В настоящее время мне кажется вполне убедительным ответ на этот вопрос, предложенный В. А. Твердисловым [285]. Натрий не потому изгоняется из клетки, что он изначально был менее желателен, чем калий. И калий входит в клетку не из-за своих особых химических или физико-химических достоинств. Дело обстояло, по-видимому, проще: при концентрациях этих катионов в древнем океане только их можно было использовать для сигнализации о целостности клетки. Однако для создания электрохимического потенциала, при котором концентрация «атрия внутри клетки была бы больше, чем в окружающей среде, необходимы очень высокие концентрации ионов натрия в клетке — порядка нескольких молей на 1 л, что весьма существенно изменило бы всю физико-химическую обстановку в клетке. Наоборот, калия в среде, в морской воде настолько мало, что даже при его вполне безобидных концентрациях внутри клетки порядка 0,1 моль!л, соотношение концентраций К внутри и снаружи оказывается достаточным для асимметричного распределения натрия и создания заметного электрохимического потенциала.
Из приведенного рассуждения следует, что при осуществлении эволюции в других условиях (если это возможно), при другом ионном составе среды вполне могут возникнуть организмы с другим набором сигнальных ионов внутри и вне клетки.
В земных организмах происходят биохимические процессы, в которых роли ионов натрия и калия противоположны. Так, существуют ферменты, активность которых увеличивается в присутствии ионов калия и угнетается в присутствии ионов натрия (см. табл. 6). Однако в соответствии с приведенным логическим построением специфическое, сигнальное значение должны иметь биохимические процессы синтеза компонент мембраны, включаемые поступающим в клетку натрием (при раздражении мембраны). Число известных нам ферментных систем,включая активируемых натрием, очень невелико (табл. 11).
В этой связи необходимо сказать, что, как следует из ряда работ [48, 62, 292, 451], влияние натрия на биохимические системы опосредовано ионами кальция. Ионы натрия, проникая внутрь клетки, вызывают изменение состояния ионов кальция — их переход из связанного в свободное состояние. Действительным исполнителем сигнальных функций в клетке, звеном, связывающим изменение состояния мембран с внутриклеточной
ТАБЛИЦА II
ВЛИЯНИЕ ИОНОВ № И К НА НЕКОТОРЫЕ БИОХИМИЧЕСКИЕ ПРОЦЕССЫ [62]
Фермент или ферментативный процесс
Иои-
актива-
тор
Иои-
ингиби-
тор
Фермент нли ферментативный процесс
Ион-
актива
тор
Иои-
ингиби-
тор
Синтез ацетилхолина
к+
Na+
РНК-полимераза
к+
Синтез белка на рибосомах
к+
Na+
ДНК-полимераза
к+
Na+
Синтез липидов
Na+
—
Фосфофруктокинззн
к+
—
Дыхание митохондрий
К+
Na+
биохимией, является кальций. Число ферментов (ферментных систем), активируемых ионами кальция очень велико.
Из табл. 11 следует, что все внутриклеточные ферментативные процессы, по-видимому, приспособились к нормальной ионной асимметрии, тогда как синтез липидов включается при увеличении концентрации Na+ — сигнала о возможном повреждении клеточной мембраны. К сожалению, надежных экспериментальных данных по этим вопросам очень мало [449],
Возможно и другое решение проблемы ремонта поврежденной мембраны в ответ на увеличение концентрации ионов натрия в клетке. Опыт показывает, что вытекающая из поврежденной клетки протоплазма немедленно покрывается вновь возникающей липопротеидной мембраной (см., например, [5, 462]). Это происходит так быстро, что говорить о включении биосинтеза, как источника материала для новой мембраны, нельзя. Некоторый аварийный запас фосфолипидов и белков для быстрой сборки мембраны в клетках имеется. (Тем не менее биосинтезы должны быть включены для восполнения этих запасов). Процесс образования поверхностной мембраны при повреждении клетки был предметом глубоких исследований и широких обобщений Л. Генльбруна [64, 396, 397]. Гейльбрун считал, что эта «поверхностная реакция преципитации» осуществляется при действии ионов кальция аналогично механизму свертывания крови. Новообразованию мембраны на вытекающей из клетки капле протоплазмы, в соответствии с этой гипотезой, происходит в результате превращения белка типа фибриногена в полимери- зующийся белок типа фибрина (и лишь затем по возникшему белковому каркасу выстраивается липопротеидная мембрана).
В заключение мне хочется обратить внимание на возможность непосредственного влияш-я ионов натрия на образование мембраны. Натриевые соли жирных кислот менее растворимы, чем калиевые: все знают, что калиевое мыло жидкое, а натриевое твердое. Вполне возможно поэтому, что проникновение в примембранный слой клетки ионов натрия стимулирует самосборку мембраны из уже имеющегося запаса липидов, фосфолипидов, жирных кислот — аналогично превращению жидкого калиевого мыла в твердое натриевое.
Глава 7
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ПРЕВРАЩЕНИЯ ЭНЕРГИИ
Создание системы энергетического обеспечения конвариантного матричного воспроизведения.
Свет — первичный источник энергии в биохимических системах. Накопление буферных рН-систем в фотохимическом преобразовании энергии. Макроэргические соединения.
Общие механизмы сопряженного фосфорилирования. Концепция Митчелла.
Гетеротрофы.
Рост кристаллов, образование полимерных молекул и аналогичные процессы завершаются при исчерпании термодинамического потенциала системы, например, тогда, когда раствор перестает быть насыщенным. Представим себе, что некоторые виды кристаллов (полимеров) могут возникать при меньших концентрациях предшественников, чем другие менее совершенные виды. Ясно, что такие более совершенные кристаллы, полимеры и победят в естественном отборе.
Кристаллизация и полимеризация, происходящие вопреки термодинамическому, концентрационному градиенту из истощенного раствора, в принципе осуществимы при сопряженном протекании экзэргонической реакции, в которой равновесие кристаллизации или полимеризации может сместиться в противогради- ентном направлении. То же справедливо и для синтезов.
Однако необходимость сверхравновесного синтеза — не единственная причина вовлечения под давлением естественного отбора источников энергии для конвариантного воспроизведения.
Совершенный процесс соединения мономеров в полимерную матричную цепь должен быть термодинамически невыгодным, требовать для своего осуществления подвода свободной энергии, быть эндэргоническим. Здесь под «должен быть» подразумевается «в результате эволюции» или «чтобы была возможной прогрессивная эволюция». В самом деле, если бы полимеризация была экзэргонической, термодинамически выгодной и полимерные цепи возникали бы -самопроизвольно, то сколько появилось бы ошибок в последовательности мономеров в полимерных нитях. Ведь нужно помнить, что любые последовательности мономеров в полимерных полинуклеотидной или полипептидной цепях термодинамически равновероятны[9]: необходимбарьер, препятствующий соединению мономеров. Полимеризация должна быть термодинамически невыгодной, должна осуществляться лишь при контролируемом подводе энергии
В ходе эволюции возникает еще одна энергетическая статья расходов, составляющая в дальнейшем значительную долю от общих потребностей. Чем совершеннее катализатор, тем он сложнее, тем вероятнее его разрушение в ходе функционирования, а также в результате термической денатурации. Становится необходимым восстановительный синтез, необходимы траты энергии для компенсации износа макромолекулярных механизмов.
Наконец, в процессе эволюции все время возрастают траты энергии на активное добывание пищи, активное создание необходимых внутренних и внешних условий существования.
Итак, энергия необходима для самого процесса полимеризации мономеров, для проведения этого процесса против концентрационного градиента, для компенсации износа и для обеспечения активной жизнедеятельности.
Посмотрим теперь, какие источники энергии могут быть использованы в ходе эволюции. Оценим сначала количество энергии, требуемой для обеспечения указанных выше нужд. Например, процесс синтеза пептидной связи относится к эндэргоничес- ким реакциям — для синтеза 1 моля пептидных связей необходимо затратить 3—4 ккал свободной энергии. По-видимому, когда- нибудь удастся из общих соображений оценить и желательную величину сверхравновесного синтеза. Допустим, что сверхравно- весная концентрация продукта в 105—106 раз превышает величину, определяемую термодинамическими соотношениями. В соответствии с известными термодинамическими формулами на это потребуется еще 7—8 ккал/моль:
С С
AF = - RT 1п ~ ж - 1400 lg кал,
где С, и С2 — концентрации исходных реагентов и конечных продуктов соответственно. Так, для обеспечения выбранной сверх- равновесной степени полимеризации аминокислот приходится тратить около 10 ккал на синтез 1 моля пептидных связей. Близкие по порядку величины мы получим при оценке энергетических расходов и в других процессах. Эти относительно очень небольшие порции энергии всего в 10—20 раз превышают kT. Кванты такой величины соответствуют инфракрасному излучению (длины волн около 3—1,5 мк), которое поглощается вследствие возбуждения колебательных степеней свободы многими органическими веществами. Казалось бы все в порядке — инфракрасное излучение может быть искомым источником энергии для наших целей. Однако оно сильно поглощается водой (недаром ИК- спектры органических молекул снимают обычно в безводных препаратах). Поэтому волны такой длины практически исключаются из числа возможных источников энергии. Следовательно, источником энергии может служить излучение, непоглощаемое водой. Значит, речь может идти о видимом свете (поэтому и видимом!) и об ультрафиолете. Короткий ультрафиолет также не годится, так как энергия соответствующих квантов сравнима по величине с энергиями связей в молекулах — будет происходить разрушение молекул.
Первичный источник энергии в биохимической эволюции. Вслед за В. П. Скулачевым [266] можно считать наиболее вероятным источником энергии во времена возникновения жизни свет. Этот вывод означает допущение первичности фотосинтеза, возникновения его в самом начале совершенствования систем энергетических превращений в биологических системах. Конечно же, речь не идет о процессе, подобном современному фотосинтезу. Ясно, что только избирательное поглощение излучения способно обеспечить энергией лишь определенные процессы, а не все реакции, как, например, при нагревании. В процессах, идущих в первичных матричных структурах, существование такой избирательности вполне вероятно. Для нуклеиновых оснований характерно сильное поглощение в области 260 нм, что соответствует (в расчете на 1 моль) порции энергии около 100 ккал. Это, конечно, слишком много, но для начала эволюционного совершенствования вполне терпимо. Основной результат поглощения света с такой длиной волны полинуклеотидными цепями — разрыв валентных связей [154], например, отрыв нуклеинового основания от рибозы, замена одного основания на другое. Вследствие интенсивных мутаций и обусловленных ими вариаций последовательности аминокислот в полипептидной цепи происходил отбор катализаторов, способствующих синтезу пигментов, которые поглощают видимое и ближнее инфракрасное излучение, соответствующее нужным квантам энергии.
Таким образом, действительный прогресс в эволюции биохимических механизмов преобразования энергии сопряжен с совершенствованием биосинтезов пигментных систем, преобразующих лучистую энергию света в химические формы энергии.
Нужно заметить, что и в этом направлении эволюция начиналась не «на пустом месте». Разнообразные пигменты легко синтезируются и без биохимических катализаторов из первичного бульона, содержащего аммиак, цианид, формальдегид, под действием например ультрафиолетового излучения. (Эти вопросы рассмотрены в ряде статей и в докладах на симпозиумах, по
священных происхождению предбиологических систем, а также в книгах А. И. Опарина [227, 228], Дж. Бернала [24], М. Кальвина [126] (см. также [1, 37, 53, 75, 127, 132, 240, 243, 250, 416, 448]).
Биологически наиболее важные пигменты — порфирины образуются -в среде, содержащей метан, аммиак, воду и водород, под действием электрического разряда и ультрафиолета. Основной частью порфиринов являются пиррольные кольца. Еще в 1915 г. А. Е. Чичибабин показал, что пирролы образуются при пропускании ацетилена и аммиака через нагретую трубку. Примерно в то же время Мейер установил образование пирролов и при взаимодействии ацетилена с цианистым водородом. Конденсация пирролов в порфирины (тетрапиррольные кольца) также представляет собой обычный химический процесс.
Порфирины замечательны сильным поглощением света в областях 400 нм (полоса Соре, энергия жванта в расчете на 1 моль примерно равна 60 ккал), а также поглощения вблизи 550 нм (энергия в расчете на 1 моль около 45—50 ккал). Такие энергии, по-видимому, наиболее удобны для наших целей — они уже недостаточны для непосредственного разрушения молекул и более чем достаточны для сдвига химического равновесия. Приняв кпд процесса фотохимического преобразования энергии равным приблизительно 50%, мы получим за счет таких порций энергии увеличение концентрации продуктов реакции, идущей вопреки термодинамическому градиенту, в 1020 раз, что заведомо достаточно для синтеза любых необходимых веществ.
Осталось совсем немного — найти способ преобразования энергии поглощенного света в энергию, обеспечивающую специфический сдвиг равновесия реакций синтеза полинуклеотидной или полипептидной цепи. Для этого нам нужно прежде всего рассмотреть способы смещения термодинамического равновесия в химических превращениях.
Возможные способы смещения равновесия в эндэргонических химических реакциях. Самый очевидный способ смещения равновесия данной реакции — осуществление сопряженной реакции, уменьшающей концентрацию одного из продуктов реакции или увеличивающей концентрацию по крайней мере одного из реагентов. Этот простейший механизм сопряжения — основа химических синтезов.
Так, например, очевидно, что выход при синтезе пептидных связей можно существенно увеличить путем поглощения воды, образующейся при конденсации аминокислот:
Н ОН
II I
СООН -f H-N-R2 ^R!-C-N-R2 -f H20. (1)
Однако в интересующем нас случае этот способ непригоден: реакция должна осуществляться в водном растворе.
Процесс синтеза пептидной связи изображен здесь не вполне верно. В реакцию вступают обычно ионизированные реагенты и ее нужно записывать следующим образом:
О ОН
II II I
Ri-C—O' + H3N+-R2 ^ Ri—С—N—R2 + Н20. (2)
Ионы в водных растворах сольватированы, т. е. окружены ориентированными вокруг них молекулами воды. Для разрушения сольватной оболочки нужно затратить энергию, и поэтому синтез пептидной связи из ионизированных реагентов термодинамически менее выгоден. Следовательно, сместить равновесие в сторону синтеза пептидной связи можно посредством уменьшения степени ионизации (диссоциации) реагентов. Это можно сделать, проводя реакцию в кислой или щелочной среде, чтобы подавить диссоциацию на ионы карбоксильной или аминогруппы. Доведение pH до нужной величины и будет способом внесения в систему энергии, необходимой для смещения равновесия проводимой реакции в нужном направлении. Однако по мере осуществления синтеза величина pH будет изменяться. При проведении синтеза пептидной связи в кислой среде последняя будет все более закисляться:
О ОН
II I
Rj-C-OH 1- H3N+-R2 <1 Rj-C-N-Rz + Н20 + Н+, (3)
при проведении реакции в щелочной среде — все более защела- чиваться:
О ОН
II I
Ri-C-O- + H2N-R2 ^ Ri-C-N -R2 + ОН'. (4)
И в том и другом случаях накопление конечных продуктов отразится на положении равновесия — выход синтеза пептидных связей будет меньше. Таким образом, мало довести pH исходных реагентов до требуемой величины, необходимо еще поддерживать оптимальное значение pH во время реакции. Способ поддержания pH общеизвестен — нужен рН-буфер. Что такое рН- буфер? Это система, в которой осуществляется непрерывная сопряженная реакция нейтрализации избытка образующихся ионов Н+ и ОН-.
Пример с синтезом пептидной связи довольно сложен — оба реагента ионизированы. Обратимся поэтому к столь же важному нам процессу синтеза полинуклеотидной цепи. Как известно, нуклеотиды связаны в полинуклеотидной цепи посредством фосфатных остатков. Рассмотрим образование этой связи с термодинамических позиций. Запишем интересующую нас реакцию при pH выше 7.
н
о
о
RI-CH,-0-P-CT + H0-C = RfiRI-CHr0-P-0 + ОН , (5)
® L©

О'
где R, и Rn — соответствующие нуклеотиды. Напомним, что ор- тофосфорная кислота имеет три гидроксила с резко различной способностью к ионизации: первый гидроксил диссоциирует при pH около 2,5, второй — при pH около 7,0 и третий — при pH около 13 (номера гидроксилов фосфорной кислоты отмечены цифрами в кружках). Ясно, что равновесие синтеза будет смещено вправо (в сторону синтеза) в кислой среде и источником свободной энергии для этого синтеза может служить рН-буфер, поддерживающий pH близким к 6,0.
Что могло бы служить в этой реакции буфером? Таких веществ с рК близким 7,0 не так-то много — фосфаты, имидазоль- ные производные, система карбонат — бикарбонат и, пожалуй, все. Таким образом, если в среде, в растворе, имеется какая- либо фосфорнокислая соль, например, однозамещенный фосфат калия, то ионы гидроксила, образующиеся при реакции синтеза межнуклеотидной связи, будут нейтрализоваться за счет сопряженно идущего процесса диссоциации второго гидроксила в буферном фосфате.
Итак, рН-буфер является источником свободной энергии, необходимой для смещения положения равновесия в эндэргоничес- ких реакциях полимеризации — синтезе полипептидов и полинуклеотидов.
Естественно, что эту энергию сначала нужно затратить при образовании буферной системы. Таким образом, обычный рН- буфер является хранилищем свободной энергии, используемой сопряженно с эндэргоническими реакциями. Такая роль буферных систем в биохимии чрезвычайно распространена, настолько широко, что ее обычно не осознают биохимики. Мы еще не раз будем обращать внимание на энергетическую, термодинамическую роль буферных систем в биохимических процессах.
Итак, мы могли бы сдвигать равновесие синтетических процессов в сторону синтеза за счет сопряженного осуществления экзэргонических реакций, а именно, за счет нейтрализации возникающих ионов ионами противоположного знака, накопленными предварительно в соответствующих буферных системах, и фотохимического преобразования энергии поглощенного света. Непосредственное использование энергии света, прямое фотохимическое сопряжение мало совершенны: синтезы могут осуществляться лишь при освещении. А ночью, в ненастье, в тени, в пещерах? Буферные системы более совершенны — они служат хранилищами теруолинамического потенциала, расходуемого по
мере надобности в любое время. Но тогда главной для нас оказывается проблема создания и наполнения буферных емкостей.
Запасание впрок световой энергии становится основной проблемой совершенствования энергетического обеспечения биохимических процессов. Такое запасание, естественно, может осуществляться лишь посредством накопления фотохимических продуктов.
Накопление рW-буферных систем при фотохимических преобразованиях энергии является одним из способов запасания энергии на первых этапах биохимической эволюции.
Преобразование энергии фотохимических превращений в свободную энергию ионных, кислотно-щелочных резервов можно осуществить лишь посредством окислительно-восстановительных процессов (см. [95]).
Рассмотрим наиболее вероятный в естественных условиях и простой вид буферной системы — соли угольной кислоты, карбонаты и бикарбонаты. Карбонатно-бикарбонатная смесь служит депо, хранилищем термодинамического потенциала для реакций, сопровождающихся изменением pH. Эта буферная смесь поддерживает pH близким к 8,0. Если реакция сопровождается образованием ионов водорода (например, при добавлении HCI), в буферной системе происходит преобразование карбоната в бикарбонат; если же в реакции образуются гидроксильные ионы (например, при добавлении NaOH), бикарбонат превращается в карбонат:
Na2COa + Н+ + Cl' -» NaHC03 + NaCl, (6)
NaHC03 + Na+ + ОН- Na2C03 -f H20. (7)
Попробуем придумать простейший преобразователь энергии света в свободную энергию буферной pH-системы. Я говорю «простейший», имея в виду заведомо добиологический, вероятный для первичного бульона в планетных условиях. Наиболее удобным процессом для указанной цели представляется окислительно-восстановительное превращение железа — восстановление и окисление ионов железа с изменением валентности железа от 3+ до 2+ и обратно. В чем особая привлекательность именно железа? В его распространенности и окрашенности его ионов и их комплексов, т. е. в способности поглощать излучение видимого диапазона, излучение, соответствующее приемлемым для наших целей энергиям. Иными словами, можно не придумывать для начала никаких пигментов: вещество, претерпевающее окислительно-восстановительное превращение, притом минеральное и широко распространенное вещество, само является пигментом, поглощает свет в нужной области спектра. Правда, у минеральных соединений железа есть недостаток—обычно они малорастворимы и легко гидролизуются. Зато железо очень легко ■образует растворимые комплексные соединения типа красной K3Fe(CN)e и желтой K4Fe(CN)6 кровяных солей.
Но сначала не будем педантично рассматривать химические процессы во всей их сложности. Начнем с раствора треххлористого железа и его восстановительного превращения в двухлористое:
FeCl3 + 1е~ -» FeCl, + С1~. (8)
Итак, мы располагаем раствором FeCl3 в воде. В том же водном растворе имеется и рН-буферная система, а то, что она необходима, мы сейчас увидим.
В нормальных условиях для превращения Fe3+ и Fe2+ нужно затратить энергию. Мы будем вводить ее в систему в виде света, поглощаемого раствором хлорного железа *. Возбужденный ион трехвалентного железа Fe3+ может превратиться в ион Fe2+, отняв у какой-либо молекулы среды один электрон. Отнять электрон в разбавленном водном растворе можно лишь у иона гидроксила, образующегося при диссоциации воды, который превращается в свободный радикал ОН, а в растворе остается неском- пенсированный положительно заряженный ион водорода Н+:
hv
Fe3+ + Н+ + ОН" К Fe!+ + Н+ + ОН. (9)
Свободный радикал ОН очень реакционно способен и может «успокаиваться» на-илучшим образом лишь соединившись с другим таким же радикалом, с образованием перекиси водорода:
НО + ОН -» Н—О—О—Н. (10)
Перекись водорода относительно стабильна. Однако именно в присутствии солей железа она разлагается на кислород и воду (в организмах эта реакция катализируется ферментом катала- зой):
2НА -»2Н20 + 02. (11)
Суммарный процесс с учетом стехиометрических соотношений реагентов запишется в виде
4hv
4Fe3+ + 4Н20 + 12C1--L 4Fe2+ + 12СГ + 4Н+ + 2На0 + 02. (12)
Четыре иона трехвалентного железа поглощают четыре кванта света и разлагают две молекулы воды на четыре протона и молекулу кислорода. (То обстоятельство, что по чисто стехио- метрическим соображениям в реакции должны участвовать четыре иона железа, кажется мне очень примечательным. Эта че- тырехтактность является условием осуществления перекисного промежуточного пути. Вероятно, именно поэтому -каталаза состоит из четырех субъединиц, четырех гемов и четырех полипептидных цепей. И гемоглобин состоит из четырех субъединиц...).
1 Фотохимия окислительно-восстановительных реакций с участием ионов Fe2+ и Fe3+, фотохимическое и радиационное разложение воды являются предметом ряда специальных работ (см. [8, 145, 156, 287, 345, 457]).
Процесс превращения трехвалентного железа в двухвалентное скоро прекратится вследствие накопления протонов, закислеиия среды. Поэтому условие относительно длительного протекания этой реакции состоит в наличии рН-буферной системы. При добавлении в раствор карбоната натрия получим:
4Н+ + 2Na2C03 -> 4Na+ + 2С02 + 2Н20. (13)
При взаимодействии четырех протонов с двумя молекулами карбоната натрия образуются две молекулы углекислого газа, выделяющегося из раствора, две молекулы воды и четыре иона натрия. Но поскольку в растворе присутствуют еще ионы хлора, в итоге получается 4NaCl. Однако при достаточном количестве молекул карбоната натрия углекислый газ не выделяется, а поглощается карбонатом натрия с образованием бикарбоната:
2Na,C03 + 2С02 + Н20 -» 4NaHC03. (14
Таким образом, итогом поглощения энергии света и превращения Fe3+ в Fe2+ оказывается накопление бикарбоната натрия, наполнение емкости выбранной нами буферной системы.
Возможности такой системы на этом не исчерпываются. Двухвалентный, двухзарядный ион железа Fe2+ можно превратить обратно в Fe3+. Последний процесс, хотя он осуществляется и самопроизвольно, может идти и при поглощении энергии света, сопряженном с разложением воды. В самом деле, превращение Fe2+ в Fe3+ сводится к отдаче электрона какому-либо подходящему акцептору. В разбавленном водном растворе возбужденный ион двухвалентного железа может отдать электрон даже протону, и тогда образуется свободный радикал Н — атом водорода. Радикал этот, естественно, неустойчив и реагирует с другим таким же радикалом (с образованием молекулы водорода Н2) «ли с каким-л,ибо иным подходящим реагентом. Напомним, что в нашей системе присутствует кислород (см. процесс [12]); тогда свободный радикал Н вступает в реакцию с кислородом (Н+02-^Нб2); затем образовавшийся Н02 вступит в реакцию с еще одним Н, и мы снова получим Н202. Знаменательно, что перекись водорода образуется в качестве промежуточного продукта в рассмотренных выше противоположных процессах. Перекись водорода разлагается с образованием Н20 и 02 и снова по чисто стехиометрическим соображениям в реакцию окисления должны вступать четыре иона Fe2+, поглощаться четыре кванта света, разлагаться четыре молекулы воды:
4 hv
4Fe2+Cl~ + 4Н20 + 202 X 4Fe3+ + 8СГ~ + 2НаОа + 40Н“ -> 4Fe3+ +
+ 8СГ- + 2Н20 + 02 -f- 40Н-. (15)
Как мы видим, при этой реакции в растворе остается избыточное количество ионов гидроксила ОН”. Ясно, что и осуществление реакции (15) зависит от рН-буферной системы, способнойпоглощать образующиеся гидроксильные ионы. Возьмем для простоты ту же карбонатную буферную систему:
4NaHC03 + 4NaCl + 40Н“ -> 4Na2COs + 4С1“ + 2Н20. (16)
Поскольку гидраты Fe3+ и Fe2+ поглощают излучение разных длин волн, нетрудно представить себе искусственную систему, в которой происходит попеременно то первая, то вторая реакция и цикл превращения железа замыкается. Цикличность этих превращений можно осуществить без попеременного изменения длин волн излучения света, а вследствие кинетических особенностей такой и аналогичных систем [102].
Радикалы, образующиеся в реакции превращения Fe2+ в Fe3+ могут реагировать не с кислородом, а каким-либо другим акцептором А, образуя АН2.
Соединим теперь реакции (12) — (16) в цикл:
4hv.
2
4hv2
О,
Мне представляется интересной эта выдуманная, искусственная неорганическая система преобразования энергии. В ней осуществляются процессы, аналогичные биохимическому фотосинтезу,— разложение воды с образованием кислорода и восстановлением акцепторов водорода. Примечательна ключевая роль перекиси водорода в обеих половинах цикла. Вместе с тем ясно, что стационарная концентрация перекиси водорода и других промежуточных продуктов может быть очень малой. Интересна необходимость четырех квантов света для прохождения половины цикла и восьми квантов для полного цикла. (Любители фотосинтеза должны взволноваться, так как пока никто не знает, почему для современного фотосинтеза необходимо восемь квантов света на элементарный цикл.)
При всем том, я не говорю, что именно так выглядела первичная, исходная в эволюции система преобразования энергии. Приведенное выше лишь служит иллюстрацией хода мысли, иллюстрацией возможности и необходимости сопряжения окислительно-восстановительных и ионных реакций при биохимических преобразованиях энергии. Наименее правдоподобно участие в этом цикле железа в виде его хлористых солей, но вполне вероятны аналогичные циклы для комплексных соединений железа,например, красной кровяной соли и желтой кровяной соли '. При полном обороте цикла — четыре молекулы Н20 разлагаются на 4Н+ и 40Н“. Это соответствует изменению энтальпии в системе на ДН = 4Х 13 = 42 ккал/моль.
Если же акцептором водорода служит не 02, а какое-либо другое вещество-акцептор А, то цикл оказывается, например,, таким:
В таком виде этот цикл почти полностью аналогичен современному фотосинтезу: разлагается вода, и образуется восстановительный акцептор водорода. Осталось еще придумать способ сопряженного синтеза АТФ...
Будем считать, что мы убедились в принципиальной возможности запасания энергии света посредством создания буферной pH-системы. Суммарная буферная емкость — это мера количества свободной энергии, которое может накопиться в системе. Запасенная свободная энергия может расходоваться по мере осуществления эндэргонических синтезов. Вполне вероятно, что эта возможность была реализована в ходе эволюции. В процессе естественного отбора возникали специализированные ферменты, катализирующие реакции в циклах, подобных рассмотренному выше. Особую заботу эволюции вызывало, вероятно, совершенствование комплексов железа — отбор таких комплексов, области поглощения которых при изменении валентности железа отличались бы наиболее резко, а также отбор систем биосинтеза пигментов и др.
«Макроэргические» соединения. Фосфорилирование, сопряженное с фотохимическим окислительно-восстановительным процессом. Теперь настало время усомниться в совершенстве запасания свободной энергии в рН-буферных системах. Термодинамически здесь, по-видимому, все в порядке. Но кинетически — отнюдь не все. Изменение pH в некотором смысле аналогично изменению температуры — эти изменения не адресованы какой- либо одной реакции, а сразу многим. Таким образом, путь к совершенствованию есть «векторизация», т. е. переход от безадресно-
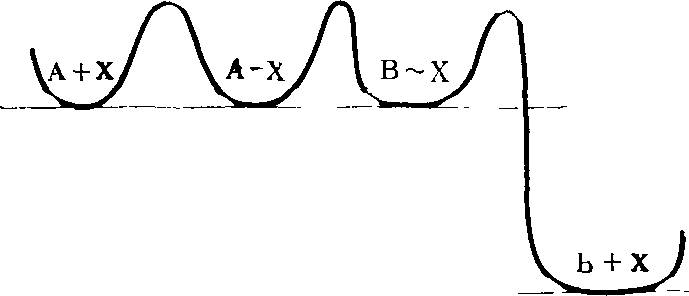

РИС. 11. Схемы, иллюстрирующие изменения свободной энергии в несопряженных и сопряженных химических превращениях

■а — несопряженное превращение А в В; б — образование промежуточного макроэргиче- ского соединения В^х сопряжено с превращением А в В; в — образование универсального макроэргического соединения х~у сопряжено с превращением А в В
го, статистического подвода энергии к снабжению ею только строго определенных процессов. Механизм такого избирательного энергоснабжения в принципе ясен — он должен основываться на специфических химических взаимодействиях. Запас энергии должен содержаться в химических группах, способных избирательно реагировать с превращаемой молекулой. Потом мы увидим, что этот запас в особых так называемых макроэргических группах также в значительной мере обеспечен запасом свободной энергии в рН-буфере. Пока же посмотрим, что это за группы. Что требуется от таких макроэргических групп? Необходимо, чтобы их отщепление от подлежащей превращению молекулы сопровождалось выделением энергии, обеспечивающей сдвиг равновесия соответствующей реакции в нужную сторону.
Главное назначение макроэргических групп — предохранение свободной энергии, выделяющейся в данном экзэргоническом процессе, от теплового рассеяния и обесценивания. Свое назначение эти группы выполняют посредством сопряженного с экзэр- гонической реакцией присоединения к промежуточным продуктам с образованием макроэргических соединений.
Когда молекула А превращается в молекулу В, свободная энергия системы уменьшается, т. е. выделяется энергия AF° (см. рис. 11, а). Если к молекуле А в реакции, почти не требующей затрат энергии, присоединена специфическая группа х образуется А^-х. Превращение А—х в В~х, в принципе, может не сопровождаться выделением энергии, уменьшением свободной энергии системы. Это эквивалентно тому, как если бы энергия превращения А в В сохранились в связи В~х — стоит разрушить эту связь — образуются свободные В и х и выделится в нашем идеализированном случае та же энергия AF°, что и при непосредственном превращении А в В (рис. 11, б). В силу сказанного связь В~х выступает как вместилище свободной энергии процесса превращения А в В, обозначается значком ~ (тильда) и называется макроэргической.
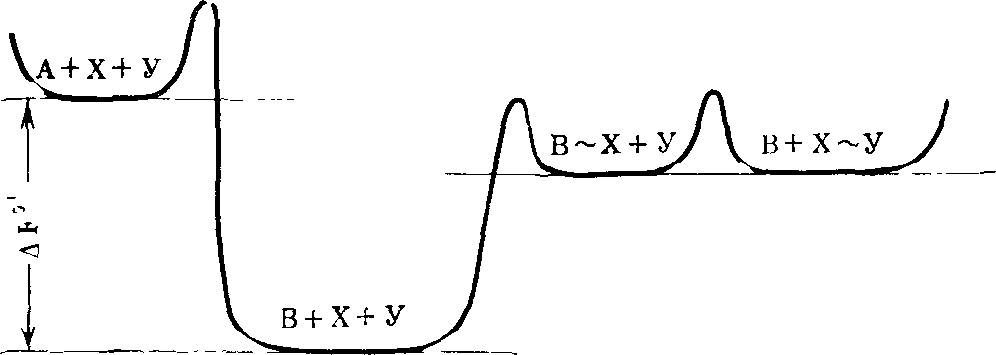
РИС. 12. Схема, иллюстрирующая возможность образования х~у при последовательном сопряжении с реакцией превращения А в В при стационарном режиме процесса
Макроэргическое соединение В ~х недостаточно удобно — оно определяется конкретной природой А и В. Термодинамический потенциал превращения А в В может быть сохранен от теплового рассеяния и перенесен для использования в других процессах посредством универсальных макроэргических соединений «х~у». Такиесоединения должны образовываться сопряженно с распадом В ~х\
В ~ х-{-у+±В-\-х ~ у.
В идеальном случае и образование х~у может идти изоэнергетически или с очень небольшим уменьшением свободной энергии системы (рис. 11, б).
Таким образом, свободная энергия, которая при несопряженном превращении А в В превращалась в тепло, теперь сохранена в связи хну
Представленная на рис. 12 схема энергетического сопряжения идеально высокоэффективна. Малая добавка вещества А приведет к образованию соответствующего числа (от добавленного Л) молекул у (Af°=0), т. е.:
_ Ml [х] [у] р '
Такое высокоэффективное сопряжение может быть чисто химическим, т. е. не требовать осуществления процесса посредством механических или электрических механизмов.
Менее совершенно сопряжение двух химических реакций посредством немакроэргического промежуточного состояния системы. Представим себе, что А сначала превращается в В, а затем В реагирует с х (см. рис. 12). Как показал Л. А. Блюмен- фельд [30], такой механизм сопряжения в нестационарном режиме малоэффективен, «импульс» увеличения концентрации А почти не дойдет до увеличения концентрации х~г/ — синтезируется лишь очень немного макроэргических молекул — в соответствии с величиной К„ реакции В + х+-В~х. Однако в стационарном режиме, при поддержании постоянной сверхравновесной концентрации А, сопряженный синтез х~у и по этому механизму может быть вполне эффективен, как эффективно перетекание жидкости в сифоне (см. схему, рис. 12).
Макроэргические соединения х~у могут служить источником энергии во всех процессах, где возможно их специфическое химическое реагирование. Этим и достигается «векторизация» траты термодинамического потенциала экзэргонических реакций.
Так, например, добавление х~у к В может привести к образованию в реакции В + х ~у^В~х + у, что никак не удалось бы при попытке провести реакцию В+х-^В~х. Таков энергетический механизм химических сопряженных реакций, открытых в начале нашего века Н. А. Шиловым [331].
Нужно специально подчеркнуть, что «макроэргичность» — это не характеристика свойств связи х и у самой по себе, а мера изменения свободной энергии в процессе превращения распада, гидролиза вещества х~у. Ясно, что величина изменения свободной энергии процесса (и даже знак этой величины) зависит от условий проведения процесса. Таким образом, микроэргическими называются соединения, распад которых (обычно гидролиз) сопровождается существенным уменьшением свободной энергии системы (ДF° около 10—20 ккал/моль) в данных условиях.
Макроэргические соединения должны образовываться за счет энергии, внешней по отношению к системе, например, за счет энергии света. Кроме того, они должны быть универсальными, т. е. годиться для энергетического обеспечения разных синтезов.
Биохимикам ясно, что сейчас речь пойдет в основном о пирофосфатах. Установление такой роли пирофосфатов — одно и. главных достижений биохимии XX в. (см. [37, 135, 157, 176, 265- 268, 297, 407, 420, 436]). Пирофосфаты представляют собой ан гидриды ортофосфорной кислоты. Неорганический пирофосфа образуется при нагревании, при прокаливании солей ортофос- форной кислоты:
О О 0 0
II II '|° I! II Т
НО-Р-ОН + НО + Р-ОНА НО-Р-О ~ Р-ОН + н2о.
I II
0~Na+ 0~Na+ 0'Na+ 0~Na+
Ангидридная пирофосфатная связь может образоваться между большим числом остатков ортофосфорной кислоты, тогд; возникают молеь^лы полифосфатов:
Пирофосфатная связь макроэргична в указанном смысле — при ее гидролизе в стандартных условиях свободная энергия системы уменьшается на 7—10 ккал/моль, т. е. гидролиз идет практически до конца. За счет уменьшения свободной энергии при гидролизе макроэргических связей и может осуществляться соединение, например, нуклеотидов в поликлеотидную цепь. Так, в реакции с участием пирофосфатаденозина (АДФ) происходит (за счет энергии, выделяющейся при расщеплении пирофосфатной связи) присоединение аденозинмонофосфорной кислоты АМФ к другому нуклеотиду, например ГМФ (реакция Грюнберг-Мона- го, Очоа):
АДФ + (ГМФ)П^: АМФ- (ГМФ)„ + Н3Р04.
Аналогичным образом можно сместить равновесие в случае синтеза пептидных связей при сопряженном расщеплении пиро- фосфатных связей в АТФ, образовании активированной, макро- эргической формы аминокислоты — аминоациладенилата. Пирофосфаты при нейтральном и щелочном значениях pH среды служат универсальными посредниками в превращениях энергии, необходимой для биохимических процессов. Однако создание больших концентраций пирофосфатных производных нуклеотидов невозможно: нуклеотидпирофосфаты активно участвуют в синтезе полинуклеотидов и их высокие концентрации могут повлиять на неконтролируемый синтез полинуклеотидов. Необходимы склады для хранения запасов термодинамического потенциала в менее активной форме.
Наиболее простой выход из создавшейся ситуации — хранение энергии в виде минерального пирофосфата или полифосфатов. Это и реализуется в клетках некоторых низших грибов и ряда бактерий (см. работы И. С. Кулаева [162]). В таких организмах хранилищем энергии оказываются минеральные полифосфаты (при числе ортсфоефатных остатков в цепи, достигающем 200— 300). От компактных осмотически (а, значит, и кинетически) неактивных частиц по мере надобности «откусываются» посредством специальных ферментов пирофосфатные группы для энергетического обеспечения биохимических процессов.
Однако эти «питательные камни» все же не очень удобны: мала поверхность взаимодействия со средой, мала предельная скорость вовлечения в метаболизм содержащейся в них энергии. Необходимы относительно мелкие, биохимически узко специализированные хранилища термодинамического потенциала.
В организмах животных эти функции выполняют гуанидин- фосфаты: креатинфосфат и (у беспозвоночных) аргининфосфат.
Ясно, однако, что хранение больших количеств энергии в виде гуанидинфосфатных и прочих макроэргических соединений также невозможно: на самом-то деле количество энергии, освобождаемой при гидролизе каждой связи, очень мало (всего около 10—20 ккал!моль). Таким образом, мы приходим к вполне тривиальному выводу — наиболее удобно хранить энергию в виде запасов пищи: жиров, углеводов, белков (см. с. 131).
Тем не менее, и в данном случае по кинетическим соображениям именно макроэргические соединения служат непосредственным источником энергии для эндэргонических биохимиче- ких процессов: свободную энергию из них можно получить в оптимальных дозах наиболее быстрым образом.
Универсальным в процессах превращения энергии в живых организмах на Земле веществом является аденозинтрифосфор- ная кислота АТФ *:

Представляется очень интересным установить причины, которые привели к выбору в процессе эволюции именно АТФ в качестве универсального посредника для подавляющего большинства видов превращения энергии в биохимических процессах. Возможно, что и в этом случае дело не столько в существенном, «деловом» превосходстве АТФ по сравнению, например, с ГТФ или ЦТФ, сколько в легкости спонтанного возникновения фосфорных производных именно аденина.
В соответствии с эволюционной логикой нашего анализа мы должны допустить, что образование АТФ сопряжено с поглощением света. Верность такого допущения можно оценить лишь при осуществлении в эксперименте синтеза пирофосфатных связей АТФ в относительно простой модельной системе при поглощении света.
Прежде, чем приступить к оценке возможностей такого эксперимента, попытаемся выяснить физико-химические причины макроэргичности пирофосфатных связей.
Вопросы эволюции биохимических механизмов преобразования энергии (см.
[37, 152, 268]).
О причинах макроэргичности пирофосфатов. Большая величина изменения свободной энергии при гидролизе пирофосфатов, т. е. их макроэргичность, проявляется в сильном сдвиге равновесия реакции гидролиза:
АТФ4" + Н20 ^ АДФ3- + HPOf- + н+
в сторону продуктов. Как известно, связь константы равновесия реакции К с величиной сопровождающего реакцию стандартного изменения в ней свободной энергии AF0 имеет вид:
AF° = — ЯГ In К,
где К — константа равновесия. Следовательно, для гидролиза АТФ можно написать:
[АДФ3-] [НРСЯ- [Н+]
AF = — RT In [АТФ4-] [Н20]
Однако в написанной выше реакции не отражена нейтрализация образующихся ионов водорода ионами гидроксила среды, т. е. реакция:
н++он-^н2о,
сопровождающаяся очень большим стандартным (AF°) изменением свободной энергии, равным примерно 20 ккал/моль. К сожалению (или к счастью), в биохимических системах отсутствуют сильные щелочи и кислоты — скрытые резервы ОН- и Н+ ионов представлены какой-либо буферной системой, т. е. обычно слабыми кислотами и их солями.
Мы уже видели, что в молекуле ортофосфата содержатся три группы, способные диссоциировать с образованием иона водорода. Однако соответствующие рК резко различны. Первый гидроксил (тот, который успевает диссоциировать первым), диссоциирует на 50% при pH около 2,0 (т. е. его рК равна 2). рК для второго гидроксила составляет 7,0, для третьего — около 13. Таким образом, при pH около 7 полностью диссоциирован первый гидроксил, наполовину — второй и совсем недиссоциирован третий. Ниже приведены структурные формулы орто-, мета- и пирофосфата:
О
И
н+о-рч
п © Ч° о о 'jj' Метафостфат ^ ||
н-о-р-о" н-о-р-о-р-о^
© I .Ф © I ® I Р
^ сгн+ о~ СГН+
© © (D
Ортофосфат Пирофосфат
Стандартное изменение свободной энергии ДF0, т. е. при единичных концентрациях исходных реагентов, нормальном давлении, £=25°, рН=7.
В пирофосфате «не хватает» второго гидроксила — он несет 2,5 отрицательных заряда вместо 3,0, как было бы в двух ортофосфатах. При гидролизе появляется недостающая половина заряда и соответствующее количество ионов водорода нейтрализуется ионами гидроксила среды. Возьмем для конкретности прежнюю карбонат-бикарбонатную буферную систему и запишем реакцию гидролиза АТФ подробнее, с учетом нейтрализации образующихся ионов водорода в буферной системе. При гидролизе пирофосфатной связи появляется новая кислотная группа — второй гидроксил ортофосфорной кислоты:
оо о о
RO-P-0-P-Cr +H305=2-R0-P-0 + H++H-0-P-Cr
®i-®i-® ®£-о © i-®
® © ® ®
Выпишем отдельные стадии процесса гидролиза и найдем для каждой константу равновесия.
Первая стадия записывается в виде:
АТФ4- Н20 ^ ЛДФ3- + Н2Р04 ;
_ [АДФ»~] [Н2Р07]
Kl ~ [АТФ4-] [Н20] ■
Во второй стадии образовавшийся ортофосфат диссоциирует:
[Н+] [НРОП
H2POJ ^ Н++ HPOf; Кг
[Н2Р04]
[СОГ] [Н+] [НСО-]
В третьей ион Н+ реагирует с ионами С032~ карбонат-бикарбо- натной буферной системы:
С02~ + Н+ ^ НСО;; К3 =
Поскольку [С032-] значительно больше [НР042-], концентрация Н+ определяется лишь свойствами буферной системы и равна:
К3 [HCOJ]
[НЧ - -
[С032~]
Учитывая приведенные выше соотношения для [Н+] и [Н2Р04_], получим ДЛЯ Ki\
К3 [НСО-] [НРО2'] [АДФ3~]
К1
х, [СО2-] [АТФ4-] [Н20]
Поскольку концентрация воды в водных растворах постоянна и равна 55,5 М, имеем для константы равновесия R суммарного
кг [ЧСО~] [HTOJ-] [АДФ*-]
К = Кх[ Н20]^-2 4
К9 [СО®-] [АТФ4-]
Из справочников находим /С2 = 1,6-10_7 и /С3г= 5,6• 10~и, откуда в данных условиях
К = К,-1,6-105.
Иными словами, для AF получаем:
AF = - 2,3RT lg Ki — 2,3RT lg (1,6 • 105) =
= — (2,3RTlgKi-t 7,3 ккал/мэль).
Как мы видим, изменение свободной энергии, вызванное нейтрализацией продуктов гидролиза и наличием воды в большой концентрации, весьма заметно. Правда, мы взяли буферную систему, поддерживающую сильно щелочную реакцию среды.
Для реакции нейтрализации ионов водорода, например, в фосфатном буфере, где в наших обозначениях Кг=К3, имеем:
^ = K,[HsO].
Нейтрализация продуктов гидролиза в этом буфере не дает выигрыша в энергии, но становится очевидной роль высокой концентрации воды
AFHs0 = - 2,3RT lg [Н20] = - 1,4 ■ 1,74 ж — 2,4 ккал/моль.
Для того, чтобы оценить R, нужно знать Л’,. Величина Ki зависит от различий свойств молекул соответствующего пирофосфата и продуктов его гидролиза. Рассмотрим поэтому еще раз процесс гидролиза пирофосфатов
О О о о
[I II II II
Н—О—Р-0 ~Р—О- 4 н2о -> Н-О—Р—О- + Н-О-Р—о- + н+.
! I I I
О- О- о- о
При гидролизе пирофосфата рвутся связи Р—О (пирофосфат- ная связь) в пирофосфате и О—Н в воде и образуются снова те же связи Р—О и О—Н, но уже в ортофосфате. Для того, чтобы судить об энергетических изменениях при таком гидролизе, нужно знать различия в энергиях этих связей в пиро- и ортофосфате, т. е. знать эффекты, обусловленные влиянием третьих атомов на энергию связи двух. Эффекты эти невелики и оцениваются 2—3 ккал/моль.
Связь Р~0 в пирофосфате менее прочна, чем связь Р—О в ортофосфате, т. е. термодинамически менее выгодна; при ее образовании выделяется меньше энергии, чем при образовании такой же связи в ортофосфате. Причины ее меньшей прочности, меньшей стабильности были рассмотрены в 1939 г. Г. Калька-
ром (см. [407]), в 1941 г. Ф. Липманном >[420], в 1950 г.— П. Оспером [438] и т. д. В пирофосфатной связи атом кислорода, соединяющий (или, если угодно, разделяющий) два фосфатных радикала, препятствует резонансному усреднению л-электронной структуры ортофосфата и возникает ситуация «Беспрепятственного резонанса» (opposite resonance). Квантовомеханический резонанс сопровождается выделением энергии, он термодинамически выгоден [233, 279]. Нарушение резонанса обусловливает термодинамическую нестабильность: гидролиз устраняет препятствия для резонанса и возникают более выгодные связи Р—О в ортофосфате. Приведенная схема иллюстрирует сказанное:
о
/и
/\\
о

НО—Р^;о-Н
В ортофосфате я-электроны двойной связи растекаются па всем четырем связям фосфор-кислород в результате резонанса структур, что выгодно термодинамически. В пирофосфате резонансное растекание л-электронов нарушено на кислородном мостике. Поэтому при гидролизе пирофосфата уменьшается свободная энергия, образуются более стабильные продукты. Все эти эффекты, насколько мне известно, не были до сих пор оценены с достаточной точностью — о их величине можно судить лишь на основании косвенных, главным образом экспериментальных, данных.
Из экспериментальных данных ДF гидролиза пирофосфатных связей оценивается в 5—8 ккал1моль в зависимости от условий [362], и поэтому на долю ДFa (т. е. К\) остается около 2 ккал[ /моль.
Таким образом, нейтрализация продуктов гидролиза и высокая «концентрация воды в воде» обусловливают основной вклад в изменение свободной энергии гидролиза пирофосфатной связи
[455].
Макроэргичность пирофосфатов определяется в значительной степени значением pH, поддерживаемым в среде, т. е. величиной буферной емкости. Расход свободной энергии, накопленной в рН-буферной системе векторизуется, т. е. направляется в нужную реакцию посредством сопряжения гидролиза пирофосфата с тем или иным эндэргоническим процессом.
Рассмотренные выше причины макроэргичности нельзя считать единственно возможными.
Так, основной вклад в изменение свободной энергии при гидролизе гуанидинфосфатов (креатинфосфат, аргининфосфат), по-видимому, вносит протонирование освобождающейся NH2- группы гуанидина и снятие запрета на резонанс структур:
NH H 0 NH О
I II II II
C-N ~ P—0~ 4- H20+ H+ C-N+H3 -1- HO-P-O-.
II
R O' R O-
Естественно, что изменение свободной энергии при гидролизе этих соединений больше в кислой среде (а не в щелочной, как при гидролизе пирофосфатов).
Особым примером макроэргического соединения, рассмотренным еще Калькаром и Липманном, является фосфоенолпируват. При его гидролизе образующиеся продукты дополнительно не ионизируются:
соон о соон о
II I II
С—О-Р-О- + Н20 С-ОН + H0-P-0-.
Н I I! I
СН2 О- СН2 О-
Основной вклад в макроэргичность вносит превращение термодинамически нестабильной енольной формы пирувата в стабильную кетоформу.
В этом случае макроэргичность молекулы не проявляется в ее физических свойствах. Термодинамическая невыгодность существования молекулы выявляется лишь в процессе ее разрушения при гидролизе. Макроэргичность такого типа, характерная для пирофосфата, фосфоенолпирувата и гуанидинфосфатов особенно привлекательна для биохимических целей. Перечисленные выше молекулы кинетически устойчивы, запас энергии, содержащийся в них, выделяется лишь при катализе — специфическом ускорении реакции их гидролиза. Вероятно, именно в силу особой кинетической стабильности и достаточной макроэргич- ности пирофосфаты и заняли столь выдающееся место в процессах превращения энергии в биологических системах.
Мне здесь хочется особенно подчеркнуть, что запасания энергии в виде одних только пирофосфатов невозможно — нужно еще располагать соответствующей буферной системой, поддерживающей pH на должном уровне.
Фотохимический синтез АТФ. Займемся теперь поисками способов запасания энергии, например, света, посредством сопряженного с поглощением света синтеза пирофосфатных связей н образования буферных систем.
Из рассмотренных выше соображений очевидно, что сами молекулы ортофосфата или пирофосфата не пригодны для фотохимического сопряженного синтеза: они не поглощают свет в необходимом для этих целей диапазоне. Вообще-то здесь нужны вещества окрашенные, поглощающие видимый свет. Однако для начяля эволюционного совершенствования пригодились бы и молекулы, поглощающие в ближнем ультрафиолете. Естественно предположить, что такими поглощающими свет -молекуламимогут быть спонтанно возникающие в первичном бульоне молекулы нуклеиновых оснований и нуклеотидов. Уже удалось осуществить синтез фосфорных производных аденозина за счет энергии ультрафиолетового излучения, но в присутствии этилмета- фосфата и метафосфата [447, 448].
Каким образом энергию, поглощенную нуклеиновым основанием, можно направить на синтез нирофосфатной связи? Мне представляется чрезвычайно привлекательной гипотеза о механизме фосфоршшрования АДФ в АТФ, выдвинутая много лет назад Л. А. Блюменфельдом и М. И. Темкиным [28].

Эта гипотеза основывается на том, что при нарушении сопря жения в пиримидиновом кольце 6-аминопурина (аденина) резк< возрастает способность азота аминогруппы переходить в поло жительно заряженное четырехвалентное состояние (четвертич ный азот):
NH2
!
С N
/\/Ч
N С СН
| || | +1е- + 2Н+
С С N
/Ч/ I
Н N R
Изменение рК-аминогруппы, например, в результате нар? шения сопряжения, как изображено выше, при восстановлен!! азота пуринового кольца с образованием свободнорадикально1 состояния углерода, соответствует изменению свободной энерги примерно на 10 ккал/моль, т. е. приблизительно на столью сколько нужно для синтеза пирофосфатной связи из ортофосфг тов. Мы видим здесь яркий пример преобразования энерги, окислительно-восстановительного превращения в энергию ионных взаимодействий.
Нарушение сопряжения в пуриновом кольце может возникнуть в результате фотохимического восстановления кольца. Для этого нужен довольно большой квант энергии света. Во всяком случае поглощение излучения с длиной волны 260 нм (энергия около 100 ккал!моль), характерное для пуриновых и пиримидиновых колец, заведомо обеспечивает возможность нарушения сопряжения в кольце — величина этих квантов даже излишне велика.
Однако самое главное здесь — представить себе, каким же образом осуществляется сопряжение, т. е. синтез пирофосфатной связи, наступающий вслед за восстановлением пуринового кольца аденина. Согласно Блюменфельду и Темкину, дело обстоит следующим образом.
Возникновение положительного заряда в аминогруппе пуринового кольца должно способствовать фиксации отрицательнозаряженного неорганического фосфата и, вероятно, также фосфатного конца молекулы АДФ или АМФ. При этом соответствующие нуклеотиды приобретают скорпионоподобную конфигурацию:
О
It
+
0
1
0-
1
о
X
сг
1 +
О NH,
1 *
I °-
О
1 II
\ О
.р'
о
/
о
-сн2-Хн н
N
сн
N
н
\
н
он он
Перенос водорода от свободнорадикальной формы аденозин- ;рифосфата на какой-либо подходящий акцептор должен сопровождаться значительным уменьшением свободной энергии — выделением энергии, способной превратиться в работу. При этом восстанавливается сопряжение в пуриновом кольце (свободно- радикальное состояние исчезает) и за счет выделяющейся свободной энергии синтезируется нормальная молекула АТФ:
t н Ф
hl I i
АДФ + фосфат -\- Н = донор ^ (АДФ)’ -)- (окисл.) донор,
Н (АДФ)- фосфат + акцептор -» АТФ + Н = акцептор.
Таким образом, синтез АТФ, сопряженный с поглощением света, оказывается двухтактным, двухстадийным; в первой стадии поглощение света приводит к возникновению свободнорадикальной формы АДФ, восстановленной водородом, поставляемым неким донором, и к образованию комплекса этой активной АДФ с молекулой ортофосфата; во второй стадии происходит рекомбинация свободнорадикальной формы пуринового кольца с сопряженным замыканием пирофосфатной связи, сопровождающемся отдачей лишнего водорода некоему акцептору.
Фантастична ли нарисованная нами картина? Выглядит она весьма привлекательно и термодинамически, по-видимому безупречна. Однако термодинамике угодить легко: ей безразличны пути реализации ее предначертаний. А что происходит на самом деле? Исчерпывающего ответа еще нет. Однако уже много лет назад Л. П. Каюшин и его сотрудники {36, 1316] обнаружили свободнорадикальную форму промежуточного продукта при гидролизе АТФ (а это делает вероятным появление такой формы и при синтезе).
Вскоре после опубликования гипотезы Блюменфельда и Темкина были проведены исследования, показавшие реальность окислительно-восстановительных превращений аденина (в частности, входящего в состав АТФ) [153, 270, 170]. Возможность сопряжения окислительно-восстановительных превращений аденина и синтеза АТФ была рассмотрена и в книге В. П. Скулаче- ва [266],
Таким образом, можно ожидать в близком будущем осуществления фотохимического или электрохимического модельного синтеза АТФ из АДФ и ортофосфата в присутствии подходящих доноров и акцепторов водорода (электронов) [267]. Мне кажется полезным попытаться теперь соединить две наших гипотетических схемы — схему процессов фотохимического разложения воды и создания буферной емкости при посредстве каких-либо соединений железа типа красной и желтой кровяной соли и схему фотохимического синтеза АТФ.
Выше уже отмечалось, что осуществление фотохимических реакций за счет энергии относительно короткого ультрафиолета биологически нецелесообразно; в самом деле, при таком облучении легко наступает разрушение уже синтезированных молекул и естественно ожидать возникновения в ходе естественного отбора биосистем, использующих энергию видимого света. Поэтому синтез пирофосфатов, сопряженный с поглощением света непосредственно пуриновым или пиримидиновым кольцом, должен вытесняться синтезом за счет энергии света, поглощаемого какими-либо окрашенными веществами — пигментами. Простейшими, способными к необходимому преобразованию энергии окрашенными молекулами являются соединения железа. Поэтому имеет смысл выяснить, вероятно ли образование свободнорадикальной формы активной АДФ, сопряженное с фотохимическими превращениями соединений железа. Мы видели выше, что переход Fe2+ в Fe3+ может происходить с передачей электрона иону водорода с образованием свободнорадикального водорода. Мы считали, что акцептором этого водорода служит молекула кислорода и образуется перекись водорода.
Представим себе теперь, что акцептором Н оказывается пуриновое кольцо АДФ, т. е. идет реакция Н +АДФ-v (АДФ)" и образуется активная АДФ. Соединяясь с ортофосфатом она должна отдать лишний водород и вернуться в нормальное, а не свободнорадикальное состояние. Для этого необходим соответствующий акцептор водорода. Им может быть тот же кислород и тогда образуется перекись водорода:
2Н (АДФ)' + 2Н3Р04 + 02 -» 24ТФ + Н202.
Объединив две наши схемы, мы получим следующее. Соединение трехвалентного железа (например, красная кровяная соль), поглощая свет, разлагает за счет поглощенной энергии воду и образуются двухвалентное железо (желтая кровянаясоль), ионы водорода, (наполняющие буферную емкость) и кислород. Двухвалентное железо при поглощении света (или спонтанно, в темноте), окисляется в трехвалентное железо при взаимодействии с водой (разлагая при этом воду на ионы гидроксила,, наполняющие соответственную буферную емкость) и реализует сопряженный синтез АТФ из АДФ и фосфата.
Мы получили комплексную систему, осуществляющую и реакции фотосинтеза (разложение воды на водород и кислород, образование восстановленного акцептора водорода) и сопряженное (быть может, темновое) образование АТФ при «обратном» транспорте электронов. Примечательно обязательное наличие перекиси водорода по крайней мере в одной половине цикла. Важно еще подчеркнуть, что наш цикл отнюдь не сводится к транспорту электронов — мы имеем здесь и транспорт протонов, и процесс образования и распада перекиси водорода, и процесс разложения воды.
Я подчеркиваю последние обстоятельства потому, что и для биохимических систем фотосинтеза и окислительного фосфори- лирования у ныне живущих организмов на Земле, также вероятны все перечисленные выше особенности — и участие воды, и перенос протонов *, и существенная роль следов кислорода и перекиси водорода.
Выше неоднократно подчеркивалась необходимость наличия Н202 и 02 в качестве участников многоэтапных превращений энергии в биохимических системах. Такое участие кислорода п перекиси водорода представляется очень вероятным из самых общих соображений. Нужно отметить здесь, что проблема роли Н202 в биохимических процессах — одна из основных и старейших в биохимии. Огромный вклад в ее исследование внес А. Н. Бах. Однако в течение многих лет после его смерти в биохимии господствовало мнение о Н202 лишь как о нежелательном побочном продукте в процессах биологического окисления.
Сейчас положение изменяется. Появляются публикации, в соответствии с которыми и Н202, и 02 (часто в очень небольших концентрациях) служат необходимыми участниками основных процессов превращения энергии.
Становится все более вероятной картина, в соответствии с которой Н202 и 02 являются ключевыми веществами в процессах синтеза и использования макроэргических фосфатных соединений. Большой материал по этому вопросу опубликован С. Е. Ма- нойловым [195, 196]. Особенно существенно в связи с обсуждаемым здесь обнаружение синтеза АТФ сопряжено с разложением перекиси водорода под действием каталазы в модельной системе. В растворе, содержащем АДФ, глюкозу, фосфат, гексо-
Об этом свидетельствуют значительные изотопные эффекты в системе митохондриального транспорта электронов при замене Н20 на D20 [125, 1816].
киназу, при pH 7, после добавления перекиси водорода и ката- лазы наблюдалась убыль неорганического фосфата (25% от исходного) и синтез АТФ.
Следует отметить также, что в работах М. Н. Кондрашовой и Г. Д. Мироновой [147] показано, что Н202 и 02 обязательно нужны для осуществления и окислительного фосфорилирования, и АТФ-азной реакции в митохондриях. Важность этих фактов как самих по себе, так и для подкрепления изложенных мною общих соображений очевидна.
Мне хочется еще раз подчеркнуть очевидную мысль — ничто в ходе эволюции не может возникнуть внезапно, без связи с уже существующими механизмами. Не может появиться . аппарат энергетического обеспечения синтезов полинуклеотидов и полипептидов, не родственный уже отобранным в ходе эволюции химическим системам. Поэтому нам следует искать механизмы преобразования энергии, механизмы сопряжения эндэргонических процессов с реакциями превращения молекул, уже существовавших на ранних стадиях эволюции. Вот почему мое внимание и привлекает фотохимическое превращение именно фосфорных производных исходных матричных молекул — нуклеотидов.
В дальнейшем всемогущая эволюция шаг за шагом усовершенствовала древние механизмы. В фотохимические процессы вовлекутся уже рассмотренные кратко пигменты (скорее всего, порфирины). Отбор каталитических активных полипептидных цепей приведет к возникновению высоко совершенных ферментов. Самое трудное (и, может быть, самое важное) найти начало. Таким началом и представляется мне фотохимическое сопряжение, синтез пирофосфатов аденозина, сопряженный с поглощением света пуриновым кольцом \ Весьма вероятно, что для такого синтеза абсолютно необходимы ионы тяжелых металлов, железа, кобальта, меди, цинка. Установить это можно лишь экспериментально, что и было выполнено недавно при изучении первичного синтеза аминокислот и нуклеотидов из первичного бульона.
В соответствии с таким подходом следующим шагом после использования непосредственного сопряжения поглощения ультрафиолета пуриновым кольцом и синтеза АТФ должно быть создание в ходе эволюции аппарата синтеза АТФ, сопряженного с поглощением света комплексными соединениями железа, пор- фириновыми или иными пигментами и лишь затем создание биохимической системы сопряжения синтеза АТФ среакциями окисления углеводов и т. п. При этом вполне возможно, что фотохимические механизмы сохраняются и в темновых процессах, например, при сопряженном фосфорилировании в митохондриях (см. ниже).
Эта мысль давно высказывалась в ряде работ. Она лежит в основе экспериментов Пониамперумы и его сотрудников по синтезу АТФ [447, 448].
Таким образом, следуя логике нашего изложения, следует признать, что фотосинтез должен был появиться в процессе эволюции раньше, чем темновые процессы биохимического преобразования энергии.
Утверждению о первичности фотосинтеза, как я думаю, не противоречит ничто из известного ныне. Мы почему-то связываем относительно позднее в эволюции биосферы Земли образование атмосферного кислорода (см. [49, 126, 1276, 228, 240, 250, 367, 452]) с эволюционным возникновением фотосинтеза. Это неверно. Кислород выделяется при фотосинтезе лишь у высших растений и водорослей. Выделение кислорода в значительных количествах отнюдь не было обусловлено тем, что этот газ столь нужен нам для дыхания. Кислород образовывался первоначально как неизбежный побочный продукт, «экскрет», при радиационном фоторазложении воды. Лишь затем, по мере выработки механизмов запасания энергии в виде энергии связи органических молекул жиров, углеводов, белков возникла необходимость и стало возможным образование макроэргических пирофосфатов, сопряженное с процессами деградации пищевых молекул, т. е. процессами дыхания и брожения.
Необходимо напомнить, что степень макроэргичности существенно зависит от концентрации воды. Поэтому фотохимический сопряженный синтез макроэргических пирофосфатов значительно легче осуществлялся бы в неводной среде. Эти вопросы мы обсудим ниже.
Ожидание осуществления фотохимического синтеза АТФ в простых модельных системах оправдались уже во время завершения в 1975 г. работы над текстом этой главы (см. с. 124). В серии работ Н. В. Гончарова и В. Б. Евстигнеев осуществили синтез АТФ за счет энергии света, поглощаемого хлорофиллом, а также (!) p-каротином или феофитином, адсорбированными на окиси алюминия [72, 74]. Кроме того эти авторы вслед за С. Е. Манойловым осуществили синтез АТФ за счет энергии, выделяющейся при разложении Н202 под действием каталазы [71] и за счет энергии, получаемой при измерении pH в суспензии частиц окиси алюминия с адсорбированными пигментами- хлорофиллами, каротином, феофином в темноте [73]. Ввиду большого принципиального значения достижений Н. В. Гончаровой и В. Б. Евстигнеева рассмотрим их работы подробнее.
На основании собственных экспериментов на бесклеточных препаратах анаэробных серобактерий и давних результатов химиков, изучавших образование перфосфатов и пирофосфатов при электролизе в щелочной среде растворов ортофосфатов, авторы пришли к очень существенному предположению. Они допустили, что в определенных условиях за счет различных эндэргоничных процессов может происходить превращение ортофосфата в метафосфат, который далее легко образует пирофосфатную связь при
реакции с молекулами свободных или замещенных ортофосфатов:
HPOJ' -> РО3 + он-
О
II
R-0-P0J- + РО~ -» R-0-P-0-P-0-.
43 II
о- о-
Превращение ортофосфата в метафосфат в водных растворах при средних значениях pH существенно эндэргонично и может быть осуществлено лишь за счет свободной энергии, выделяющейся в сопряженных энзэргонических процессах. Сами авторы полагают вероятным механизм превращения ортофосфата в ме-' тафосфат, основанный на промежуточном образовании свободно радикальных форм: фосфорила, гидроксила атомарного кислорода. Мне не кажутся эти допущения вполне вероятными. Но это и не важно. Важен сам факт осуществления сопряженного фосфорилирования в очень простых системах, например, сопряженно с реакцией разложения перекиси водорода, катализируемой каталазой! [71]:
2Н202— -АДФ + Н3Р04
2Н20 + 02<- АТФ + Н20
Реакция разложения перекиси водорода резко экзэргонична: ДF° 59 ккал/моль Н202 {436], поэтому в принципе при разложе- йии моля Н202 могло бы образоваться из АДФ и ортофосфата 6-=-8 молей АТФ. Однако количественная характеристика этого процесса явно преждевременна. Само осуществление его является большим событием в биохимии. Синтез АТФ сопряженно с каталазной реакцией осуществлен в очень простой модельной системе — в растворе 7-10~2 М Н202, ЫО-2 М КН2Р04; 0,3- • 10-2 М АДФ в присутствии — 1 -10—6 М каталазы. Примерно 30% АДФ превращалось в АТФ (что было установлено прямым анализом при помощи хроматографии на бумаге).
Затем был осуществлен синтез АТФ за счет энергии света, поглощаемого хлорофиллом, адсорбированным на частицах суспензии окиси алюминия [72]. В 1977 г. эти же авторы опубликовали результаты опытов [74], в которых наблюдали синтез макроэргических фосфатов за счет энергии света, поглощаемого не только хлорофиллами а и Ь, но феофитином (хлорофиллом без атома Mg в центре порфиринового кольца и p-каротином). Модельное фотофосфорилирование было осуществлено также в очень простой системе — суспензии соответствующих пигментов в частицах окиси алюминия в водных растворах КН2Р04 и АДФ. В том же 1977 г. Н. В. Гончаров и В. Б. Евстигнеев сообщили [73] о синтезе АТФ, сопряженном с быстрым изменением pH в суспензии адсорбированных на частицах окиси алюминия хлорофилла в водном растворе КН2Р04 и АДФ и измененным pH от 4,0 до 7,8—8,4. Наблюдалось образование 0,4 мкмолей АТФ
-из 10 бывших исходно мкмолей АДФ, т. е. около 4% —выход продукта нельзя считать большим, но сам факт синтеза АГФ в этих условиях представляется чрезвычайно важным.
ЗАПАСАНИЕ ЭНЕРГИИ ДЛЯ ДЛИТЕЛЬНОГО ХРАНЕНИЯ.
БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ И УГЛЕВОДОВ.
ВОЗНИКНОВЕНИЕ ГЕТЕРОТРОФОВ. АНАЭРОБНЫЙ МЕТАБОЛИЗМ
Макроэргические соединения даже в случае предельно эффективного сопряженного с экзэргоническими реакциями их синтеза мало пригодны для длительного запасания больших количеств энергии — очень уж мала их удельная энергетическая емкость. Свободная энергия гидролиза макроэргических соединений равна 10—20 ккал/моль. Для синтеза макромолекулы среднего размера из нескольких сотен мономеров нужно соответственно в 2— 3 раза большее число сотен молекул типа АТФ (или креатинфос- фата). Накопление высоких концентраций макроэргических соединений физико-химически невозможно: высокая ионная сила, большое осмотическое давление и т. д. затрудняют сохранение необходимой конфигурации макромолекул. Нужны более компактные хранилища энергии. Один из видов таких хранилищ (уже упоминавшиеся выше полифосфаты) — полимеры ортофос- форной кислоты, остатки которой соединены ангидридными пи- рофосфатными связями. Однако эти «питательные камни», по- видимому, имеют принципиальный недостаток — при использовании накопленной в них энергии образуются те же высокие концентрации кинетически активных молекул фосфорной кислоты и множество катионов, связанных на полифосфате, как на поликатионите. И самое главное как мы видели, макроэргичность макроэргических соединений обусловлена в основном емкостью рН-буферных систем. Запасание больших количеств энергии в макроэргических соединениях возможно лишь при высокой концентрации рН-буфера, т. е. при высокой ионной силе большом осмотическом давлении и других «неприятных» физико-химических условиях. Таким образом, по самой своей природе макроэргические соединения не пригодны для запасания значительных количеств энергии. Процесс запасания энергии должен происходить не в результате ионных превращений, а вследствие превращений электрически нейтральных молекул.
Направление эволюционных поисков в связи со сказанным ясно — энергия должна запасаться в молекулах, окислительная деградация которых сопровождается освобождением большого количества энергии. Наиболее экзотермичны реакции полного окисления углеводородов:
СПН2П+2 + V2 (Зп + 1) 02 - п С02 + (п + 1) Н20.
Изменение энтальпии АН при таком полном окислении (сжигании) углеводородов составляет примерно 106 ккал на 1 мольизрасходованного кислорода. Синтез углеводородов, сопряженный с поглощением энергии, излучаемой Солнцем, можно считать пределом термодинамического совершенства запасания энергии. Кислород, необходимый для освобождения энергии, накопленной в синтезированных углеводородах, образуется в том же процессе их фотосинтеза (нужно, правда, удержать 02 до момента темнового окисления).
Однако углеводороды гидрофобны и, следовательно, сечение взаимодействия, т. е. поверхность, доступная действию ферментов, оказывается очень малым. Кинетика использования углеводородов весьма несовершенна (мы снова убеждаемся здесь в несоответствии термодинамического и кинетического критериев биологического совершенства). Для запасания энергии нужны гидрофильные вещества. Среди них на первом месте — жирные кислоты (особенно ди- и трикарбоновые) и углеводы. Синтез углеводов, сопряженный с фотохимическим разложением воды (аналогичным выдуманному нами процессу разложения воды при превращении Fe2+^=Fe3+), представляет наиболее рациональный способ запасания легко мобилизуемой энергии и вещества. Когда быстрое использование энергии не требуется, а нужно запасти ее и отправить на длительное хранение, может происходить фотосинтез жирных кислот с длинной цепью (их триглицеридов — жиров и масел) полисахаридов, белков (как, например, в семенах, клубнях ныне существующих растений).
Фотосинтезирующие первичные автотрофы казалось бы располагают всем необходимым для достижения предельного (пока лишь биохимического) совершенства. Каков же биологический смысл возникновения гетеротрофов, зачем они появились? И не просто возникли, а еще достигли столь высокого совершенства. В сущности и автотрофы—химеры; их вполне можно (мысленно, а иногда и в действительности) разделить на две части — фотосинтезирующую и потребляющую. Потребляющая часть авто- трофа и есть гетеротроф, еще неотделенный от автотрофа. (Зеленую эвглену можно заставить разделиться так, что хлоропласт останется лишь в одной дочерней клетке. Другая, бесхлоро- пластная клетка — это гетеротроф, нуждающийся в органической пище).
Итак, что же позволяет гетеротрофной части первичного автотрофа отделиться и перейти к самостоятельному, хотя и зависимому от автотрофа существованию?
Я думаю главное здесь — возможность завоевания ареалов, недоступных автотрофам. В самом деле, фотосинтезирующие автотрофы могут расселяться по лику планеты в местах с достаточно интенсивным потоком солнечного света. Гетеротрофы способны существовать всюду, куда поступают тем или иным способом питательные продукты жизнедеятельности автотрофов. Возможность почти неограниченного расселения гетеротрофов особенно широко реализуется при возникновении в ходе эволю-
РИС. 13. Общая схема аэробных превращений энергии в биосфере
Ei— энергия солнечного света; £2 — низкокачественная тепловая энергия; А— акцептор водорода; ~Р— макроэргические фосфаты
ции аппаратов перемещения в пространстве, о чем мы специально будем говорить в следующих главах. Разделение живых существ на две группы — автотрофов и гетеротрофов — обусловливает разветвление дальнейшего пути эволюционного совершенствования биохимических процессов — на одном пути происходит биохимическое совершенствование автотрофов, на втором — биохимическое совершенствование гетеротрофов (сначала гетеротрофной части холотрофа — первичного автотрофа).
При достижении предельного совершенства и автотрофов и гетеротрофов устанавливается замкнутый цикл преобразования веществ и энергии в масштабах биосферы планеты. Автотрофы за счет энергии поглощаемого ими света разлагают воду на водород и кислород. Водород соединяется с какими-либо акцепторами — в основном с С02 с образованием, сохраняющих энергию молекул пищи (фотосинтез). Кислород уходит в атмосферу. Гетеротрофы осуществляют обратный процесс — образуют воду, освобождая законсервированную ранее энергию при соединении кислорода с водородом органических молекул (дыхание) (рис. 13). Прохождение этого термодинамического цикла, естественно сопровождается деградацией энергии — «высококачественное» видимое и ультрафиолетовое излучение Е, в конце концов превращается в «низкокачественное» тепловое излучениеЕ2, т. е. в относительно низкотемпературный белый шум (см. рис. 13). При осуществлении такого цикла, реализующегося сопряженно с процессами деградации солнечной энергии, совершаются все процессы жизнедеятельности, вызванные к жизни (в буквальном смысле этого словосочетания) биологической эволюцией. Сведение всех весьма сложных биохимических процессов превращения энергии к разложению и образованию воды— вероятно, одно из самых сильных научных обобщений нашего века.
Нефть, уголь и кислород[10]. К счастью (для нас — дышащих кислородом), этот цикл замкнулся не сразу. Сначала фотосин- тетическое разложение воды преобладало над образованием воды в процессах дыхания. Гетеротрофы могут использовать кислород при его не очень низкой концентрации в атмосфере.
Замечательно, что стационарный уровень концентрации кислорода в атмосфере соответствует количеству необратимо разложенной при фотосинтезе воды.
Следовательно, количество биогенных углеводородов (и других органических веществ), накопленных в истории развития аэробной атмосферы Земли, должно быть эквивалентно количеству кислорода в атмосфере (которое, согласно оценке, составляет 1,5-1015 т).
Нефть и другие каустобиолиты являются «зеркальными аналогами» кислорода. Нефть, уголь, торф и другие образуются в результате фоторазложения воды и накапливаются по мере увеличения концентрации кислорода в атмосфере. Отсюда следует, что, если верна теория биогенного образования кислорода атмосферы, то полное окисление всех каустобиолитов приведет к поглощению всего кислорода атмосферы. Кроме того, их полное окисление приведет к образованию воды, и уровень воды в Мировом океане может повыситься примерно на 3 м. Эти рассуждения позволяют оценить запас по всей Земле органического вещества в каустобиолитах примерно в 1 • 1015 т.
Таким образом, до накопления в атмосфере достаточного количества кислорода гетеротрофы могли получать энергию лишь в анаэробных процессах (бескислородных). Анаэробные условия сохраняются в ряде мест и после установления стационарной концентрации кислорода в атмосфере. Это объясняется все теми же диффузионными ограничениями — в толще воды, при отсутствии перемешивания концентрация кислорода быстро падает — там могут жить лишь гетеротрофы, способные к добыванию энергии в анаэробных процессах. Практически анаэробные условия легко возникают и в «толще» многоклеточных организмов. Этим объясняется странное, на первый взгляд, обстоятельство. Широкое распространение анаэробных биохимических превращений веществ и энергии на протяжении многих сотен миллионов лет после установления стационарного уровня концентрации кислорода, а странным это могло бы показаться, поскольку анаэробные превращения во много раз менее энергетически эффективны, чем аэробное окисление (дыхание). Итак, высокосовершенные анаэробные процессы деградации пищевых веществ — условие существования гетеротрофов (и фото-автотро- фов в темноте). Анаэробные биохимические процессы типа гликолиза или брожения настолько совершенны и неожиданно сложны, что почти невозможно использовать метод их дедуктивного, «умозрительного» построения. В этом случае правильнее сразу посмотреть их действительные свойства. Гликолиз и брожение — прекрасный пример предельно совершенного решения конкретной эволюционно-биохимической задачи. Энергетический и химический смысл этих процессов — выделение свободной
энергии в результате внутримолекулярных перегруппировок и расщепления исходных молекул. При гликолизе молекула глюкозы превращается в две молекулы молочной кислоты и свободная энергия системы уменьшается на 50 ккал/моль:
С.Н120. -» 2QHe03, AF° = — 50 ккал/моль.
При полном окислении глюкозы в стандартных условиях имеем:
С„Н120„ + 602 -» 6С02 + 6Н20, AF* = — 688 ккал/моль
Сущность анаэробного процесса гликолиза состоит в перегруппировке атомов. Составляя баланс разрываемых и образующихся связей, находим, что превращение состоит в образовании одной связи С=0 и одной С—Н вместо одной связи С—О, одной О—Н и одной С—С.
Изменение энтальпии при таком превращении можно оценить по таблицам энергии связей (см. например, книгу Я. К- Сыркина и М. Е. Дяткиной [279]. Результаты оценки дают, что при разрыве связей расходуется 248 ккал/моль, а при образовании новых связей выделяется 240 ккал/моль. Учитывая неточность табличных данных, можно сказать, что при гликолизе изменение энтальпии молекул АН, равное 8 ккал/моль, пренебрежимо мало. Нас, однако, интересует изменение свободной энергии. По-видимому, главной причиной энергетической выгодности превращения глюкозы в молочную кислоту являются изменения энтропии (образование двух частиц из одной), диссоциация (рН=буфер!) и сольватация образовавшихся ионов карбоксила.
Процесс брожения аналогичен гликолизу и отличается от него лишь на последних стадиях. В итоге молекула глюкозы распадается на две молекулы этилового спирта и две молекулы С02:
CeH12Oe -» 2CsH6OH + 2С02, AF0 = — 54 ккал/моль.
Существенным отличием брожения от гликолиза является улетучивание углекислого газа. С02 выходит из реакции, в результате чего происходит еще больший (чем при гликолизе) сдвиг равновесия слева направо, и «бродящие» гетеротрофы могут осуществлять такой расточительный процесс и дальше. (Это в сущности, возмутительная расточительность—при образовании спирта теряется большая часть энергии молекулы глюкозы, которую можно было бы получить при полном сжигании до С02 и Н20).
Ясно, что брожение эффективно лишь в условиях, позволяющих улетучиваться углекислому газу. Поэтому оно осуществляется лишь микроорганизмами, живущими в относительно открытых средах — в мелких водоемах, на поверхности питательных субстратов; если же сосуд закрыт, то С02 насыщает раствор, и брожение останавливается. Так происходит при изготовлении шампанского и других шипучих вин. Может показаться, что и второй продукт брожения — этиловый спирт улетучивается, и равновесие еще больше сдвигается вправо. Недаром же этиловый спирт называется по латыни spiritus vini — винный дух. Однако, как это ни удивительно, спирт, образующийся при брожении, не улетучивается из водного раствора: скорость его испарения при умеренных температурах (15—20°) только приближается (снизу) к скорости испарения воды[11]. Так достигается предельная концентрация спирта в сусле (около 16%), останавливающая жизнедеятельность дрожжей и превращающая его в сухое вино.
Второе весьма важное преимущество брожения (в тех случаях, когда оно возможно) перед гликолизом состоит в том, что молочная кислота очень сильная, а угольная — слабая. Физиологически допустимая концентрация молочной кислоты при обычных запасах рН-буферов порядка 10-2 М, предельная концентрация спирта около 2—3 М, т. е. примерно в 50 раз выше.
Следует все же отметить, что речь идет о предельной концентрации для дрожжевых клеток. В клетках животных даже очень небольшие концентрации спирта вызывают нарушение деятельности поверхностных и иных мембран. Клетки теряют способность создавать ионные градиенты, генерировать нервные импульсы и т. д. Поэтому животные не могут воспользоваться некоторыми энергетическими преимуществами брожения еще и из-за устройства мембран своих клеток. Следовательно, процесс брожения все равно остается неприемлемым для животных. Поэтому достойна всяческого осуждения привычка иных гетеротрофов сохранять образующийся при брожении спирт и потреблять в чрезмерных количествах конечный продукт анаэробного катаболизма.
Так или иначе, но эти, пусть и энергетически малоэффективные превращения углеводов виртуозно совершенны с точки зрения химика.
В их совершенстве легко убедиться, рассмотрев даже простую схему гликолиза или брожения [176] (т. е. схему, не учитывающую своеобразия кинетического поведения системы, динамики взаимной регуляции отдельных этапов и т. п.). Ясно, что осуществление таких процессов, как брожение или гликолиз, требует решения в процессе эволюции ряда сложных задач. Главная задача — обеспечение возможно быстрого, однозначного и строго последовательного превращения веществ. Строгий порядок, точное выдерживание последовательности стадий в многоэтапных превращениях достигаются посредством строгой специфичности ферментов к своим субстратам. Из множества броунирую- щих в среде молекул фермент выбирает молекулу своего субстрата. Поэтому эволюция первичных гетеротрофов в основном состояла в совершенствовании специфичности ферментов. Итак, длительное, сотни миллионов лет сохраняемое, существование
анаэробных процессов объясняется диффузионными ограничениями доставки кислорода. Одним из способов функционирования в таких условиях свободноживущих организмов, как было показано в гл. 5, является ограничение размеров клеток. Облигатные анаэробы (т. е. живущие только в анаэробных условиях) и факультативные анаэробы по-необходимости — мелкие 1— 3 мк — клетки. Поскольку в анаэробных условиях, в процессах1 гликолиза в многоклеточных организмах в особенности, лимитирующим является процесс отвода конечных продуктов. Кинетически выгодно иметь возможно большую поверхность выхода этих продуктов из клеток. Поэтому, вероятно, образование каких-либо надмолекулярных комплексов полиферментных систем для гликолиза не дает физико-химических преимуществ — гликолиз осуществляется в гомогенных растворах ферментов и субстратов. Однако до сих пор точно неизвестно, имеется ли структурная организация процессов гликолиза и брожения у ныне живущих организмов.
ПРЕВРАЩЕНИЕ ЭНЕРГИИ ПОСРЕДСТВОМ АСИММЕТРИЧНЫХ МЕМБРАН. КОНЦЕПЦИЯ МИТЧЕЛЛА
Из изложенного ясно, что степень совершенства функционирования системы преобразования термодинамического потенциала экзэргонических реакций в термодинамический потенциал макроэргических соединений становится на ранних стадиях развития жизни фактором эволюции, критерием естественного отбора.
Совершенство образования и использование макроэргических фосфатов слагается из двух компонентов: кинетического — скорости соответствующих реакций, разнообразие и концентрации необходимых ферментов и термодинамического — эффективности (КПД) сопряженного преобразования энергии.
Кинетика этих как и других биохимических процессов определяется совершенством соответствующих ферментов и величиной диффузионных ограничений.
Термодинамическая эффективность сопряженных преобразований, хотя это и кажется на первый взгляд странным, определяется реальными путями, механизмами перехода системы из одного состояния в другое. В самом деле, мы видели, что Л может быть превращено в В при полном рассеянии — превращении в тепло выделяющейся энергии. Эта энергия до тепловой деградации может быть направлена на выполнение какой-либо работы! Таким образом, качество использования термодинамического потенциала, КПД его преобразования в полезную работу- Полностью определяется (до теоретического предела) реальным ide: ханизмом преобразования энергии. Таким образом биолбгиче* ская эволюция должна быть направлена на выработку предельно совершенных механизмов сопряженного образования и-иёпользО- вание макроэргических фосфатов. ...v.. ..v.zi:
т
Рассмотрим «а-приорные» свойства таких механизмов.
В сущности сам переход от накопления энергии в простых буферных емкостях к биохимии макроэргических соединений был решающим шагом к такому совершенству. Он, как мы видим, состоял в векторизации потока энергии от безадресного изменения pH к точно направленному переносу термодинамического потенциала к строго определенным молекулам и процессам посредством молекул АТФ.
От чего зависит эффективность сопряженного синтеза АТФ?
Вероятно, более всего эффективность сопряжения зависит от соответствия квантов энергии, поступающей в систему синтеза АТФ и порций энергии, необходимых для синтеза одной макро- эргической связи. В самом деле, при фотохимическом сопряжении кванты света относительно очень велики 100—40 ккал/моль, а для синтеза ~Р связей нужно около 10 ккал/моль. Следовательно, фотохимическое «одноактное» сопряжение не может быть эффективным. Такая же картина наблюдается при фосфорилиро- вании, сопряженном с поглощением кислорода и окислением, например, водорода. Эта реакция «гремучего газа» сопровождается выделением 55 ккал/моль образующейся воды. Общеизвестен выход из этого затруднения, найденный в ходе эволюции— Замена одноэтапных превращений — многоэтапными, в которых энергия выделяется относительно небольшими порциями. В результате процесс окисления водорода органических молекул до воды осуществляется посредством последовательной цепи «транспорта электронов» — системы окислительно-восстановительных переносчиков. Теперь возникает новая сложная задача — обеспечения синтеза АТФ сопряженно с окислительно-восстановитель- ными переходами разных переносчиков. Кроме того, создание системы переносчиков с идеальными перепадами термодинамического (окислительно-восстановительного) потенциала очень сложно — эффективность энергетических преобразований может быть низкой. Как мы видели, синтез АТФ из АДФ и Н3Р04 при нормальных pH сопровождается образованием ОН- — защела- чиванием среды.
АДФ»~ + HPOJ- -> АТФ4- + ОН-.
Процесс синтеза АТФ может быть продуктивным лишь при сопряженной нейтрализации образующихся ионов гидроксила в реакции нейтрализации: он- + н+^н2о,
Эта реакция экзэргонична. Величина АР ее вполне приемлема (~20 ккал/моль) для энергетического сопряжения с синтезом макроэргических фосфатов (AF0'~ 10 ккал/моль).
Мы видели, однако, что в реальных буферных системах с рК близким к рКг фосфатных групп нельзя обеспечить сопряженное изменение свободной энергии более чем на 3—5 ккал/моль.
MEMИРАНА Окзэргоннческие процессы (фотосинтез, дыхание), создающие градиент рН(
ПРАВАЯ СТОРОНА
ЛЕВАЯ СТОРОНА
-ч
АДФ
+
н3ро,
II
АТФ
ГТ20
I \
НО ^ И + он"
Нт + ОН
: Н,0
ОН - \ •
РИС. 14. Схема синтеза АТФ в асимметричной мембране за счет энергии градиента pH, создаваемого в процессах фотосинтеза, дыхания и т. д. в соответствии с концепцией П. Митчелла Из активного центра Н+— АТФ-азы, локализованной в асимметричной мембране, ио- иы Н+ могут диффундировать только направо; ионы ОН- только налево. Неиони- зоваиные молекулы АДФ, АТФ, Н3РО4 растворяются в гидрофобной зоне мембраны. Синтез АТФ в этих условиях из АДФ и НэР04 происходит почти изоэнергетически. В целом процесс осуществляется за счет изменеиня свободной энергии при образовании двух молекул воды: слева при нейтрализации иона ОН- в кислой среде, справа — иона Н+ в щелочной среде
Выход из этого, непреодолимого для биохимических реакций в гомогенных растворах затруднения, состоит в механизме, предложенном П. Митчеллом [426—428]. Сущность этого механизма состоит в проведении синтеза АТФ посредством асимметричной мембраны, обеспечивающей при синтезе каждой молекулы АТФ расход вместо одного иона Н+ — двух, что приводит к большему сдвигу равновесия в сторону синтеза и, соответственно к большей эффективности сопряжения.
Представим себе асимметричную фосфолипопротеидную мембрану, по разные стороны которой существенно различна концентрация ионов водорода (рис. 14). Пусть например, слева от мембраны раствор имеет рН=4, а справа рН = 8. Слева в растворе имеются АДФ и ортофосфат. В кислой среде они в заметной концентрации будут находиться в недиссоциированном, незаряженном состоянии. В таком виде АДФ и Н3Р04 легко растворяются в мембране и попадают в гидрофобный активный центр АТФ-синтетазы. Так, в гидрофобном окружении АТФ не макро- эргична, и мы примем, что синтез ее из АДФ и H3POt идет изо- энергетично (ДР^О).
АДФ + Н8Р04 ^ АТФ + Н20.
Мембрана асимметрична — из активного центра образовавшаяся АТФ может диффундировать только обратно, налево. Образовавшаяся вода из активного центра может диффундировать лишьпосле диссоциации — налево ионы ОН- (левая сторона мембраны, омываемая кислым раствором с избытком Н+ ионов заряжена положительно), направо — ионы Н+ (правая сторона мембраны, омываемая щелочным раствором с избытком ОН" ионов заряжена отрицательно). Н+ и ОН- из образовавшейся в активном центре АТФ — синтетазы воды вступают, соответственно слева и справа в две реакции нейтрализации. Запишем теперь весь прочесе:
й шаг НР042-+АДФ3- + 5Н+^Н3Р04+АДФ
й шаг Н3Р04+АДФ->в мембрану
й шаг Н3Р04+АДФ^АТФ + Н20 в мембране
й шаг в мембране АТФ->АТФ4~-)-4Н+ в среду слева
в мембране Н20->-Н+ в среду направо \
ОН- в среду налево
й шаг слева ОН_ + Н+ ^Н20
справа Н+ + 0Н-^Н20
Сравним теперь процессы синтеза АТФ в гомогенном буферном растворе и в асимметричной мембране при наличии градиента pH:
в растворе: НР042_ +АДФ3 + Н+^АТФ4~ + Н20;
в мембране: НР042-+АДФ3++2Н++0Н^АТФ4-+2Н20. Видно, что в механизме Митчелла за счет асимметричного распределения Н+, т. е. градиента pH, синтез АТФ сопрягается с образованием вместо одной двух молекул воды, тем самым достигается существенный сдвиг реакции в сторону синтеза АТФ.
В самом общем виде смысл механизма Митчелла — дополнительная векторизация потока энергии, обеспечиваемая строго ориентированной переносом веществ соответственно пространственному градиенту pH.
Таким образом, к рассмотренным ранее аспектам кинетических достоинств структурной организации биохимических процессов (гл. 5, с. 87) следует добавить увеличение термодинамической эффективности преобразования энергий. Синтез АТФ в соответствии с механизмом Митчелла, посредством специальной структурной организации осуществляется с высокой эффективностью, поскольку он сопрягается с двумя относительно «низкопотенциальными» реакциями — нейтрализацией 2Н+ за счет 20Н~ при относительно малых перепадах pH.
В асимметричную мембрану могут быть встроены переносчики электронов, пигменты, ферменты-синтетазы. Энергия света, поглощаемого пигментами (фотосинтез), или энергия, выделяющаяся при транспорте электронов от водорода субстратов к кислороду (дыхание), тратится в соответствии с механизмом Митчелла на создание градиента концентрации ионов водорода (или других ионов), что проявляется в возникновении разности pH (или электрических зарядов) по обе стороны мембраны.
Синтез АТФ сопрягается не с отдельными разнообразными
реакциями, а с одной и той же на всех этапах реакцией нейтрализации— образовании воды из ионов водорода и гидроксила. Это сопряжение катализируется универсальным ферментом: мембранной Н+АТФ-азой [271]. Разность электрического потенциала, создаваемая градиентом концентрации отличных от Н+ и ОН-, катионов и анионов, например, К+ и С1~ легко преобразуются в разность концентраций Н+ и ОН-:
К+ СГ ^к+он- СГН+
Н20 Н20 ^ н+ ОН-.
Накопление энергии в виде градиента концентраций «иных» катионов и анионов может быть очень удобным ввиду больших запасов этих ионов в протоплазме. Энергетическая эквивалентность градиентов концентраций ионов водорода и гидроксила и разных катионов и анионов создает особо совершенную кинетику включения и выключения сопряженного фосфорилирования— градиенты электрических зарядов в отличие от градиентов концентраций— относится к дальнодействующим — это особенно существенно при трансмембранной передаче энергии — диффузия через мембрану процесс относительно медленный. Диэлектрическая постоянная гидрофобной мембраны мала — соответственно велика напряженность трансмембранного электрического поля и высокоэффективна трансмембранная передача энергии электростатических взаимодействий. Электростатический механизм накопления свободной энергии экзэргонических реакций в силу дальнодействия градиентов электрических зарядов позволяет сопрягающей мембране функционировать как единое целое, эффективно используется все, в том числе весьма малые сами по себе вклады в создание разности электрохимического потенциала по обе стороны мембраны.
Верность термодинамической схемы, т. е. самого общего механизма, предложенной П. Митчеллом в 1961 г. неоднократно иллюстрировалась экспериментально. Одной из наиболее ярких иллюстраций является опыт Ягендорфа и Урайба, осуществивших в 1966 г. синтез АТФ в изолированных хлоропластах за счет искусственно созданного градиента pH [404]. С тех пор число экспериментальных работ, подтверждающих концепцию Митчелла, значительно увеличилось. Замечательным примером осуществления фосфорилирования за счет энергии света, преобразуемой в энергию градиента pH, являются процессы в пурпурных, солелюбивых бактериях [374]. В мембранах этих клеток содержится зрительный пигмент — родопсин. Поглощаемая им световая энергия используется для работы «протонного насоса», создающего градиент pH. АХ'Ф синтезируется за счет этого градиента. Большой вклад в выяснение общих механизмов сопряженного фосфорилирования и особенно идущего при посредстве асимметричных мембран внесен В. П. Скулачевым и его сотрудниками, [266—271]. (См. также [360, 360а, 415,418,419,463,470]).
Глава 8
ПРОБЛЕМЫ МОРФОГЕНЕЗА
Диффузионные ограничения скорости биосинтезов — предел биохимического этапа эволюции.
Их преодоление посредством формирования все более совершенной морфологии живых организмов и (или) посредствомаппаратов активного перемещения в пространстве.
Отсутствие необходимости в предположении об особом морфогенном поле.
Достаточность кодирования на всех уровнях сложности синтеза определенных молекул и времени их синтеза (временная организация).
Цветковые растения — пример морфологического совершенства.
Первый макроэтап биологической эволюции завершается при образовании предельно совершенной системы матричного воспроизведения, предельно совершенных катализаторов системы преобразования энергии и механизмов отграничения дискретных организмов от внешней среды.
С некоторого времени скорость все более совершенных процессов биосинтеза начинает лимитировать диффузия (см. гл. 5, с. 81).
Некоторый эволюционный потенциал, т. е. возможность увеличения кинетического совершенства, содержится в ускорении диффузии посредством создания более резких градиентов концентрации. Эту возможность мы уже рассматривали. Она реализуется при образовании структурно-организованных ансамблей ферментов, например, в митохондриях и хлоропластах. Однако таким способом, посредством «игры на градиенте», достигается скорее экономия материалов, чем макроскопический выигрыш в суммарной скорости процессов. Поэтому показателем дальнейшего прогресса становится скорость диффузии, скорость потока метаболитов, превращаемых в вещество данного вида. Прогресс можно теперь измерять в см/сек. «Диффузионный барьер» не может быть преодолен чисто химическим путем. Дальнейшее совершенствование биохимических механизмов (катализа, маршрутов реакций) оказывается бесполезным.
Ясно, что суммарный поток можно увеличить, увеличивая поверхность, через которую осуществляется диффузия. Предметом отбора становится форма организма, его геометрические характеристики. Наименее выгодной формой организма данной массы является шар, так как его поверхность минимальна, наиболее выгодной — нить. Следовательно, первый шаг эволюции в направлении морфологического прогресса должен состоять в выработке механизмов построения нитевидных организмов вместо шарообразных. Эта задача уже довольно сложна. Ведь нить не может быть длинной — из-за медленности диффузии нарушатся связи между ее частями. По-видимому, пределом совершенства в данном эволюционном направлении является многолучевой еж, организм типа известных из зоологии беспозвоночных солнечников. Но такая сферически симметричная многолучевая морфология предельно совершенна лишь в отсутствие градиентов пищевых веществ и энергии. Равномерно питательная во всех направлениях среда окружает лишь планктонные организмы (и то, если пренебречь, например, градиентами света). В большинстве же случаев существует строгая направленность потоков пищи и энергии, что приводит к более сложным морфологическим задачам. Их можно решить путем выработки геометрических структур типа ветвей, стволов, листьев. Этот путь, путь морфологического прогресса для преодоления диффузионных ограничений, привел к возникновению сложной и рациональной геометрии растений (деревьев, трав) и животных, ведущих прикрепленный образ жизни (кораллы, актинии).
Совершенно ясно, что морфологические задачи возникают в дальнейшем и в связи с другими эволюционными проблемами. Однако именно вследствие необходимости преодоления диффузионных ограничений должны выработаться все основные морфогенные механизмы. Понятно, что естественный отбор наиболее совершенных геометрических форм организмов может осуществляться лишь при наследственном закреплении сведений о наиболее удачных морфологических конструкциях. Следовательно, в матричных текстах должны быть записаны не только сведения о строении (свойствах) ферментов, структурных белков, биодетергентов и т. д., но и инструкции для построения данных морфологических, геометрических конструкций, форм. Способ кодирования все тот же — посредством определенной последовательности нуклеотидов в полинуклеотидной цепи. Проблема перевода нуклеотидного текста в пространственное взаиморасположение частей организма, его морфологию—центральная проблема современной биологии. Однако прежде чем перейти к этой увлекательной проблеме, нужно все же сказать, что морфологический прогресс — не единственный способ преодоления диффузионных ограничений. И даже не лучший. Значительно большим эволюционным потенциалом характеризуется совершенствование путем создания аппаратов активного перемещения в пространстве. Этим направлением мы займемся в следующей главе, а сейчас продолжим рассмотрение проблем морфогенеза *.




ФИС. 15. Гигантские одноклеточные водоросли [163]
a— Caulerpa prolifera; 6 — Botridium granulatum; в — Acetobularia medi- terrianea (/) и A. wettsteini (2) и вегетативный гибрид (1+2) — регенерация стебля (}), посаженного на ризонд (2). Видно, что форма зонта определяется ядром, находящимся в основании ризонда
Когда говорят о морфологии организмов, невольно возникает образ многоклеточного организма сложной формы. В многоклеточном организме морфология обусловлена главным образом пространственным расположением клеток друг около друга.
Однако некоторые вопросы преобразования наследственной информации в морфологические особенности можно сформулировать уже для одноклеточных организмов и даже для неклеточных— вирусов и фагов. В самом деле, многие одноклеточные организмы имеют весьма сложную морфологию. На рис. 15 изображены гигантские (для клеток) одноклеточные (точнее, неклеточные) водоросли каулерпа, ботридиум и ацетабулярия, на рис. 16 — сложнейшие из одноклеточных животных — инфузории разных родов и жгутиконосцы, замечательные своей уникальной морфологией. Уникальная и очень сложная морфология свойственна и отдельным органеллам одноклеточных животных, (рис. 17), а также изученным Н. К. Кольцовым сперматозоидам насекомых, пауков и ракообразных (рис. 18). О сложности морфологии вирусов и фагов можно судить по рис. 19.
Таким образом основные механизмы морфогенеза, вероятно,, в наиболее чистом виде можно надеяться выявить уже для одноклеточных и даже неклеточных форм жизни.
Морфологические структуры могут возникать в результате самосборки из соответствующих макромолекул. В последние годы было выполнено очень много исследований по самосборке биологических структур (см. [303]). Правда, основные данные были получены при изучении самосборки вирусов и их частей («деталей»), Большой обзор интересных сведений приведен в книге Б. Ф. Поглазова [237]).
Попробуем опять подойти к этому вопросу на основании общих эволюционных положений. Речь идет, следовательно, об отборе в процессе эволюции молекул, агрегация которых автоматически приводила бы к построению все более биологически целесообразных структур. Наиболее естественно было бы выбрать с этой целью белки-—вариация их аминокислотного состава и последовательности аминокислот заведомо обеспечивает любое необходимое разнообразие свойств молекул. Свойства молекул, синтезируемых нематричным путем (например, липидов или полисахаридов), могут варьировать в процессе эволюции лишь посредством значительно более громоздких механизмов. Для синтеза любой новой молекулы типа моносахарида или фосфолипида необходимо большое число строго специфичных ферментов. Таким образом, кажется вероятным, что когда потребовалось не просто отграничить клетку от внешней среды, но придать ей уникальную форму, для ее построения понадобились специальные структурные белки. Мысль эта находит подтверждение во всех случаях биоморфогенеза. Определяющая роль белков в морфогенезе на молекулярном уровне была выяснена в замечательных исследованиях самосборки вирусов ([см. 237]). Начало было положено при изучении вируса табачной мозаики (ВТМ). Этот вирус состоит из РНК (около 5% по весу) и белка. Частица ВТМ распадается на составные части под влиянием различных воздействий: разбавленной щелочи, концентрирован-
- В
РИС. 16. Одноклеточные — инфузории, жгутиконосцы, радиолярии а — Euplotes patella (инфузория); б — Poludlnium mysozeum (инфузория из кишечника слона); в —скелет Cycladopora pantheon (радиолярия); г — Heteronema spirale (жгутиконосец, эвглеиовые); д — Ceratium longlpes (жгутиконосец, перидиниевые); е — Gonyaulax polyedra (жгутиконосец, перидиниевые); а, б, в заимствованы из книги [921, остальные — ■из книги [163]
РИС. 17. Стрекательная капсула одноклеточного организма-споровика Nasema
Капсула имеет двойную оболочку и ввернутую в капсулу полную нить. Выстреливание обусловлено быстрым развертыванием нити
РИС. 18. Спермий длиннохвостого рака Galathea squamifera [144]
РИС. 19. Иллюстрация сложной морфологии вирусов и фагов [237]
а—модель строения вируса табачной мо-
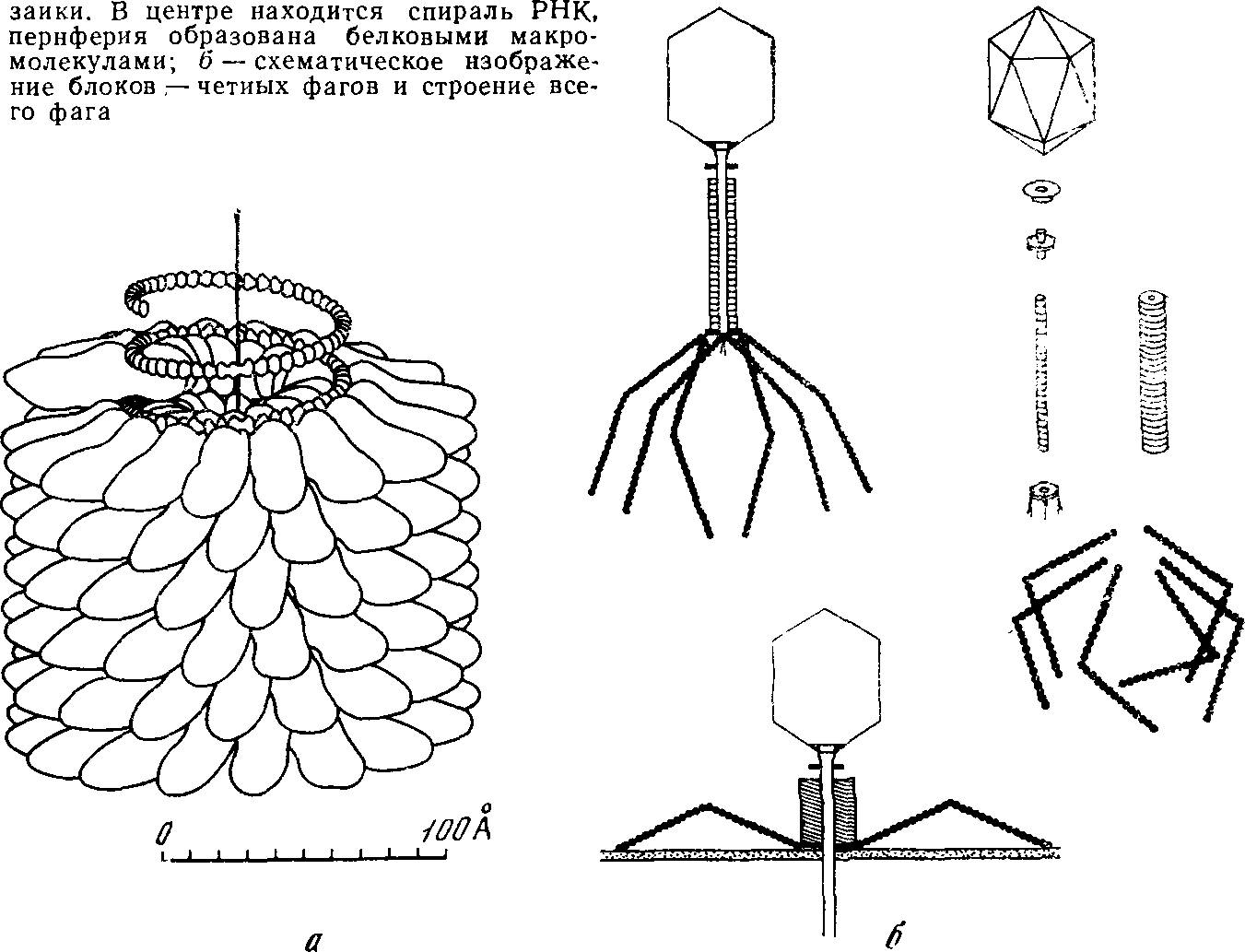
мой уксусной кислоты, мочевины, додецилсульфата и других агентов. При защелачивании среды до pH 10,5 происходит дезагрегация белковой оболочки ВТМ на небольшие субчастицы — А-белки. При подкислении щелочного раствора А-белка начинается упорядоченная агрегация, самосборка — образуются длинные палочкообразные структуры, внешне не отличающиеся от ВТМ. Чрезвычайное волнение вызвало в 1955 г. сообщение Френ- кель-Конрата о самосборке, реконструкции целого инфекционного вируса при агрегации А-белка в присутствии РНК, выделенной из ВТМ. Это было первое искусственное воссоздание (почти создание) живого организма. (Вирусы вполне подходят иод определение живого организма — объект биологической эволюции, находящийся или способный находиться в процессе био- .логической эволюции. А то, что вирусы в ходе эволюции обленились и разучились самостоятельно добывать, готовить, переваривать и использовать пищу — просто характеризует их очень ллохо...) И это воссоздание осуществилось посредством самосборки. Строение ВТМ показано на рис. 19, а. Видно, что самосборка структурных элементов может происходить и при не очень •сложных белковых субчастицах, субъединицах — они должны иметь специфические контакты, определяющие их соединение как друг с другом по спирали, так и с РНК. Успех искусственного осуществления самосборки ВТМ вызвал интенсивные исследования этого процесса на других вирусах. Было установлено, что в результате самосборки из белковых молекул образуются большей частью или полые цилиндры, или плоские мембраны, способные образовывать замкнутые сферы. Большие успехи были достигнуты при изучении самосборки мембраны малых сферических вирусов (237, с. 29], в частности, вирусов, вызывающих рост незлокачественной опухоли папилломы. Крик и Уотсон предположили в 1956 г., что из идентичных белковых молекул (морфологических единиц) могут возникать плоские мембраны, полые сферы, эллипсоиды, полые цилиндры (трубы) и их комбинации. Каспар и Клуг подтвердили их предположение посредством геометрического анализа (см. [237]).
Характер автоматически возникающей структуры зависит в основном от числа контактов, которые способны осуществлять имеющиеся в системе морфологические единицы. Число таких контактных точек в них в свою очередь зависит от числа образующих их белковых субъединиц.
Так, морфологические единицы, имеющие по шесть контактных точек и состоящие из шести субъединиц, образуют плоскую однослойную мембрану (рис. 20, а). Для образования из такой мембраны замкнутой поверхности многогранника она должна быть перегнута во многих местах. Образующиеся ребра делят поверхность на серию треугольников. Для этого наряду с шестиконтактными морфологическими единицами в системе должны находиться и пятиконтактные, состоящие из пяти субъединиц

РИС. 20. Модели, иллюстрирующие принципы самосборки надмолекулярных структур [237]
а — шестиконтактиые морфологические единицы, образуют плоские мембраны (одиа из единиц зачернена; б — пятикоитактные морфологические единицы (одиа из иих зачернена) образуют вершниы в ограненных мембранах
(см. рис. 20, б). Пятиконтактные морфологические единицы встраиваются в вершины многогранников, не нарушая непрерывной упаковки [237]. Наименьший многогранник состоит лишь из двенадцати пятиконтактных морфологических единиц. Различные сочетания пяти- и шестиконтактных морфологических единиц обеспечивают решение сложных морфологических задач.
Модель вируса папилломы, предложенная Финчем и Клугом (см. [237]), представляет собой икосаэдр из 72 морфологических единиц, образованных 420 субъединицами первого порядка. Если морфологические единицы образованы не только шестью п пятью субъединицами молекул белка, но и меньшим их числом, то при участии в построении надмолекулярных структур и неассоциированных субъединиц первого порядка мо^ут автомати- чески возникать разнообразные геометрические формы — трубы, эллипсоиды, икосаэдры (см. рис. 20). Еще более сложные морфологические структуры получаются при самосборке отдельных блоков Т-четных фагов (см. рис. 19, б). Самосборка этих фагов определяется большим числом генов — синтез белковых субъединиц контролируется 17 генами, около 30 генов регулируют самосборку образующихся белковых субъединиц и отдельных блоков фага [237].
Для наших целей совершенная экскурсия в область изучения самосборки вирусов достаточна. Мы видим, что морфология вирусов является следствием структуры, морфологии специфических белковых молекул. Теперь нужно попробовать выяснить, могут ли по тому же принципу строиться значительно более крупные образования типа клеток инфузорий, сперматозоидов или водорослей?
Интересующий нас вопрос можно сформулировать и иначе: является ли морфология клеток прямым следствием морфологии молекул белков? В таком виде этот вопрос эквивалентен вопросу о возможности многократного увеличения молекул с сохранением их морфологических особенностей. Подобная возможность кажется сомнительной. Форма и морфологические особенности зерен серебра никак не связаны с формой фотографического изображения. Ситуация с клеткой, ее морфологией и формой белковых макромолекул представляется сходной. Диаметр субъединицы белка порядка 10 А, диаметр клетки порядка 10 мк, т. е. их диаметры различаются на 4 порядка. (В фотографии диаметр зерна серебра тоже отличается от линейного размера детали изображения на 4 порядка.) Однако мы не удивляемся тому, что форма даже гигантских монокристаллов является следствием прямого увеличения элементарной кристаллической ячейки атомарно-молекулярных размеров. Но теперь взглянем на рисунки, на которых изображены инфузории, неклеточные гигантские водоросли— вошерия, ацетабулярия, ботридиум и др. (см. рис. 15, 20) уж очень их морфология отличается от морфологии кристалла. Очень уж эта морфология «неправильна» и «причудли-за». Однако на примере вирусов мы видели, что в зависимости •от строения морфологических единиц, а, следовательно, от набора не очень разнообразных субъединиц, автоматически образуются разные достаточно сложные морфологические структуры — полые сферы, эллипсоиды, трубки. Поэтому, если последовательность появления (биосинтеза) тех или иных субъединиц заложена в генетической программе, то можно представить себе последовательность образования очень сложных морфологических структур непосредственно из макромолекул белка.
Следовательно, морфология клетки и ее частей может быть .записана в нуклеотидном коде в виде текстов (генов), определяющих синтез в необходимой последовательности полипептид- лых цепей с определенной последовательностью аминокислот. Естественно, мы говорим сейчас лишь о принципиальной возможности такого механизма морфогенеза клетки. Весьма привлекателен и другой принцип морфогенеза на этом уровне. В самом деле, мы по существу рассматривали следующую модель. Набор разных кубиков разной формы с липкими гранями насыпают в ящик и какое-то время трясут (тепловое движение); кубики слипаются так, чтобы суммарная свободная поверхность, смазанная .клеем, была минимальной, т. е. образуется сложная морфологическая структура (эта модель была реализована Ферстером [303]. Затем в ящик добавляют кубики другой формы и снова трясут— прежняя морфологическая структура достраивается, превращаясь в структуру нового типа, и т. д. В этом случае реализуются структурные особенности морфологических единиц и их субъединиц, причем их реализация происходит при стохастическом взаимодействии структурных единиц.
Никаких кинетических особенностей, никакой кинетической организации пространства нам не требовалось. Дело в том, что кинетические особенности вполне могут приводить к пространственным геометрическим эффектам. Это обязывает нас поставить вопрос — не используются ли в биологических морфоген- яых процессах и кинетические механизмы? А если нет, то почему эволюция прошла мимо столь замечательных явлений?
По-видимому, первым изученным примером кинетического образования структуры были кольца Лизеганга. Они представляют собой правильные периодические структуры, возникающие при образовании осадка в результате реакции равномерно распределенного вещества с другим реагентом, диффундирующим в зону реакции. Кольца Лизеганга легко получить, если пропитать желатиновый слой фотопластинки хроматом калия, а затем капнуть на него раствор азотнокислого серебра. По-видимому, для образования кристаллического осадка хромата серебра необходимо наличие затравки, появляющейся лишь при некоторой пороговой концентрации хромата калия. Поэтому при диффузии ионов серебра через область, обедненную (в результате предыдущего осаждения и диффузии) хроматом калия, осадок не образуется. При достижении некоторого критического соотношения ионов серебра и хромата лавинообразно развивается процесс кристаллизации, с избытком ионов серебра реагируют ионы хромата из прилежащих областей (последние обедняются этим анионом). И вновь, при продолжающейся диффузии серебра в этих обедненных зонах не появляется осадок. Так образуются концентрические кольца — пустые кольца чередуются с кольцами осадка хромата серебра. Получение колец Лизеганга произвело большое впечатление на минералогов — концентрические структуры в яшме, малахите и т. п. стали рассматриваться как результат сочетания процессов диффузии и осаждения в период образования этих минералов. Позже, особенно после открытия (методом электронной микроскопии) тонкой структуры миелино- вого волокна с его концентрическими слоями миелина [417], стали высказываться мнения о возможном участии механизмов, аналогичных рассмотренным выше, и в образовании морфологических структур в биологических системах (например, структуры миели- нового волокна). Но мне кажется это очень сомнительным — нельзя «доверять» сферически симметричной диффузии тонкую морфологическую работу. Кроме того давно и точно установлено, что концентрическая структура миелинового волокна возникает при многократном наворачивании тонких блиноподобных клеток на аксон.
Одним из наиболее интересных примеров кинетического механизма организации пространства является возникновение пространственных структур в процессе распространения химической колебательной реакции. Такие реакции были капитально исследованы в нашей лаборатории [101, 102]. При реакциях, изученных А. М. Жаботинским и А. Н. Заикиным, образуются подчас весьма сложные узоры, обусловленные распределением в пространстве окисленных и восстановленных форм соответствующих катализаторов. Таким образом возможны кинетические по происхождению структуры, кинетическая организация пространства. Пространственные характеристики определяются в данном случае кинетическими параметрами — скоростями реакций, соотношением положительных (активация) и отрицательных (угнетение) обратных связей в реакционной цепи. Следовательно, пространственная организация структуры легко поддается регулированию. А это именно то, что совсем не нужно в морфологии (вернее, обычно не нужно). Морфология должна быть достаточно консервативной, инвариантной относительно не сильных модификаций условий ее возникновения. Мне кажется, что кинетический морфогенез не мог устроить эволюцию.
До сих пор мы говорили о морфологических (морфогенных) процессах на уровне клетки. Наиболее сложны, вероятно, проблемы морфогенеза многоклеточных организмов.
Многоклеточные организмы возникают из одной клетки — ?иготы. Морфология на этом уровне определяется взаимным рас
положением и взаимодействием клеток в пространстве. Значит, и в данном случае должен существовать однозначный механизм перевода нуклеотидного текста в план, чертеж постройки из клеток. В самом деле, что определяет соединение клеток друг с другом в строго определенных положениях? Можно представить себе несколько ответов на этот вопрос. Проще всего предположить аналогию в механизме образования морфологических структур из клеток и из макромолекул. Геометрия построек из клеток может определяться геометрией клеток — числом возможных контактов каждой клетки и ориентировкой контактных участков, их расположением на «шкуре» клетки. Как мы видели, сочетание гексамерных и пентамерных морфологических единиц приводит к возникновению разнообразных структур. Гексамерные, шестиконтактные клетки должны образовывать плоские структуры, двухконтактные — нити.
Другим возможным способом морфогенеза мог бы быть такой процесс дробления яйца (или любой клетки на поздней стадии морфогенеза), при котором взаимное расположение клеток определяется направлением митотического веретена и тем самым границы, разделяющей дочерние клетки. Последовательная переориентация митотического веретена в последовательных актах клеточного деления могла бы приводить к образованию любых сложных морфологических структур (такая возможность казалась особенно привлекательной А. Г. Гурвичу (76]). Однако указание на такой способ морфогенеза лишь видоизменяет задачу— все равно нужно выяснить причины той или иной ориентации митотического веретена при очередном делении клеток. Но самое •сильное возражение против подобного механизма, как я думаю, состоит в том, что морфология многоклеточных образований не определяется процессом размножения клеток. Это следует из замечательных опытов по реагрегации клеток после дезагрегации органов и тканей (и даже организмов)!
Такие исследования начались на губках. В начале нашего века было обнаружено, что если нарушить связи между клетками, «слегка помяв» кусок губки в морской воде, то происходит мацерация, и свободные клетки начинают плавать в морской воде. Однако очень быстро наступает реагрегация клеток — они собираются и вновь восстанавливается исходная типичная для тубок структура [215]. Аналогичный процесс реагрегации можно наблюдать при мацерации печени или почек эмбриона мыши или цыпленка. Реагрегация почки с ее сложной морфологией особенно впечатляюща — клетки разных типов располагаются в нужных местах, образуя характерные морфологические структуры. Следовательно, в этих случаях взаимное расположение клеток не •определяется ориентацией митотического веретена. В процессе реагрегации клетки активно перемещаются друг около друга, а их мембраны совершают волнообразные движения. Как только жлетки оказываются в «правильном» положения, их движенияи колебания резко прекращаются; такая реакция остановки является существенным элементом формообразования в многоклеточных постройках [47, 190, 191]. Вполне возможно, что клетки останавливаются тогда, когда совпадают их контактные участки. Отличие самосборки макромолекул в надмолекулярные структуры от самосборки многоклеточных построек, быть может, и состоит лишь в способе перемешивания — в первом случае мы имеем стохастическое тепловое движение, а во втором — целенаправленное или случайное активное перемещение клеток. А геометрия построек в обоих случаях определяется геометрией морфологических единиц — макромолекулярных комплексов или; целых клеток.
Так может быть все уже ясно? И можно допустить, что считывание генетических текстов — записей инструкций о морфологии многоклеточных организмов-—происходит посредством возникновения в нужной последовательности во времени клеток, необходимой формы с нужным числом контактов на поверхности? Но в ряде случаев клетки образуют структуры без непосредственных контактов и находятся друг около друга на расстоянии около 200 А. Промежуток же между клетками заполнен колла- геновыми волокнами или иными контактными веществами. Это- первое затруднение. Второе заключается в том, что при формообразовательных процессах в эмбриогенезе наблюдается перестройка структур; например, превращение бластулы в гаструлу происходит посредством изменения взаимного расположения уже образовавшихся ранее клеток, а именно, клетки некоторой зоны, (полюса) сферической бластулы вдавливаются, инвагинируются и образуют гаструлу. Трудно (трудно, но все же можно) представить себе детерминированное изменение морфологии клетки,, такое изменение числа контактных участков, ори котором из- сферы автоматически образовалась бы двуслойная гаструла.. Третье затруднение состоит в том, что одни и те же клетки в разных условиях дают разные структуры. При пересадке клеток эмбриона они образуют структуры, как правило, соответствующие тому месту, в которое их пересадили. Данная морфологическая структура индуцируется окружающими клетками и тканями. Явление индукции — предмет многих замечательных исследований в экспериментальной эмбриологии (см. [35, 291 ^ 295а, б]). Пожалуй, наиболее ярким, убедительным примером индуцированного морфогенеза является формирование оптической системы глаза. Глаз развивается в ходе эмбриогенеза из 'двух источников: передний мозг образует вырост—глазной бокал (превращающийся впоследствии в светочувствительную' ткань — сетчатку и в зрительный нерв), который индуцирует в ближайшей прикрывающей его эктодерме развитие оптической системы глаза — хрусталика и роговицы. Глазной бокал можно пересадить в разные места зародыша [182] и в этих местах в эктодерме будет индуцироваться развитие хрусталика и роговицы. Природа агентов, вызывающих индукцию данной структуры, не ясна. Вопрос этот очень сложен. Индуцирующим действием могут, например, обладать и заведомо неспецифические повреждающие воздействия, что когда-то поразило исследователей и заставило их надолго отказаться от поисков эндогенных специфических индукторов. С сожалением должен признаться, что и в данном случае наиболее вероятна доминирующая роль химии (конечно, биохимии), а не физики; определенные вещества, вероятно, индуцируют определенные структуры, а неспецифические повреждающие агенты способствуют выработке или освобождению таких веществ. Эти специфические индукторы, по-видимому, представляют собой факторы, снимающие репрессию определенного белкового синтеза.
Однако здесь уместно задать традиционный вопрос: Есть ли в жизни место физике? Иными словами нас интересует, существуют ли кардинальные биологические процессы, в которых фундаментальную роль играют именно биофизические механизмы. Существование химических индукторов не решает вопроса в пользу химии. Не вызывает сомнения, что регуляция считывания химического нуклеотидного текста наиболее успешно достигается посредством химических веществ. Но, учитывая сказанное
о возможном отсутствии прямых клеточных контактов и о возможности перестройки морфологии структур, можно допустить осуществление пространственных структур из клеток и посредством физических механизмов. В самом деле, наиболее естественно предположить, что пространство организуется под действием различных физических полей, т. е. совокупности градиентов каких-либо потенциалов — электрических, магнитных, механических, электромагнитных, акустических, гравитационных, а также (и тут нельзя избежать химии!) градиента концентрации каких- либо веществ. Замечательной особенностью процессов биологического морфогенеза является то, что эти поля (если они существуют, вернее, если они определяют процессы морфогенеза) вовсе не должны существовать сразу в завершенном виде в самом начале морфогенеза.
Морфогенные (биологические) поля в каждый данный момент должны характеризоваться относительно несложным распределением градиентов в пространстве. Никакого «образа» морфологии возникающего органа в морфогенном поле не должно быть. Последовательное во времени, соответствующее генетической программе изменение морфогенного поля может определить требуемое последовательное построение необходимой структуры из клеток. Таким образом, широко обсуждавшееся в свое время понятие биологического поля [18, 19, 76] соответствует интегралу по времени от действительного физического морфогенного поля , (если оно существует — мы еще не решили этот вопрос).
Итак, мы вероятно, должны заниматься не столько природой 'биологических полей, сколько исследовать возможную «сущность» виртуальных биологических полей, т. е. полей, существование которых дало бы эквивалентный эффект. Ввиду важности, этого вопроса задержимся на нем подробнее.
Представим себе, что последовательный во времени морфогенез осуществляется посредством соответствующего последовательного изменения поверхностных свойств клеток. Например,, допустим, что биполярные клетки, образующие нити и тяжи, становятся триполярными и вступают в контакты друг с другом в. трех точках. Возникшая в результате последовательного изменения клеток архитектура будет соответствовать некоторому виртуальному морфогенному полю. При таком подходе к проблеме биологического морфогенного поля основная задача заключается в выяснении механизма, определяющего относительно простую геометрию клетки. В основном речь идет о числе возможных точек контакта клетки с другими клетками. Вполне правильное- расположение этих контактов на поверхности клетки, вероятно, не обязательно — микроошибки в геометрии клеток сгладятся в многоклеточном образовании, как сглаживаются, становятся несущественными нерезкости изображения отдельных кадров при: просмотре кинофильма.
Чрезвычайно существенно также установить природу контактов— выяснить роль электростатических взаимодействий, ком- плементарности поверхностей (как при образовании комплементарное™ антиген-антитело), коллагенового клея, белково-поли- сахаридных контактных веществ [191]. Огромная важность этих проблем для теоретической и прикладной биологии очевидна. Геометрия, способность клеток образовывать контакты интересны не только в сеязи с морфогенезом при онтогенезе и регенерации, но и в особенности в связи с проблемой злокачественного роста.
Последовательное закономерное образование клеток разной формы, с разным числом по разному расположенных контактов может, следовательно, обеспечить закономерно сложную морфологию многоклеточных систем. Механизм образования клеток разной формы в соответствии с генотипическими предначертаниями вряд ли может основываться на простой самосборке из молекул с разным числом контактов.
Мне кажется весьма интересной модель механизма морфогенеза клеток, предложенная Л. А. Мартыновым [198, 423]. Он отметил аналогию между морфогенезом клеток и созданием формы надувных игрушек. Форма надувной, например, резиновой игрушки зависит от растяжимости отдельных участков ее оболочки, определяющейся толщиной и химическим составом самой оболочки. При надувании лучше растягивающиеся участки выпячиваются. «Надувание» клеток происходит в результате избыточного- внутреннего давления. Оно имеет осмотическую природу, создается работой мембранных противоградиентных насосов и поддерживается за счет свободной энергии гидролиза АТФ.
Мне кажется еще более наглядным аналогом процесса создания формы клетки работа стеклодува — получение предмета определенной формы путем воздействия на растяжимую разогретую стеклянную оболочку. Последовательно, целесообразно1 разогревая разные участки, стеклодув выдувает из стеклянного' пузыря причудливую елочную игрушку или лабораторный прибор. Последовательное соответствующее генетической программе образование разных компонент оболочки клеток, создающих то более, то менее растяжимые участки поверхности, затвердевание участков, растянутых ранее в результате биохимической доработки, наконец, воздействие постоянного тургора, внутреннего^ давления, поддерживаемого метаболическими процессами — таковы представления Мартынова о морфогенезе клетки. Мартынов отнюдь не ограничился созданием этой гипотезы, но выполнил также экспериментальное исследование по ее проверке. Он выбрал классический объект — уже упомянутую выше ацетабу- лярию (см. рис. 20, в). Сложная морфология зонта этой гигантской клетки диаметром около 5 см запрограммирована в единственном ее ядре, находящемся у основания корня — ризонда. Если отрезать зонт, то в случае неповрежденного ядра он возникает снова. Процесс регенерации зонта ацетабулярии и послужил Мартынову основанием для его модели. Он измерил параметры, определяющие механические свойства клеточной оболочки (модуль Юнга, внутриклеточное давление), а также исследовал оптическими методами расположение волокон целлюлозы в разных местах оболочки клетки на разных фазах морфогенеза.
Я не буду дальше излагать содержание и результаты опытов Мартынова — с ними можно познакомиться по его работам. Мне кажется, однако, что «стеклодувная модель» морфогенеза клетки хорошо соответствует современной молекулярной биологии — она заполняет разрыв между представлениями о самосборке надмолекулярных структур из разных макромолекул и представлением о пространственно упорядоченном расположении клеток в многоклеточной структуре. Так могут образоваться клетки с разным числом по разному расположенных контактов. Механизм морфогенеза, следующий из модели Мартынова, также не предполагает существования действительного сложного биологического морфогенного поля — в каждый момент времени происходит лишь несложный процесс, соответствующий составу клеточной оболочки.
Итак, остается вопрос — существуют ли помимо виртуальных действительные физические морфогенные поля? Он в значительной степени эквивалентен вопросу о существовании дальнодействия при взаимодействии клеток. Естественным способом осу-
ществления дальнодействия клеток в водной среде является их рлияние друг на друга посредством выделяемых в среду веществ.
Клетки могут получать сигналы о присутствии себе подобных посредством «запаха» или «вкуса». Это очень экономный способ•сигнализации: изготовленное сигнальное вещество может длительное время без дальнейших энергетических затрат со стороны клетки-изготовителя выполнять свои сигнальные функции, тогда как сигнализация посредством электромагнитных или акустических полей требует непрерывных трат для генерации сигналов. Конечно же информация о локализации источника сигналов, получаемая при помощи химических веществ, не очень совершенна — пресловутая организация пространства посредством градиентов концентрации данного вещества затрудняется беспорядочной тепловой диффузией. Однако на не очень больших расстояниях (порядка микрон), вероятно, и диффузия не слишком мешает нахождению клетками нужного направления движения и необходимых мест контакта. «Вкус и запах» естественные следствия классической биохимической эволюции. Восприятие присутствия и оценка концентрации определенных химических веществ в среде в принципе легко осуществляются нормальными биохимическими механизмами — по изменению скоростей ферментативных реакций в результате конкурентного или аллостерического взаимодействия этих веществ с макромолекулами ферментов. В ходе эволюции возникли высоко совершенные механизмы химической сигнальной регуляции взаимодействия клеток — системы гормональной регуляции, взаимодействие посредством антибиотиков, сигнализация запахами у животных.
Таким образом, и здесь кажется возможным обойтись чисто химическими, точнее биохимическими средствами. Однако можно поставить вопрос так. А нет ли каких-либо физических свойств, с неизбежностью проявляющихся при биохимических процессах, которые могли бы в ходе эволюции использоваться для дальнодействия при взаимодействии клеток в процессе морфогенеза? На вопрос в такой постановке можно ответить определенно: такие свойства есть — это электрические и акустические сигналы, неизбежно генерируемые клетками при протекании в них биохимических процессов. Генерация электрических сигналов — следствие изменения ионных градиентов и измененийконформаций макромолекул, несущих заряженные группы. Акустические сигналы могут возникать вследствие конформационных изменений макромолекул и их ансамблей. В каждом ферментативном акте макромолекулы ферментов в большинстве случаев претерпевают циклические изменения конформации, сопровождающиеся изменением упаковки в системе макромолекула —• вода, изменениями объема этой системы. Каждая макромолекула фермента в процессе катализа служит макрогенератором акустических сигналов. Взаимодействие одинаковых макромолекул, синхронизация их работы, может привести к генерации звуковой волны с частотой, определяемой числом оборотов фермента в каталитическом процессе. Таким образом, разнообразие и интенсивность работы ферментов в данный момент определяет частотный спектри интенсивность акустического поля, генерируемого клеткой. Любые генотипически или фенотипически обусловленные изменения внутриклеточных биохимических процессов должны в таком случае отражаться в изменении свойств акустического поля клетки. Возможно, не является чистой фантазией (если высказанные выше предположения верны) осуществление музыкального анализа, когда, «выслушивая клетку», удастся получать самые достоверные и оперативные сведения о ее внутренней жизни.
Из общих соображений представляется вероятным, что акустическое поле вполне пригодно для морфогенеза. Всякое строительство связано с толчками, передвижениями и прочими механическими воздействиями — что же может быть лучше толчков и механических давлений градиентов, создаваемых клетками, для строительства многоклеточных конструкций?
Помимо чисто акустических сил, синхронизированные в макрообъеме конформационные колебания ферментов и обусловленные ими пульсации мембран клеток, могут иметь существенное морфогенное значение в силу гидродинамических следствий таких колебаний. Пульсирующие клетки, разделенные слоем подвижной воды, будут притягиваться или отталкиваться в зависимости от частоты (и степени синфазности) пульсаций. На возможную роль гидродинамических сил, возникающих при пульсации клеток, обращает внимание Уоддингтон в книге «Морфогенез и генетика» [2956]. Я просто процитирую часть его текста, посвященную этому вопросу ...«Быть может, наиболее простой случай возникновения таких сил — это так называемый эффект Гийо — Бьеркнеса. Этот эффект, известный уже более ста лет, заключается в том, что две пульсирующие сферы, погруженные в жидкость, притягиваются, если их пульсации совпадают по фазе или отталкиваются, если разность фаз равна 180°. Этот эффект обусловлен действием гидродинамических сил, возникающих вследствие того, что давление в установившемся течении' жидкости тем меньше, чем больше скорость течения (теорема Бернулли). Фаберже попытался объяснить действием этих сил спаривание гомологичных хромосом в профазе мейоза. Он основывал свои рассуждения на простейшей модели двух сферических тел, пульсирующих в радиальном направлении по всей поверхности... В действительности, однако, мы имеем дело с гораздо более сложной системой. В ней имеется много пульсирующих тел разных размеров и с различными (по крайней мере вначале) периодами пульсации. По-видимому, эти колебания взаимодействуют не только вследствие резонанса, возникающего благодаря среде, в которую погружены эти тела; при этом имеют значения изменения электрического поля вокруг частиц, обусловленные изменениями конфигурации молекул, вызывающими пульсацию. Кроме того, в больших макромолекулах — и: даже в большей степени в частицах — пульсация, по-видимому,.
происходит несимметрично; вероятно, имеется несколько мест пульсации, расположенных в определенных участках на поверхности органелл» [2956, с. 162—163].
Нам сейчас не очень важно, действует ли акустическое поле, возникающее при конформационных колебаниях макромолекул, в результате гидродинамических эффектов или непосредственно. В любом случае кажется вероятным, что такое поле может определять пространственные взаимоотношения клеток (и самих макромолекул), быть существенной частью механизма биологического морфогенеза.
Взаимодействие клеток может обусловить сложные пространственные эффекты в результате интерференции генерируемых ими акустических сигналов. Так, две одинаковые, одинаково звучащие клетки, расположенные на близком расстоянии друг от друга, создадут в результате интерференции «тихую плоскость»— в этой плоскости спокойно, без толчков смогут находиться другие клетки. Четыре «летки-генератора акустических полей могут образовать «тихий луч» и т. д. Различие частот, генерируемых разными клетками, изменение этих частот по.мере морфогенеза способны обеспечить весьма сложную и совершенную пространственную ориентацию клеток (дополняемую еще и специфическими контактами).
Попробуем оценить теперь, в какой мере реальные для биохимических систем клетки частоты возможных звуковых полей могут обеспечить геометрические задачи морфогенеза. Ясно, что речь может идти лишь о волнах, длины которых сравнимы с размерами клеток, т. е. о звуковых волнах длиной порядка 1 — 100 мк\ при нормальной скорости звука их частоты порядка 107—109 гц. Однако в желеобразных протоплазматических структурах распространяются преимущественно лишь поперечные сдвиговые волны. Их скорость, как показал А. П. Сарвазян с сотрудниками [253] порядка 1 м!сек. Соответственно, необходимые длины волн могут быть получены при частотах — порядка 104—-10е гц. Этот диапазон частот вполне биохимически реален — на таких частотах работают основные ферменты. Подробнее мы рассмотрим эти вопросы в 9-й рл. Сейчас же достаточно отметить, что числа оборотов большинства ферментов соответствуют слышимому диапазону частот (см. табл. 2 в гл. 4). Это наводит на приятные размышления о природе слуха, происхождении музыкального восприятия и о многом другом, что принадлежит уже области биохимической эстетики, а не биохимии морфогенеза.
И все же, вероятно, наиболее трудной задачей является объяснение механизмов морфогенеза не многоклеточных организмов, в сложно построенных одноклеточных (см. рис. 15, 20). Поэтому наиболее актуальным представляется здесь исследование механизмов морфогенеза клеток.
Может показаться, что морфологический прогресс в процессе эволюции вовсе не обязательно связан с ростом кинетическогосовершенства. Действительно, образование опорного скелета обусловлено приспособлением к существованию в условиях земного притяжения, сложнейшие морфофизиологические корреляции цветков и насекомых — совершенствованием в ходе сопряженного естественного отбора процессов опыления цветков и питания насекомых. Все это верно, однако, конечным результатом всех этих приспособлений, конечным решающим критерием естественного отбора во всех случаях служит итоговая интенсивность преобразования веществ в вещества данного вида, т. е. биологическое (кинетическое) совершенство.
Цветковые растения — пример морфологического прогресса. По-видимому, предела увеличения кинетического совершенства на пути «чистого» морфологического прогресса достигли на Земле покрытосеменные (цветковые) растения. Поэтому (следуя принятой манере изложения) после анализа общих возможных механизмов морфогенеза рассмотрим некоторые существенные в эволюции особенности цветковых растений.
Цветковые — господствующая на Земле группа растений — отличаются многими замечательными чертами от всех остальных растений. Причины внезапного бурного заполнения всех возможных ареалов Земли цветковыми растениями служили предметом часто весьма эмоциональных и ярких построений ботаников. (Как и в других аналогичных случаях, я предпочитаю предоставить слово специалистам). Н. В. Первухина в книге «Проблемы морфологии и биологии цветка» [234] пишет, что появление покрытосеменных, или цветковых, растений представляет собой один из самых значительных этапов эволюции органической жизни на Земле. М. И. Голенкин назвал свою книгу, посвященную цветковым растениям, «Победители в борьбе за существование» [70]. В самом деле, покрытосеменные являются господствующей группой растений на нашей планете, они составляют главную массу растительного вещества (высших растений). В настоящее время, как указывает Первухина, цветковые занимают то место растительных космических деятелей, которое в прежние эры было занято ныне исчезнувшими папоротникообразными, образователями толщ каменного угля. Современный животный мир, особенно высшие позвоночные (млекопитающие, птицы) и насекомые, всецело зависят от покрытосеменных. Человек, по образному выражению Голенкина, является порождением покрытосеменных. Победа цветковых была переломным этапом, революцией в судьбах всего живого населения Земли. Поэтому середина мелового периода — время массового появления покрытосеменных на Земле — начало новой эры в жизни нашей планеты. Замечательно, что эта новая эра наступает внезапно. Сьюорд [280] говорил о победе покрытосеменных на Земле как о «головокружительно блестящей карьере», сравнимой со «всемирным взрывом». Еще Дарвин отмечал, что вопрос о происхождении покрытосеменных окружен «ужасной тайной». В самом
деле, что обусловило столь явное биологическое совершенство цветковых растений? Вероятно целый комплекс достоинств. Покрытосеменные отличаются от других растений чрезвычайно высокими темпами роста и развития. Голенкин считал наиболее характерной жизненной особенностью покрытосеменных их способность выдерживать максимальные дозы яркого солнечного света. В. Н. Любименко по приспособленности к световому образу жизни покрытосеменных считал покрытосеменных «венцом эволюции». Из приведенного «конспекта» текста Н. В. Первухиной видно, что главным отличием цветковых растений от всех остальных является их наиболее быстрый рост и развитие, обусловленные наиболее интенсивным фотосинтезом. Это позволяет цветковым растениям быстро совершать жизненный цикл (накапливая большую биомассу!), приспосабливаться к жизни & местностях с кратковременными периодами, благоприятными для жизнедеятельности (краткое лето в околоарктических широтах, краткие периоды достаточной влажности в пустынях и степях). Кратковременность, односезонность жизненного цикла позволяет цветковым растениям (первично древовидным) существовать в виде биологически весьма совершенных трав [283, 284]. Все в целом приводит к резкому ускорению эволюционного развития цветковых растений.
Теперь уместно попытаться ответить на вопрос: что является ведущим звеном в процессе биологического совершенствования цветковых растений?
Узловым моментом здесь, вероятно, оказывается скорость движения веществ в проводящих путях цветковых растений, М. И. Голенкин [70] характеризует систему проводящих пучков покрытосеменных как доведенную до последней степени совершенства, что обеспечивает одновременно быстрый приток минеральных веществ и воды в столь же быстрый отток образовавшихся в листьях веществ1. А. Л. Тахтаджян полагает, что способность покрытосеменных к значительному увеличению количества синтезируемого в процессе фотосинтеза органического вещества связана с возникновением высокосовершенных проводящих сосудов. По существу речь здесь идет о следствиях морфологического прогресса. У большинства предшественников цветковых растений движение воды (водных растворов) сопряжено с неоднократным прохождением ее через одревесневшие стенки трахеид (например, у голосеменных). Ясно, что такое движение не может быть достаточно интенсивным. У цветковых растений трахеиды становятся члениками сосудов — цилиндрическими сегментами водопроводной трубы, иногда непрерывно идущей от корней к листьям [357]. Однако известно, что водопроводные сосуды многократно и независимо возникали в ходе эволюции у растений разных групп — у плауновидных, папоротников, голо-
семенных, покрытосеменных (см. ниже). Таким образом дело не в том, что лишь покрытосеменные растения «научились» изготовлять сосуды без перегородок. Эта задача решалась и раньше в эволюции менее прогрессивных групп. Зато, вероятно, только цветковые растения смогли извлечь из резкого ускорения потока воды максимальную пользу, что обусловливается особым совершенством устьичного аппарата, биохимии фотосинтеза и, наконец, самым явным преимуществом покрытосеменных, а именно, образованием настоящих семян, приспособленных для сохранения и распространения жизни в весьма суровых и разнообразных условия*. На фоне всего этого комплекса достоинств покрытосеменных возникновение непрерывных водопроводных сосудов оказалось истинным ароморфозом, резко увеличившим биологическое совершенство этой группы и обеспечившим способность покрытосеменных быстро заполнить занятые ранее другими экологические ниши.
Итак, исчезновение перегородок в водопроводных сосудах именно у покрытосеменных представляется мне преодолением последнего узкого места, что позволило этим растениям совершить головокружительную карьеру. Теперь вода быстро движется непрерывным тяжем, нитью за счет корневого давления и капиллярных сил, сил поверхностного натяжения.
Конечно же, и трахеиды в ходе эволюции достигли предельно возможного для них совершенства. В их одревесневших стенках возникли относительно тонкостенные поры. В одревесневших мембранах этих пор у голосеменных возникли микроскопические •отверстия диаметром порядка 10-4 см.
Попробуем дать количественную оценку возможных скоростей .движения воды по трахеям — водопроводным сосудам у цветковых растений и по системе трахеид (например, у хвойных).
Трахея цветковых растений представляет собой капилляр с радиусом порядка 5-10-3—5-10~2 см и длиной порядка 1 —102 см. Вода движется непрерывным потоком по трахее. Трахеиды-- мертвые одревесневшие .клетки диаметром порядка ЫО-2 ем и длиной порядка 10-1 см. Вода движется по системе трахеид, проходя сквозь поры толщиной 10-4—10“3 см (отверстия в них) от одной трахеиды и другой.
Оценим соотношения скоростей потоков воды по трахеям и трахеидам.
В идеализированной схеме система трахеид аналогична трахее ■с пористой перегородкой, толщина которой определяется числом трахеид, приходящихся на единицу длины трахеи. Примем, что на каждый см. длины приходится 10 пористых мембран (поскольку размер трахеиды 10-1 см). Тогда толщина /2 суммарной пористой перегородки будет равна 4=Ю /Д где — общая длина системы трахеид (равная длине сравниваемой с нею трахеи), d — толщина единичной перегородки (в см), соответствующая диапазону 10~4—10~3 см.
Движение жидкости в трубах описывается формулой Пуа- зейля.
Q = АРП^4 18(х ’
где Q —расход жидкости в см3/сек, Ар —перепад давления (дн/см2), [х — вязкость (пз), / — длина и Я —радиус капилляра. Допустим теперь, что суммарная площадь отверстий в перегородках трахеид равна площади поперечного сечения трахеи, т. е. лпЯ22 = п#Д где 7?, —радиус трахеи, R2 — радиус отверстия в поре трахеиды, п — число отверстий в порах трахеиды. Пусть в обеих системах будут одинаковы перепады давления. Тогда отношение а объемной скорости потока воды в трахее по сравнению с системой трахеид определится формулой
_ Qi _ l^_ _
Q* hRl hRl
Ниже приведены численные оценки по этой формуле для принятых выше геометрических характеристик. Для самой узкой трахеи ^, = 5-10‘3 я самой тонкой перегородкой трахеиды d= 10~4 см а = 2,5.
R1 МО'3 МО-3 5-10-3 5-10-3 1-10-2 1 10- 2 10-! 1C-1 d МО"4 МО'3 МО'4 1 -10“3 1 • 10~4 МО'3 МО-4 МО'1 а 0,1 1 2,5 25 10 Ю3 3 Ю4 10'
Как видно, трахеиды с очень тонкими перегородками не уступают, а даже превосходят очень узкие трахеи (^ = 10“3 см) по’ скорости транспорта воды. Это, конечно, не следует понимать буквально, в этом случае сопротивление движению жидкости по длине капилляра внутри трахеиды оказывается сравнимым с сопротивлением в перегородке. При радиусах трахей больших МО-3 см их преимущество перед трахеидами становится очевидным. Для трахей с Rt порядка 10“г—10-1 см( как у ясеня, дуба и других лиственных деревьев) эти преимущества становятся весьма значительными — в 102—104 раз.
Из этих оценок ясно направление эволюционного совершенствования проводящих путей растений: утоньшение перегородок, пористых мембран трахеид и увеличение радиуса трахей.
Мы приняли, что суммарная площадь отверстий в перегородках трахеид равна площади поперечного сечения трахеи. На самом деле, эта суммарная площадь, вероятно, существенно меньше. Таким образом наши оценки можно считать достаточно осторожными.
Естественно, попытаться найти им фактическое подтверждение. Однако получение данных по объемной скорости потока воды — в см3/сек в разных растениях очень сложно. Гораздо более реально измерение средней линейной скорости перемещения воды в стеблях. Попытаемся оценить величины этой скорости для трахей и трахеид в соответствии с уже принятыми допущениями. Средняя линейная скорость течения жидкостей по капилляру задается формулами:
у = -9— = АР^2
nR2 18[х
Для численных оценок V по этой формуле необходимо знание величин Др и [1. Перепад давления на метр длины стебля Ар можно принять равным порядка 1 атм (Ю4 дн/см2 [155]), для воды 10-2 пз. Тогда для средней (линейной) скорости передвижения воды в трахеях (Ri= 10_3-М0-1 см) получим величины, сведенные ниже.
R, см
ю-4
10-3
10-2
10-1
V, см/сек
ю-3
10-!
10
103
V, м/час
0,0036
0,36
360
36-103
Посмотрим, чему равна средняя скорость движения воды в системе трахеид при толщине перегородок d и длине одной трахеиды 10-1 см. Можно принять, что в системе трахеид основной перепад давления приходится на отверстия в порах. На 1 м длины стебля приходится 103 перегородок трахеид общей длины Ю3 d Отсюда средняя скорость движения воды по трахеидам будет равна (^2=Ю-4см)
= — Rl см/сек.
d,
см
10~4
10~3
10~2
V,
см/сек
1
10-[12]
10~2
V,
м/час
30
3,0
0,36
В этой оценке мы полагали отверстия в порах — «узким местом», определяющим суммарную скорость потока. На самом деле для узких трахеид с радиусом порядка 10_3 см существенно замедление потока и при движении до и после пористых перегородок. Из сравнения величин, приведенных в табл. 13 и 14. видно, что линейная скорость движения воды в трахеях существенно превышает скорость движения воды в системе трахеид при радиусе поперечного сечения трахеи порядка 10-2 см.
Если наши оценки правильны, то предельно возможная скорость движения воды по трахеидам приближается к нижней границе скорости движения воды по трахеям. Мы вправе считать, что эти оценки свидетельствуют в пользу правильности утверждения о возможности резкого ускорения движения воды при переходе от трахеид голосеменных к трахеям цветковых растений.
Теперь пришло время попытаться выяснить, каковы действительные скорости движения воды в стволах (стеблях) голосеменных и покрытосеменных растений. Удивительно, но вопрос о
действительных скоростях движения воды в стволах деревьев все еще нельзя считать выясненным *.
Скорость движения воды в ксилеме измеряли последние 50— 100 лет, применяя самые разные методы, однако, среди них нельзя назвать ни одного вполне удовлетворительного. Может быть поэтому Гейне [398] в своем обзоре, опубликованном в 1971 г., в основном опирается на измерения 1905—1930 гг. Не привело к значительному прогрессу и применение радиоактивных индикаторов [46]. В книге Крамера и Козловского [155] приводятся, в частности, данные Губера и Шмидта о максимальных скоростях движения воды в ксилеме покрытосеменных и хвойных деревьев. Эти авторы измеряли движение воды термоэлектрическим методом — по скорости распространения по стволу нагретой в ксилеме воды — и установили, что максимальные скорости движения воды у лиственных пород варьируют от 45 до 3 м/час (дуб — 43,6, ясень — 25,7, вяз — 6, орех — 3,7 м/час), а у хвойных составляют менее 0,5 м/час.
Приведенные значения порядка 1 см/сек для лиственных пород близки к полученным оценкам, а для хвойных (0,01 см/'сек) на 1—2 порядка меньше моих оценок. Причины этого расхождения скорее всего в завышении суммарной площади отверстий в порах трахеид в приведенных расчетах. Однако то обстоятельство, что скорость движения воды у лиственных пород примерно на два порядка выше, чем у хвойных, по-видимому, подтверждает главную мысль — основой кинетического совершенства цветковых растений является морфология водопроводной системы, обеспечивающая высокую скорость движения воды в растении.
Глава 9
ПЕРЕМЕЩЕНИЯ В ПРОСТРАНСТВЕ. МЕХАНО-ХИМИЧЕСКИЕ ПРЕОБРАЗОВАНИЯ ЭНЕРГИИ. ПРОИСХОЖДЕНИЕ МНОГОКЛЕТОЧНОСТИ
П реодоление диффузионное о барьера посредством перемещений в пространстве.
Формирование специальных аппаратов движения на основе: а) изменений поверхностного натяжения, б) ферментативных механо-химических преобразователей энергии. Ограничение предельных размеров и частоты биений макромолекулярных комплексов в двигательных аппаратах диффузией и концентрацией субстратов.
Переход от конформационных циклов к скольжению. Локальные изменения pH в качестве первичного механизма мехам-химических преобразований. Мчогоклеточность как эволюционный результат специализации и интенсификации функций.
Многоклеточность — следствие перехода к движению посредством миофибрилл.
Морфологический прогресс — относительно малоэффективный способ преодоления диффузионного барьера. Второй — высокоэффективный способ — активное перемещение в пространстве организмов или среды к организмам. Первым шагом на этом пути служит беспорядочное движение — простое перемешивание. Ненаправленное перемешивание приводит к увеличению сечения взаимодействия реагентов, к росту вероятности столкновения молекул, и, следовательно, к возрастанию скорости реакций. Эволюционный потенциал в этом направлении эволюции определяется возможностью векторизации перемещений в пространстве — выработки механизмов все более целеустремленного, все менее беспорядочного движения. «Геометрический образ» эволюционного процесса и в данном случае (как и в случае катализа или матричного воспроизведения) — переход от сферически симметричных траекторий к уникальной. Предел эволюции здесь физически определен весьма жестко. Скорость биохимических реакций лимитируется скоростью потока реагентов и продуктов, равной скорости свободного пробега молекул в среде, т. е. скорости звука. Эволюционный потенциал этого этапа соответствует возможному изменению скорости заполнения пространства веществом данного вида от скорости диффузии (10~2—10_3 см/сек) до скорости звука (105 см/сек), что составляет 7—8 порядков.
Вопрос о максимальной скорости биосинтеза живого вещества рассматривал много лет назад В. И. Вернадский [49]. Он оценивал скорость заселения нашей планеты разными организмамипри условии их беспрепятственного размножения. Максимальная скорость «передачи жизни» была найдена Вернадским для бактерий и оказалась равной 330 м/сек, т. е. скорости звука в воздухе. С такой средней скоростью распространялась бы по поверхности Земли пленка бактерий. Заселение Земли слонами происходило бы гораздо медленнее — со скоростью порядка 0,1 см/сек. Однако, если учесть различия в массах бактерий и слонов, то станет ясно, что по скорости прироста биомассы слоны не уступают бактериям. Замечательно, что Вернадский связывал скорость звука со скоростью биохимических процессов, а именно, с предельной скоростью дыхания. Он писал: «Получив для самых быстро размножающихся бактерий скорость размножения, равную по порядку скорости распространения звука в воздухе, я не счел это случайным совпадением, но принял за реальный факт тесно связанный с дыханием бактерий» [49, с. 313].
Движение, перемещение в пространстве посредством роста и размножения лишь конечный итог, результат активных перемещений организмов для обеспечения биосинтезов необходимыми веществами и освобождения от конечных продуктов метаболизма. Однако в некоторых случаях и движение вследствие роста и размножения является способом увеличения вероятности взаимодействия организма к источников пиши.
Попробуем придумать возможные механизмы перемещения в пространстве, исходя из эволюционных достижений предыдущих этапов. В результате этих достижений образовались клетки, содержащие необходимый комплект матричных макромолекул, белков-ферментов, липопротеидных структур — поверхностных мембран и внутриклеточных оргаиелл.
ПЕРЕМЕЩЕНИЕ В ПРОСТРАНСТВЕ ПОСРЕДСТВОМ ИЗМЕНЕНИИ ПОВЕРХНОСТНОГО НАТЯЖЕНИЯ
Простейшим механизмом, обеспечивающим перемещение в пространстве нашей схематизированной клетки, представляется изменение поверхностного натяжения на границе раздела: наружная мембрана — внешняя среда (вода). Причиной увеличения или уменьшения поверхностного натяжения может быть изменение соотношения гидрофобных и гидрофильных групп в липопротеидных комплексах, образующих мембрану. Если расстояние, на которое должны переместиться клетка, превышает ее линейные размеры, аппарат, обеспечивающий движение, должен работать периодически. Поэтому п изменения поверхностного натяжения должны быть периодическими. Периодические, обратимые изменения поверхностного натяжения в разных местах наружной мембраны приведут к беспорядочному, разнонаправленному «перетеканию» клетки с места на место — образованию псевдоподий и (к) амебоидному движению. Если такие изменения поверхностного натяжения будут происходить лишь в некоторыхместах поверхностной мембраны, клетка будет получать импульс и перемещаться в каком-либо определенном направлении, совершать более или менее «целесообразные» амебоидные движения.
Наиболее вероятным представляется изменение поверхностного натяжения наружной мембраны в результате выделения из клетки низкомолекулярных веществ (лигандов), связывание которых с липопротеидными комплексами изменяет в них гидрофобно-гидрофильные соотношения. Такими лигандами могут быть спирты, жирные кислоты или двухвалентные катионы, например ионы кальция.
В такой модели клетка уподобляется капле жидкости, поверхностное натяжение которой изменяется в результате реакции поверхностных макромолекул с веществом, порциями поступающим изнутри капли (клетки). Форма капли резко изменяется, и сама капля перемещается за счет сообщаемого ей этим изменением импульса. Вещество, вступившее в реакцию с поверхностными макромолекулами, расщепляется в ходе ферментативного процесса, катализируемого, например, самими этими макромолекулами, и форма капли возвращается к исходной. Многократное, периодическое изменение формы клетки может привести к непрерывному перемещению организма в пространстве, причем его направление определяется локализацией участков поверхности, изменяющих свои свойства. Прежде чем анализировать эту модель, следует отметить, что она была создана около 100 лет назад и была очень популярна в начале нашего века.
В 1888 г. Квинке предложил теорию протоплазматического (амебоидного) движения, основанную на аналогии с процессом растекания капель масла по поверхности воды. Очень хорошее изложение взглядов Квинке и всей проблемы в целом было осуществлено замечательным физиологом и биологом Ж. Лебом в его классической книге «Динамика живого вещества» [167]. Для иллюстрации такого способа движения предлагалось несколько моделей. Одну из них и рассматривает Леб. В его модели к кап^ ле оливкового масла добавлялось такое количество хлороформа, чтобы удельный вес смеси сравнялся с удельным весом раствора соды (0,5—2%), в который и погружали каплю смеси. В оливковом масле всегда присутствуют значительные количества жирных кислот, и на поверхности капли они образуют мыло (т. е. соли с ионами натрия окружающего раствора). Поверхностное натяжение в месте образования мыла резко падает, и из капли вырастает «псевдоподия», в которую устремляется содержимое капли. При этом площадь взаимодействующей со средой поверхности растет, образуются новые порции мыла — капля движется.
Таким образом, наша исходная модель отнюдь не нова. Впрочем, именно наглядность, явная и грубая упрощенность модели Квинке вызывала в свое время вполне справедливые упреки в «механицизме».
Аппараты биологического перемещения в пространстве, реальные механизмы движения протоплазмы или амебоидного движения бесспорно специфичны и сложны, поскольку они возникли в результате сотен миллионов лет эволюции. Однако исходные для эволюции физические механизмы, вероятно, вполне соответствуют принципам, механизмам движения неспециализированных макромолекулярных комплексов. И сейчас, с высоты уже имеющихся знаний о специфичных и сложных аппаратах биологического перемещения в пространстве мы можем попытаться найти эти исходные принципы и посмотреть, куда должна привести биологические системы с такими исходными данными последующая эволюция. (В предложенной Л. X. Эйдусом [351] теории мышечного сокращения изменение поверхностного натяжения, капиллярных сил рассматриваются в качестве основного механизма).
Кинетически механизм перемещения в пространстве, основанный на изменении поверхностного натяжения наружной мембраны, может быть довольно совершенным. Можно представить себе быстрое выделение необходимых лигандов вслед за «возбуждающим» сигналом через мембрану, а также довольно быстрое обратное всасывание или ферментативное разрушение этих веществ, что необходимо для периодичности функционирования механизма движения.
Однако термодинамически этот механизм оказывается мало совершенным. Выделяемые вещества будут вымываться в среду, создание нужной их концентрации достигается ценой расточительных затрат.
Работа по перемещению в пространстве совершается в данном случае за счет перепада концентраций лиганда при его связывании липопротеидными структурами (A^~RT 1п(с2/с1)). Этот перепад концентраций не может быть очень большим — при возбуждении лиганд переходит из связанного состояния внутри клетки в свободное. Такой переход может осуществляться при резком изменении сродства лиганда к какому-либо внутриклеточному макромолекулярному комплексообразователю. Трудно представить себе наличие в клетке большой концентрации этого комплексообразователя — без чего нельзя обеспечить высокую концентрацию этого поверхностно активного вещества — лиганда с наружной стороны липопротеидной мембраны. Таким образом, перемещения в пространстве посредством изменения поверхностного натяжения липопротеидных структур наружной мембраны клетки несовершенны по двум причинам: 1) из-за большого объема наружной среды, что сопряжено с расходом больших количеств вещества для обеспечения нужной концентрации лиганда, и 2) из-за низкой удельной энергопродукции — малой величины механической работы, производимой на единицу массы лиганда, связываемого «рабочим телом» наружной мембраны. Последнее обусловлено как малыми реальными пере^падами концентрации лиганда в рабочем цикле, так и большим «паразитным» объемом системы.
Эти причины несовершенства, «узкие места» первоначальных, неспециализированных аппаратов перемещения в пространстве — физико-химические по существу — и становятся критериями естественного отбора, направляющие эволюцию, т. е. являются очередными физико-химическими факторами биологической эволюции.
Предельно совершенным решением эволюционной задачи создания молекулярного механизма перемещения в пространстве будет предельно возможное уменьшение паразитного объема, переход к созданию локальной концентрации лиганда в активном центре макромолекулы, изменяющей свою конформацию при связывании лиганда. Для обеспечения не просто -периодичности, а выполнения полезной работы связываемый лиганд должен расщепляться на одной из стадий циклического процесса в экзэрго- нической реакции.
Всем этим условиям удовлетворяет ферментативный механо- химический преобразователь энергии.
ФЕРМЕНТАТИВНЫЙ МЕХАНО-ХИМИЧЕСКИИ ПРЕОБРАЗОВАТЕЛЬ ЭНЕРГИИ
Таким образом, предельно совершенным молекулярным аппаратом перемещения в пространстве будет предельно совершенный фермент, максимально возможно изменяющий свою третичную (конформацию) или четвертичную (конфигурацию комплекса макромолекул) структуру сопряженно с превращением субстрата в продукт [58].
Мы видели в главе 4, что изменение конформации — необходимое условие совершенного ферментативного катализа. Однако вообще в ферментативном катализе изменение конформации — лишь необходимое условие, средство проведения реакции по наиболее выгодному маршруту. В аппаратах перемещения циклические изменения конформации в некотором смысле — «основной продукт» реакции. Поэтому в ходе эволюции должны возникнуть специализированные на механо-химических преобразованиях ферменты.
Рассмотрим общие принципы работы идеальных ферментативных механо-хим'ических преобразователей, следуя в основном работам А. Е. Букатиной и сотр. [40, 41, 42, 87, 214, 430, 431].
Представим себе макромолекулярную «пружину» с двумя одноименными (пусть это будут ®) электрическими зарядами на концах (рис. 21, Е). Из-за отталкивания одноименных зарядов пружина находится в максимально растянутом состоянии. При появлении в среде вещества S-молекулы с двумя отрицательно заряженными концевыми (например, карбоксильными) группами пружина сжимается (рис. 21, Е) и производит меха-


РИС. 21. Схема работы ферментативного механо-химического преобразователи энергии
Е — свободный фермент; 5 — субстрат; ES — фермент-субстратный комплекс; ЕР — фер- мент-продуктиый комплекс; /g —длина £: /gs “ Длина £ в ЈS комплексе /Ј>/eS
РИС. 22. Изменение потенциального профиля при работе идеального ферментативного механо-химического преобразователя энергии
/ — стадия образования £5-комплскса п >корочения Е. 1-й барьер понижен. II стадия — £5-комплекс превращается в £Р-комплекс. происходит каталитическое превращение субстрата в продукт. 1-й барьер повышен, 2-й — понижен; III стадия — £Р-комплекс расщепляется. Оба барьера повышены
ническую работу. При завершении сжатия макромолекулярной пружины на макромолекуле образуется каталитический центр, в котором осуществляется реакция (в нашем примере) «дека- питации», S —обе отрицательно заряженные группы уничтожаются в реакции декарбоксилирования (рис. 21, ЕР) пружина распрямляется. Такая молекулярная пружина будет циклически совершать механическую работу за счет энергии, выделяющейся при расщеплении субстрата S. Замечательно, что сама по себе реакция разрушения субстрата может и не быть экзэргонической. Энергия может выделяться и на другой стадии цикла, например, при связывании субстрата. Поэтому определяющим условием работы такого механо-химического преобразователя будет целесообразное изменение кинетических потенциальных барьеров — целесообразное изменение каталитических свойств в определенных фазах цикла. Субстрат S не должен расшепляться несократившимся ферментом, что обеспечивается большим потенциальным барьером, препятствующим расщеплению. Этот барьер должен резко понижаться в конце процесса укорочения «пружины» и вновь резко возрастать, как только субстрат превратится в продукт. Иначе будет идти обратный процесс — ресинтез субстрата из продукта за счет механического напряжения макромолекулы под влиянием нагрузки. Новая молекула субстрата не должна соединяться с ферментом до его полного распрямления — иначе амплитуда перемещения (Д/) уменьшится, и соответственно уменьшится выполняемая работа. Это может быть обеспечено целесообразным изменением потенциального барьера, препятствующего образованию комплекса (рис. 22). Понижение барьеров аналогично открыванию клапанов в паровой машине или двигателе внутреннего сгорания. Открылся первый клапан, т. е. понизился первый барьер (в результате растяжения пружины при отталкивании одноименных зарядов в нашей модели) — субстрат получил доступ к центру связывания фермента, пружина в £5-комплексе сократилась, совершилась механическая работа одновременно повысился первый барьер, т. е. закрылся первый клапан, образовался каталитический центр, резко понизился второй барьер, т. е. открылся второй клапан, субстрат превратился в продукт, пружина (каждая макромолекула в отдельности) распрямилась, второй барьер повысился, второй клапан закрылся, но снова открылся первый, и т. д. Так работает механо-хнмический ферментативный «трехтактный» преобразователь энергии (рис. 23).

t
Наш трехтактный ферментативный механо-химический двигатель построен на основании общих соображений, теоретического анализа, проведенного в работах [41, 42]. Реальные механизмы биологической подвижности не обязательно должны соответствовать этой абстрактной схеме. Разным видам биологической подвижности могут отвечать различные модификации предельно совершенного образца. Однако во всех случаях речь идет о ме- хано-химических, вернее, химико-механических преобразованиях энергии. В связи с этим необходимо подчеркнуть, что сам принцип механо-химических ферментативных преобразований был сформулирован, н его первые экспериментальные подтверждения были получены В. А. Энгельгардтом и М. Н. Любимовой в 1937— 1946 гг. [184, 354]. Непреходящая ценность работ М. Н. Любимовой и В. А. Энгельгардта, открывших АТФ-азную активность миозина (актомиозпча пи современной терминологии), состоит
РИС. 23. Характеристики идеального «ферментативного трехтактного двигателя»
Изменения: а —длины lЈ макромолекулы фермента; б —высоты потенциального барьера, определяющего скорость связывания субстрата 8j и в — высоты потенциального барьера, определяющего скорость превращения субстрата в продукт 8 ij. По оси абсцисс — времяименно в обнаружении экспериментальных оснований для формулирования общего принципа. Этот механо-химический принцип должен по праву называться принципом Энгельгардта. По существу, ставшее столь популярным представление о конформа- ционных движениях макромолекул фермента в связи с его функцией есть развитие принципа Энгельгардта. Естественно, что наиболее явственно он выступает в механизмах биологической подвижности.
В последовавшие за работами Энгельгардта годы принцип механо-химических преобразований при ферментативном катализе получил широкое признание и развитие [175, 176, 409].
Особое, весьма большое значение для развития этой области имели работы А. Качальского и В. Куна и их учеников и сотрудников [409]. Созданные ими на основе биополимерных материалов двигатели осуществляли механо-химические преобразования энергии. Строгий анализ термодинамических и кинетических закономерностей при механо-химических преобразованиях был осуществлен А. Е. Букатиной и В. Н. Морозовым [41, 42], как это отмечено выше.
СИНХРОНИЗАЦИЯ КОНФОРМАЦИОННЫХ ИЗМЕНЕНИЙ МАКРОМОЛЕКУЛ ФЕРМЕНТОВ
Пока мы рассматривали механо-химический преобразователь энергии, состоящий из одной свободно плавающей макромолекулы фермента. При сколь угодно эффективном преобразовании химической энергии в механическую такая макромолекула мало пригодна для перемещений в пространстве макрообъектов. Ее собственные перемещения в пространстве будут осуществляться лишь реактивно, посредством импульсов отдачи. Действительно, в растворе таких макромолекул, движущихся несинхронно и в случайных направлениях, будет происходить лишь некоторое перемешивание, эквивалентное возрастанию амплитуд «броуни- рования» (по терминологии Н. И. Кобозева [136]). Таким образом, «эволюционный потенциал» — возможность эволюционного совершенствования — определяется переходом от беспорядочно движущихся макромолекул ферментов, осуществляющих механо- химические преобразования к предельно векторизованным перемещениям макроскопических многомолекулярных ансамблей таких макромолекул.
Простейшим и, вероятно, исходным способом создания таких многомолекулярных ансамблей является синхронизация кон- формационных циклов свободно растворенных макромолекул ферментов.
Реальна ли синхронизация конформационных движений отдельных макромолекул? Мне кажется, что вполне [335—338, 342, 458]. В са,мом деле, представим себе обстановку в протоплазме. Макромолекулы разных ферментов, отделенные друг отдруга большим или меньшим слоем воды, грызут или сшивают свои субстраты, претерпевая циклические изменения своей конформации. Что это значит? Изменения конформации макромолекул состоят ,в изменении взаимного расположения отдельных функциональных групп макромолекулы, т. е. радикалов аминокислот. Эти группы сближаются и раздвигаются, таким образом, для контактов с окружающей водой и субстратами открываются то одни, то другие аминокислотные остатки. Соответственно изменяются электрические заряды на поверхности макромолекулы. К открывающимся заряженным группам устремляются молекулы воды и соответствующие мелкие противоионы. Движущаяся, изменяющая свою конформацию макромолекула оказывается источником, генератором электромагнитного поля. Правда, противоионы и молекулы воды компенсируют в значительной степени это поле (радиоволны плохо распространяются в морской воде), но конфирмационные колебания — каталитические циклы — происходят с относительно очень малой частотой (1 —105 гц). Такие низкочастотные колебания, такие сверхдлин- ные волны (3-105 — 3 км) вполне хорошо проходят и сквозь морскую воду. Однако сейчас нам важно лишь, что -изменения электрических зарядов на поверхности макромолекул в ходе их конформационных колебаний могут передаваться и на некоторое расстояние от молекулы фермента.
При изменении взаимного расположения аминокислотных радикалов в макромолекуле будут то открываться, то закрываться для контактов с водой и неполярные группы — гидрофобно-гидрофильная мозаика поверхности макромолекулы будет претерпевать циклические изменения рисунка, соответствующие конформационным изменениям макромолекулы. Всякое изменение степени экранированное™ гидрофобных групп, любое изменение рисунка полярно-неполярной мозаики на поверхности макромолекулы вызовет немедленное .изменение ориентации молекул воды около макромолекулы. Молекулы воды, повернутые к полярным группам макромолекулы, отвернутся от возникших на их месте неполярных, образуя связи по преимуществу друг с другом, образуя структуру чистой воды. Это вызовет переориентацию в следующих слоях воды, и волна структурной перестройки воды распространится на некоторое расстояние от поверхности макромолекулы. Таким образом, претерпевающая конфор- мационные колебания макромолекула белка должна быть также и генератором волн структурной перестройки окружающей воды.
Наконец, все эти изменения (и изменения электрических зарядов, и изменения полярно-неполярных соотношений) будут сопровождаться объемными эффектами (о чем мы уже говорили выше). Так, упаковка молекул воды, ориентированных в направлении заряженных групп, и упаковка ее молекул вблизи гидрофобных поверхностей, различна. Конформационные колебаниядолжны, как правило, сопровождаться колебаниями объема системы молекулы белка — вода. Значит, претерпевающая конформационные колебания макромолекула должна быть также и генератором акустических колебаний.
Я излагаю здесь все эти соображения с единственной целью — показать, что между отдельными макромолекулами, испытывающими конформационные колебания, может происходить взаимодействие той или иной природы. Не существенно, сильно оно или слабо — этот вопрос могут решить лишь строгие физические исследования; здесь важно лишь, что оно физически реально. Следовательно, реальна и синхронизация конформационных колебаний многих макромолекул одного сорта. Общеизвестно явление синхронизации, взаимного затягивания по частоте и фазе колебательных систем, генераторов электрических, акустических, механических[13]. Связь между генераторами может быть сколь угодно слабой, но захват, синхронизация все равно осуществляется. Должны возникать синхронно колеблющиеся ансамбли макромолекул. Размеры таких ансамблей должны быть достаточно велики для создания потоков жидкости (а, следовательно, и для перемещения самих ансамблей в жидкости), обеспечивающих преодоление диффузионного барьера в биохимических процессах. Попробуем оценить размеры таких ансамблей.
ПРЕДЕЛЬНЫЕ РАЗМЕРЫ АНСАМБЛЕЙ МАКРОМОЛЕКУЛ В АППАРАТАХ БИОЛОГИЧЕСКОЙ ПОДВИЖНОСТИ
Итак, мы хотим построить макроскопический двигатель, работающий за счет энергии, выделяющейся в ходе катализируемой в самом двигателе реакции. Ясно., что его размеры, размеры такого ансамбля макромолекул должны быть ограничены условиями диффузии. Синхронность конформационных изменений отдельных макромолекул фермента в ансамбле будет определяться отнюдь не только условиями синхронизации в ходе взаимодействия генераторов. Необходимо, чтобы она не нарушалась вследствие различия в доступе субстрата ко всем макромолекулам фермента в ансамбле. Для конформационных колебаний с частотой порядка 1 кгц время образования фермент-субстратно- го комплекса не должно превышать 10_3 и даже 10~4 сек, для колебаний с частотой порядка 10 кгц оно порядка 10-5 сек. Следовательно, при учете скорости диффузии размеры двигательного ансамбля макромолекул должны быть порядка 100— 1000 А. По-видимому, можно найти оптимальные соотношения между частотой конформационных колебаний в ходе ферментативных актов и размерами ансамбля макромолекул, обеспечивающего макроскопические перемещения. Более эффективным
двигателем будет, вероятно, относительно низкочастотный фермент. Чем ниже частота конформационных колебаний, тем больше размер ансамбля, в котором возможна синхронизация. 'Гак, при частоте 10 гц можно пренебречь диффузионными искажениями синхронности в ансамблях размером порядка долей микрона (а то и целых микрон)., Итак, мы приходим к выводу о возможности построения в биохимических (биологических) системах аппаратов перемещения в пространстве на основе макромолекул ферментов, испытывающих в ходе каталитических циклов синхронные конформационные колебания и объединенных в дискретные ансамбли. Такие дискретные ансамбли могут быть органами движения, элементарными движителями. Зато число их в клетке может быть большим — нужно только, чтобы все ансамбли клетка могла обеспечить горючим (например, АТФ).
Таким образом, в ходе эволюции должны возникнуть белки- ферменты, способные расщеплять АТФ с предельно высокой эффективностью преобразования свободной энергии гидролиза макроэргических связей в механическую работу.
Как сказано выше, вероятно, что такие молекулярные осцилляторы, даже разделенные средой (водой), образуют синхронно ьолеблющиеся ансамбли. Однако в ходе эволюционного совершенствования вырабатываются более надежные химические, а не физические способы создания определенных ансамблей специфических макромолекул ‘. Эти способы известны и естественны— самосборка ансамблей макромолекул в результате специфического взаимодействия макромолекул друг с другом, основанного «а химических и физико-химических механизмах. Речь идет здесь об образовании макромолекулярных агрегатов определенной формы, обусловленной последовательностью аминокислот в полипептидной цепи. Существование таких ансамблей не ограничено лишь временем их функционирования. Возникшие молекулярные ансамбли проходят свой обычный эволюционный путь — совершенствование посредством специализации, разделения функций. Автоматический ритмический режим изменения конформаций должен быть подчинен задачам целесообразного движения, что достигается наиболее естественным образом путем регуляции ферментативной активности макромолекулы и тем самым частоты ее конформационных колебаний.
Следовательно, из общих эволюционных, физических и химических соображений мы пришли к представлению об универсальных элементарных аппаратах движения в биологических объектах. Они должны представлять собой дискретные, самособира- ющиеся ансамбли ферментативных макромолекул, претерпеваю
щих синхронные, или во всяком случае, упорядоченные во всем ансамбле конформационные колебания, конформационные циклические изменения, происходящие за счет выделения свободной энергии в экзэргонической реакции.
ЛОГАРИФМИЧЕСКАЯ ЗАВИСИМОСТЬ ЧАСТОТЫ КОНФОРМАЦИОННЫХ ЦИКЛОВ ОТ КОНЦЕНТРАЦИИ СУБСТРАТОВ
Одним из интересных признаков кооперативного функционирования всего ансамбля макромолекул фермента в этом дискретном двигательном аппарате должна быть логарифмическая зависимость частоты конформационных колебаний ансамбля макромолекул от концентрации АТФ. Иными словами,
г л т I, dv г/ 1 dC 1 ,
~ In [АТФ], или — = /( — ; отсюда — = — dC С dv К

где С=[АТФ] — концентрация АТФ, v — частота колебаний, пороговая концентрация АТФ при v = 0.
Физический смысл этой зависимости, по-видимому, таков: число макромолекул фермента (АТФазы), вовлеченных в ферментативный процесс, пропорционально концентрации АТФ. Макроскопические колебания возникают вследствие синхронизации по ансамблю конформационных колебаний отдельных макромолекул. По мере увеличения числа вовлекаемых в процесс макромолекул вклад каждой следующей в синхронные колебания становится меньше. Таким образом, приращение частоты колебаний с увеличением концентрации АТФ (или, что то же, с увеличением числа вовлеченных в колебания макромолекул) обратно пропорционально числу уже вовлеченных макромолекул, величине уже образовавшегося синхронного ансамбля. Количественный анализ этой логарифмической зависимости может дать очень важные сведения о размерах такого ансамбля, о степени кооперативное™ конформационных колебаний отдельных макромолекул. Для такого анализа нужны, однако, дополнительные экспериментальные исследования. Стоит, однако, выяснить, почему вообще частота макроскопических колебаний, может зависеть от числа макромолекул, вовлеченных в такой ансамбль.
Дело в том, что речь идет не о синхронизации осцилляторов, имеющих собственную частоту ко-нформационных колебаний, а о синхронизации механо-химических циклов [41]. В каждом цик- те происходит преобразование энергии макроэргических связей АТФ в механическую работу. Время оборота такого цикла зависит как от каталитической активности, неодинаковой на разных фазах цикла, так и от скорости преодоления механического сопротивления среды. Чем быстрее это сопротивление будет преодолено (сопротивление вязкой среды, инерция перемещаемого тела), тем быстрее наступит фаза расщепления следующей молекулы АТФ. При кооперативной работе макромолекул скорость преодоления механического сопротивления растет. При механических деформациях убыстряется и каталитическое расщепление макроэргических молекул. Тем самым скорость оборота, частота механо-химических циклов растет по мере образования синхронизированного ансамбля макромолекул.
Проведенное рассмотрение логарифмической зависимости частоты механо-хим,ических циклов от концентрации субстрата имеет, возможно, более широкий биологический смысл: логарифмическая зависимость вообще характерна для процессов реагирования живых систем, для физиологических процессов. Логарифмическая зависимость величины ответной реакции от силы воздействия (например, громкость звука определяется логарифмом его интенсивности) — давно известна в физиологии под названием закона Вебера — Фехнера. Закон этот неоднократно критически переосмысливался и млого раз сужалась область его применимости, но зависимость такого рода исе же существует для значительного диапазона интенсивностей раздражителя. Я убежден, что причина логарифмической зависимости самых разных ощущений от интенсивности раздражителя всегда одинакова— наличие кооперативно-ансамблевого аппарата восприятия.
Наиболее важным для дальнейшего выводом из анализа зависимости частоты макроскопических колебаний от концентрации «топлива» является существование предельной частоты таких колебаний. В самом деле, концентрация АТФ в клетке не может превышать некоторую, довольно низкую величину, порядка 10‘4-М0~3М. Следовательно, и частота конформационных циклов ансамблей макромолекул не может быть выше некоторой величины, пропорциональной логарифму предельной концентрации АТФ.
Итак, размеры макромолекулярных ансамблей и частота их конформационных колебаний-биений предельно ограниченны.
РОЛЬ КОНФОРМАЦИОННЫХ ДВИЖЕНИИ И ЛОКАЛЬНЫХ ИЗМЕНЕНИИ pH В АППАРАТАХ БИОЛОГИЧЕСКОЙ ПОДВИЖНОСТИ
Существенной частью механизма описанной выше модели ферментативного механо-химического преобразователя являются силы взаимодействия заряженных групп макромолекулы. Следует отметить, что модель биологической подвижности, основанную на конформационных изменениях полипептидных цепей (спиралей) в результате изменения взаимодействия электрических зарядов боковых групп, предложил в 1929 г. К. Майер^см. [407]). Он полагал, что заряды на полипептидной цепи изменяются при изменении pH внутриклеточной среды. Мысль эта была тогда вполне естественной. Мейархоф, казалось бы, однозначно установил, что мышца сокращается в результате закис- лення, вызванного образованием в процессе гликолиза молочной кислоты [407]. Впоследствии (после работы Лундсгаардта в лаборатории М'ейерхофа) «стало ясно», что мышца сокращается не потому, что образуется молочная кислота, а за счет расщепления макроэргических фосфатов [407], а после знаменитых работ Дж. Хэнсон, X. Хаксли и Э. Хаксли (см. [104]) была оставлена идея конформационных изменений макромолекул, как причина биологической подвижности и стала общепринятой «теория скользящих нитей»— сокращение мышц и другие виды биологической подвижности в результате встречного скольжения не изменяющих свою конформацию фибриллярных структур. Однако «на самом деле» установление уникальной роли макроэргических фосфатов и открытие явления скольжения не опровергает ни ключевого значения изменений электростатических взаимодействий, определяемых в свою очередь уменьшениями pH, ни изменений конформации макромолекул в аппар.ата.х биологической подвижности. «Скольжение» основано на конформационных движениях ТММ-фрагментов миозиновых молекул, циклически замыкающих и размыкающих связь («мост») с нитями фибриллярного актина [86]. Важную роль изменений конформаций макромолекул в явлениях скольжения неоднократно подчеркивал Г. М. Франк [307]. Возможное значение изменений pH в аппаратах биологической подвижности практически ускользает из поля зрения современных исследователей. В соответствии с представлениями, изложенными в гл. 7, основной способ накопления свободной энергии в биохимических системах — наполнение емкости рН-буферов. Там же приведены утверждения, согласно которым биохимическое назначение макроэргических пирофосфатных связей состоит в локальном (в нужном месте и в нужное время) изменении pH, сопровождающем гидролиз этих связей. Я полагаю вероятным, что такой же смысл имеет расщепление АТФ в ходе АТФ-азной реакции при мышечном сокращении. Миозинавая АТФ-аза, расщепляя АТФ, осуществляет быстрое локальное закисление района активного центра — области контакта ТММ-участка молекулы миозина с актином. Такое локальное изменение концентрации водородных ионов приводит к изменению конформации белков и к возникновению механической силы. Если меромиозиновый мост уже соединен с актино- в-ой нитью, то развивается натяжение —форма мышечной клетки изменяется, т. е. мышца сокращается.
На самом деле, ситуация, конечно, много сложнее (см., например, [40, 84, 85]); однаш сейчас серьезных возражений против такого смысла элементарного акта АТФ-азной реакции, по- видимому, нет. Вполне вероятно, что смысл гидролиза макро-эргических фосфатов состоит в обеспечении локальных, контролируемых изменений pH. Так, что, может быть, Нобелевская премия, присужденная более 50 лет назад А. Хиллу и О. Мейер- хофу, была не напрасной (см. [407]).
ДВУХКОМПОНЕНТНЫЕ МЕХАНО-ХИМИЧЕСКИЕ ПРЕОБРАЗОВАТЕЛИ.
ПЕРЕХОД ОТ КОНФОРМАЦИОННЫХ ДВИЖЕНИЙ К СКОЛЬЖЕНИЮ
Итак, мы пришли к выводу, что, следуя физико-химическим критериям, естественный отбор должен привести к возникновению универсальных механизмов, обеспечивающих перемещение клеток в пространстве. Эти универсальные механизмы должны представлять собой ансамбли ферментативных механо-химиче- ских преобразователей энергии. Размер этих ансамблей порядка 1 —10 мкм ограничен диффузией субстратов и продуктов. Частота периодических изменений конформации макромолекул в ансамбле, ограниченная концентрацией АТФ, имеет порядок десятков гц. Непосредственной причиной изменений конформации макромолекул при преобразовании химической энергии в механическую может быть локальное изменение pH.
Ясно, что все это соответствует реально существующим универсальным двигательным аппаратам клеток — ундулоподиям — жгутикам и ресничкам [259]. Специализация ундулоподий может привести к усложнению их структуры и разделению функций — ферментативное расщепление макроэргических субстратов может осуществляться в специализированном активном центре в отдельной макромолекуле, а изменение размеров—.производство механической работы — в других макромолекулах. Такое разделение единого ма.кромолекулярного механо-химического преобразователя на специализированные макромолекулы обусловлено, вероятно, задачей увеличения эффективности механо-химиче- ских циклов (А. Е. Букатина). В самом деле, в нашей модели- пружине с двумя одноименными зарядами требовалось довольно искусственное предположение о полном уничтожении зарядов субстрата в результате, например, декарбоксилирования— замене сильных кислотных групп карбоновых жислот слабой угольной кислотой. Кроме того, «обратный ход» — распрямление пружины — сопряжен с преодолением более алт менее сильного взаимодействия внутри пружины. Существенное облегчение «обратного хода», предотвращение или сильное «уменьшение трат энергии на этот процесс достигается в двухкомпонентной системе— взаимодействии двух «независимых частей» .пружины, например, по тому же электростатистическому механизму. Теперь обратный ход — «расслабление»—может осуществляться почти без затрат энергии.
Переход к двухкомпонентным .системам, преобразование химической энергии в .механическую работу— переход от совершения работы за счет конформационных движений макромолекулы самого фермента — к «скольжению» — активному «встречному» перемещению двух макромолекулярных компонентов обеспечивает максимальную эффективность механо-химических преобразований.
В двухкомпонентных системах активное встречное движение макромолекулярных компонентов, т. е. сокращение системы может осуществляться за счет конформационных движений. Обратный ход — [расслабление — может осуществляться без изменения конформации компонентов системы. Для этого встречное движение— скольжение — должно осуществляться в результате относительно большого числа циклов изменения конформации одного из компонентов (периодического замыкания и размыкания мостов ТМ1М и актина), а обратное движение — скольжение несцепленных макромолекул должно осуществляться пассивно.
Такая двухкомпонентность—наличие актина и миозина — ха|ра,ктерна для большинства систем биологической .подвижности [86].
Итак, логика рассмотрения физико-химических критериев естественного отбора при эволюционном построении аппаратов перемещения в пространстве цривела нас от изменений поверхностного натяжения к изменяющим свою конформацию ферментам, и, наконец, к двукомпонентным механо-химическим ферментативным преобразователям, м.акромолекулярные компоненты которых не обязательно .изменяют свою конформацию.
ОТ УНДУЛОПОДИЙ к МЫШЦАМ
Пока мы обращали внимание лишь на термодинамическую эффективность и на скорость (кинетику) преобразования энергии в аппаратах перемещения организмов в пространстве. Однако имеются еще чисто механические .критерии совершенства аппаратов перемещения в пространстве. Совершенство перемещения в пространстве в значительной степени определяется абсолютной скоростью движения. Чем быстрее организм достигнет источника пищи или избегнет опасности, тем больше вероятность его выживания. Абсолютные скорости перемещений в пространстве определяются размерами рычагов. Размеры ундулоподий, как было показано, ограничены диффузией субстратов и продуктов ферментативного катализа. Единственным способом увеличения скорости перемещения посредством ундулоподий оказывается увеличение их числа у каждого организма — клетки. Так возникают клетки, покрытые большим числам ресничек.
Было бы о.чень интересно рассчитать предельно возможную скорость перемещения организмов .посредством жгутиков и ресничек. Однако расчет этот очень сложен. Движение клеток посред-«ствам ресничек было предметом ряда исследований (см. [259]). Как показал Л. Н. Серавин [260, 231], движение инфузорий осуществляется посредством движения «чехла» воды, прилегающего ,к поверхности клетки. Реснички при биениях .вызывают движение этого поверхностного слоя. Таким образом, основная энергия затрачивается на преодоление вязкого сопротивления б этом слое движению ресничек. Это следует .из весьма низких значений чисел Рейнольдса, характерных для движения инфузорий и жгутиконосцев [368, 464, 472]. Следовательно, предельная скорость движения таких организмов определяется трением. При биениях ресничек в прилегающем слое воды накапливаются импульсы и клетка «разгоняется»—скорость ее перемещения относительно (отдаленных слоев воды или) этого прилегающего слоя воды постепенно возрастает. Ясно, что существует предел этой скорости, заданный частотой биения ресничек. При некоторой скорости движения клетки реснички цр,и биении не будут передавать импульс окружающей среде. Таким образом, предельная скорость движения клетки будет лимитирована предельной частотой биений, лимитируемой, в свою очередь, диффузией субстратов («топлива») к ферментативным мехаио-химическим преобразователям. Поэтому задача увеличения .максимальной скорости перемещения в пространстве может быть решена лишь посредством увеличения скорости движения «весел». Это нельзя сделать посредством увеличения частоты биений, но это, казалось бы, можно осуществить посредством увеличения длины рычага. Мы уже видели, что размеры у.ндулолодий также лимитированы диффузией. Поэтому можно представить себе удлинение рычага лишь при условии, что большая часть этого рычага погружена в среду, содержащую «топливо» (например, АТФ). Это значит, что большая часть двигательного аппарата должна находиться внутри клетки. Мы приходим к новому принципу движения — клетка движется посредством максимального рычага, равного ей и даже превосходящего ее размеры, и движение совершается посредством изменений фор,мы клеток. Для этого необходимо прикрепить «сократительную» нить изнутри к двум максимально отдаленным участкам клеточной стенки. При сокращении этой нити клетка будет (более или менее резко) изменять свою форму. Скорость движения клетки будет определяться скоростью движения «концов» клетки при ее, например, сжатии и распрямлении. Такой аппарат является, по сути, мышцей.
Ясно, что организмы, вставшие на этот путь эволюции, будут увеличиваться в размерах: чем больше размеры рычага, клетки, органа, организма, тем выше абсолютная скорость движения организма.
В настоящее время трудно оценить из общих соображений величину выигрыша в абсолютной скорости движения при переходе от ресничек к мышцам. Воспользуемся поэтому экспериментальными исследованиями и их анализом, выполненными
РИС. 24. Зависимость максимальной скорости перемещений
У су /сен

в воде от длины тела различных животных [346]
to Z Wj to31, см
/ — жгутиковые; 2 — ресничные; 3— весточогие ракообразные; 4 — рыбы* 5 — киты
В. В. Шулейкиным и его сотрудниками еще в 1939 г. [347]. На рис. 24, взятом из [346], приведены данные о зависимости максимальной скорости перемещения в .воде различных животных от длины их тела. Видно, что для движения посредством ундулоподий характерна предельная скорость порядка 10-1 см/сек. Для движения веслоногих рачков, обусловленного сокращением мышц, парящего движения взвешенных в воде организмов характерна предельная скорость порядка 10 см/сек, рыбы движутся со скоростью порядка 5-10—5-102 см/сек, и, наконец, китообразные с предельной скоростью порядка 103 ом/сек.
Следовательно, переход от движения посредством ундулоподий к движению посредством мышц позволяет в принципе увеличить абсолютную скорость перемещения в пространстве на четыре порядка (от 10'1 до 103 см/сек). Таким образом, направление эволюции от ундулоподий к мышцам характеризуется большим эволюционным потенциалом, скорость .перемещений в пространстве оказывается мощным фактором биологической эволюции.
Эволюция в направлении совершенствования перемещений в пространстве посредством мышц приводит к формированию множества новых свойств :и приспособлений. Становится возможным не только полное овладение водной стихией, но и выход на сушу и полет в воздухе [140]. Одним из важнейших следствий эволюционного совершенствования перемещений в пространстве посредством мышц является образование многоклеточных организмов.
ПРОИСХОЖДЕНИЕ И БИОЛОГИЧЕСКИМ СМЫСЛ -МНОГОКЛЕТОЧНОСТИ
Переход от ундулоподий к мышцам сопряжен с возникновением многоклеточности. В самом деле, каждая сокращающаяся (мышечная) клетка с сократительной нитью, прикрепленной к внут- генней стороне оболочки клетки, функционально равнозначнаодной ресничке или одному жгутику. Задача управления движением, осуществляемым с максимально возможной скоростью, решается посредством 'объединения в один организм некоторого числа специализированных, сокращающихся лишь в определенном направлении мышечных клеток. Специализация (мышечных) клеток на совершении сокращения лишь одного вида — естественный путь эволюционного совершенствования их работы. Образование многоклеточных организмов — естественное следствие такой специализации. Колоссальное увеличение скоростей перемещения, возникающее при замене ресничек мышцами, дает многоклеточным макроорганизмам огромные преимущества в естественном отборе.
Миофибриллы, обеспечивающие быстрое изменение формы клеток, появляются уже у одноклеточных организмов — инфузорий[14]. Однако действительно эффективным средством не только быстрого изменения формы, но и перемещения в пространстве миофибриллы становятся при специализации всей клетки на движении в данном направлении. Быстрое перемещение в произвольном направлении в пространстве может быть достигнуто лишь у многоклеточных организмов.
Эволюция многоклеточных животных в дальнейшем в значительной мере определялась совершенствованием самих органов движения я аппаратов управления их работой— системой рецепторов, нервных клеток и нервных центров. Глубокие обобщения в проблеме эволюции локомоторных органов беспозвоночных животных принадлежат Л. А. Зенкевичу (112].
Из сказанного не следует, что переход от ундулоподий к мышцам и специализация мышечных клеток — единственная причина возникновения многоклеточное™ [15]. Многоклеточность обусловливает предельное совершенство перемещений в пространстве, предельно возможную абсолютную скорость перемещений и управления этими перемещениями.
Однако наиболее общей причиной многоклеточности является выгодность специализации функций, частным случаем которой оказывается и возникновение мышечных клеток.
Ясно, что преимущественный синтез белков для обеспечения какой-либо преобладающей в данный момент функции основан на репрессии синтезов других белков, не нужных в данный момент. Это требует очень громоздкой и неэкономичной системы регуляции считывания матричных текстов — больших концентраций разных на разных стадиях жизни клетки белков-репрессоровг сложного аппарата управления синтезом самих репрессоров, существования механизмов целесообразной дерепрессии (активации) синтеза очередных белков.
Помимо громоздкости и неэкономичности такой системы существенна медленность включения очередных функций — необходимо длительное время для прекращения уже идущих синтезов, появления репрессоров, дерепрессии очередных генов и т. д.
Специализация клеток состоит прежде всего в специализации их генома — утрате или прочной репрессии большей части генов. Предельно специализированной клеткой является эритроцит млекопитающих. Он содержит практически (по массе) лишь один белок — гемоглобин— и выполняет лишь одну функцию: транспорт кислорода и углекислоты. В гораздо более сложных мышечных клетках преобладает синтез сократительных белков.
Итак, специализация — основной путь совершенствования функций — достигается лишь в многоклеточных организмах. Специализация обеспечивается дифференциацией клеток — стойкой репрессией .или даже утратой значительной части наследственных текстов в процессе онтогенеза.
Ясно, что специализация клеток в многоклеточном организме' возможна лишь при возникновении системы взаимосвязи специализированных клеток, объединяющей их в единый организм — «государство клеток», как говорил Р. Вирхов. Такая связь осуществляется посредством химических (гормональных) или электрических (нервных) сигналов.
Само существование многоклеточных организмов сопряжено с новыми эволюционными «заботами» — обеспеченней определенной «внутренней среды», целостности, регенерации при повреждении и т. п. Обсуждение этих проблем выходит за пределы задач этой книги.
ОБЗОР РЕАЛЬНЫХ МЕХАНИЗМОВ БИОЛОГИЧЕСКОЙ ПОДВИЖНОСТИ
На самом деле имеется несколько типов явлений биологической подвижности. Прежде всего это движение протоплазмы’. Потоки протоплазмы омывают или даже увлекаюте собой клеточные органеллы, осуществляя активное перемешивание внутриклеточного содержимого. Кинетический смысл такого перемешивания очевиден. Движение протоплазмы происходит в клетках растений, животных, низших грибов. Удивительным образом его механизм, несмотря на примерно 200-летнюю историю исследований, все еще не ясен. Об этом свидетельствует чрезвычайное изобилие остроумных гипотез и теорий, объясняющих этот феномен. Яркая особенность движения протоплазмы состоит е правильной периодической смене направления движения потоков. Мне представляется наиболее вероятной причиной движения протоплазмы синхронизация конформационных движений ансамблей специализированных макромолекул. Изменения направления — колебания вектора напряжения — могут быть результатом явления, аналогичного «волнам структурной перестройки» — перекристаллизации то по одной, то по другой затравке. Следует отметить, однако, что Н. С. Аллен [361] сообщила об обнаружении в эндоплазме нителлы нитей, аналогичных жгутикам, биением которых обусловлено движение протоплазмы.
Ко 2-му типу явлений биологической подвижности можно отнести амебоидное движение, очень часто неотличаемое от движения протоплазмы. Однако мне представляется их различие существенным — образование и исчезновение псевдоподий сопряжено не только с перетеканием протоплазмы, но и образованием (исчезновением) поверхностной мембраны, а также с изменением кортикального слоя клеток типа гель^±золь (см. [362]).
3-й тип явлений биологической подвижности движения хромосом при митозе и мейозе. При этом существенна все еще необъяс- ненная правильность пространственного взаиморасположения гомологичных хромосом. Расхождение хромосом к соответствующим центриолям происходит в результате сокращения нитей веретена, пучков, микротрубочек, состоящих из белка тубуллина (см. [362]) ‘.
Своеобразные движения 4-го типа осуществляются в результате изменения тургора — гидростатического давления, обусловленного осмотическими или иными механизмами. Так, посредством амбулакральной системы перемещаются иглокожие. Изменения тургора обусловливают движения растений — открывание и закрывание устьиц, опускание и поднятие листьев.
К 5-му типу движений относится перемещение бактерий (прокариот вообще) посредством жгутиков. Каждый жгутик бактерий состоит обычно из трех фибрилл, образованных последовательно соединенными глобулами белка флагеллина. В большинстве случаев нити жгутиков длиной порядка микрон и диаметром около 120 А, лишены оболочки (мембрана).
Как следует из работ недавнего времени [366], механизм дви- женин жгутиков бактерий неожиданно своеобразен. Сам жгутик является лишь пассивным органом — двигатель расположен в основании жгутика в мембране — это базальное тельце. Движе- ние осуществляется посредством вращения базального тельца в мембране. Существует представление, в соответствии с которым это вращение непосредственно обусловливается градиентом концентраций ионов водорода [269, 366]. Таким образом, в биологических структурах давным-давно изобретено колесо (!). Более того, это колесо аналогично ротору электродвигателя. Во всех этих типах движений скорость перемещении имеет лишь второстепенное значение. Этим объясняются малые абсолютные значения скоростей перемещения этих типов.
Скорость перемещений становится критерием естественного отбора, фактором биологической эволюции при решении задачи поиска подвижной добычи или активного избегания опасности. Эта задача решается посредством движений трех различных типов. При своеобразных движениях 6-го типа происходят быстрые изменения формы тела инфузорий и некоторых жгутиковых. Ярким примером такого движения является быстрое сокращение стебелька сувойки с укорочением в несколько раз за сотые доли секунды (скорость движения порядка 20 см/сек). Сокращение этого типа осуществляется за счет изменения упаковки (конформации субъединиц) лолиглобулярнсй белковой фибриллы —- спазмонемы (мионемы). Непосредственной причиной такого изменения является увеличение концентрации (активности) ионов, кальция [363]. Такой механизм позволяет сувойке и ее родственникам быстро, «мгновенно» выходить из опасного контакта с врагом. Однако обратное движение, лимитируемое, по-видимо- му, откачкой ионов кальция специфическими насосами, происходит медленно. В то же время для любого перемещения в пространстве ,на расстояния, существенно превышающие размеры тела, нужны быстрые, многократные, периодические движения.
Отличие спазмонемы от мышцы состоит в том, что в первом случае возможен лишь один акт сокращения при увеличении концентрации ионов кальция вблизи сократительной системы, и производимая работа определяется непосредственно величиной изменений концентраций (активностей) ионов кальция, а в мышце— после запуска ионами кальция начинается циклический процесс, на один импульс ионов кальция эсвобождаются многие сотни порций энергии макроэргических фосфатов, происходит резкое возрастание мощности. Пределом совершенства таких механизмов являются перемещения 7-го и 8-го типов — посредством жгутиков и ресничек эукариот и посредством мышц. Сведения об устройстве и функционировании жгутяков и ресничек можно найти в книгах Л. Н. Серавина [259] и Н И. Арронега [10], а также в [362, 402]. Поэтому нет необходимости подробно излагать имею- шиеся данные. Замечательна однотитность строения жгутикови ресничек эукариот. Жгутики н реснички (ундолоподии) всех эукариотных организмов состоят из 9 пар периферических фибрилл и 2 центральных фибрилл, погруженных в относительно бесструктурную протоплазму и окруженных трехслойной мембраной. Сами же фибриллы, как и в жгутиках бактерий, состоят из глобулярных макромолекул белка тубулина и динеина, полимеризированного в виде нити или трубочки. Важным научным достижением было создание моделей жгутиков и ресничек, способных к ритмической активности. Это впервые удалось сделать Гофман — Берлин гу IB 1953—1954 гг. Он показал, что у мертвых, экстрагированных 50%-ным раствором глицерина, простейших и сперматозоидов после помещения в раствор, содержащий хлористый калий (в концентрации 0,1 М), ионы магния (в концентрации порядка 10~3М), буферную смесь, поддерживающую pH близ 7—8 и АТФ в концентрации порядка 10_3М, возобновляются энергичные биения жгутиков. С тех пор такие глицеринизированные препараты ундолоподий были получены из разных организмов (см. книгу Н. И. Арронета [10]). При обработке глицерином и другими экстрагирующими веществами разрушается клеточная мембрана, вымывается протоплазма, и остаются лишь фибриллы. Образующие их макромолекулы белка расщепляют АТФ. И этот процесс, эта АТФ-азная реакция сопровождается ритмическими достижениями фибрилл. В моделях частота биения жгутиков и ресничек пропорциональна логарифму концентрации АТФ. Значение этого обстоятельства рассмотрено выше. Движение ундолоподий отражает, вероятно, циклические изменения конформации — кон- формационные колебания — макромолекул фермента в каталитическом акте. Способность к таким колебаниям, равно как и способность образовывать многомолекулярные ансамбли, должна быть свойственна самим белковым макромолекулам, образующим фибриллы ундулоподий.
Наблюдавшиеся мною [335—339, 458] и другими авторами [293, 323] синхронные в макрообъеме конформационные колебания в растворах белков актимиозинового комплекса или креа- тинкиназы происходят без субстратов и не отражают непосредственно циклические изменения макромолекул в ходе ферментативного катализа. Эти колебания беспорядочны, непериодичны, однако в них, вероятно, проявляется способность этих белков к синхронным колебаниям в ходе ферментативного катализа. Однако колебания, посредством которых работают аппараты биологического перемещения в пространстве, должны осуществляться за счет расщепления макроэргических соединений, вероятнее всего за счет расщепления АТФ.
Так, Паутард (см. [362, с. 97]) наблюдал колебания (сокращения частиц геля контр актильного белка, выделенного из ундулоподий сперматозоидов рыб, в среде с АТФ в присутствии ионов магния.
Весьма интересные наблюдения описаны А. Оплаткой и Р. Ти- рошем [440, 441]. В работах А. Оплатки и Р.Тироша [440] описано явление, возможно имеющее прямое отношение к приведенным рассуждениям. Авторы показали, что в растворах универсально- ю 'Сократительного белкового комплекса ,актомиозина, выделенного из плазмодия (или же из мышц кролика) и помещенного в узкие капилляры, в присутствии АТФ происходит энергичное движение потоков раствора. Направления этого движения время от времени ритмически изменяются.
Оплатка и Тирош полагают, что движением растворах белков актомиозинового комплекса происходит в результате конформационных колебаний макромолекул за счет энергии АТФ. Это их предположение соответствует всей системе рассуждений, приведенных выше. Однако прежде всего необходимо подтверждение феномена, описанного этими авторами, в новых экспериментальных исследованиях \
Поскольку биохимии и биофизике мышечного сокращения посвящено большое число фундаментальных работ [20, 40, 81, 82, 84, 85, 104, 376], и особенно книга В. И. Дещеревского [86], нам нет необходимости описывать современные представления по этому вопросу.
Глава 10
УПРАВЛЕНИЕ ДВИЖЕНИЕМ. НЕРВНАЯ СИСТЕМА
Совершенствование перемещений в пространстве. Возникновение рецепторов и нервов. Необходимость концентрации нервных центров
(образование мозга) и рецепторов
для обеспечения высокого совершенства управления движением.
Эволюционный потенциал эгапа эволюции, начинающегося с выработки .аппаратов активного перемещения, обусловлен в основном возможностью векторизации перемещений в пространстве, выработки механизмов строго направленного движения. Наиболее сложно при этом обеспечение перемещений к произвольно движущейся цели. Такая задача возникла в процессе эволюции после выработки механизмов произвольного перемещения к относительно неподвижным целям. Но особенно острой она становится при «взаимодействии» жертвы и хищника. Речь здесь идет о высокосовершенной жертве, способной к сложным произвольным движениям, и соответственно о не менее совершенном хищнике. Преследование совершенной жертвы совершенным хищником сопряжено с предельно совершенным управлением перемещений в пространстве.
Как хищник, так и жертва должны прежде всего быть способны воспринимать сигналы, информацию о местонахождении «друг друга». Оба они должны, кроме того, быть в состоянии прослеживать траектории взаимного движения, обобщать увиденное и услышанное, создавать абстрактный образ, т. е. находить общую закономерность перемещения «партнера», запоминать обобщенную картину, сопоставлять ее с реальной траекторией, направлять свое движение в ожидаемую точку пространства и, победив, съесть жертву или, избежав съедения, продолжить свое существование.
Отсюда ясно, что предельно совершенные жертвы и хищники должны обладать предельно совершенной нервной системой, способной к анализу поступающих сигналов, к сопоставлению образов (отсюда и произошло слово соображение «со — ображение»), обладающей памятью, способной к абстракции, к принятию и реализации решений. Таким образом, при решении задачи управления движением в ходе эволюции рано или поздно должны возникнуть предельно совершенные нервная система и система рецепторов.
В самом деле, представим себе жертву, которая может перемещаться в пространстве не менее совершенно, чем хищник. Хищ- ■ ник не в состоянии догнать жертву и превратить ее в добычу, но
on зато может подстеречь ее, пересечь ее возможную траекторию движения. Но и жертва «сохранится в пэтомках» лишь при условии учета и разгадки возможных маневров хищника.
Конечно же, рассматриваемая здесь ситуация — далеко идущая абстракция. Равенство скоростей и длительности перемещения жертв и хищников практически невероятно. В действительности же либо хищники оказываются намного мощнее жертв, либо жертвы в ходе естественного отбора становятся столь мощными, что перестают быть жертвами. Сложная картина взаимоотношений жертв и хищников, эволюционная роль специализации, деспециализации, соотношения мощностей хищников и жертв рассмотрена И. И. Шмальгаузеном в его книге «Факторы эволюции» [332]. Сложнее поведение при взаимодействии жертв и хищников, выражающееся в экстраполяции траекторий перемещения, подстерегании, устройстве засад, срезании криволинейных траекторий при преследовании, сложные произвольные изменения направления пути преследуемой жертвы — все это наблюдали многие зоологи, охотники и охотоведы. Общая концепция построения произвольных движений, основанная на экстраполяции и анализе перемещения с учетом конечной цели, была развита в 30—40 гг. Н. А. Бернштейном [26]. На протяжении ряда лег Л. В. Крушинский экспериментально исследует способность животных экстраполировать траекторию движущегося предмета [159—161].
Попробуем теперь, как и в предыдущих задачах, представить себе ход эволюции и ее результаты на основании общих соображений.
В гл. 9 мы уже рассматривали причины возникновения много- клеточности, однако мы забежали вперед — многоклеточные организмы с мышцами-клетками в начальный период своего возникновения могут управлять движением существенно хуже, чем высокосовершенные одноклеточные или многоклеточные организмы, движущиеся посредством ресничек или жгутиков. Поэтому начнем наш анализ возможных эволюционных путей возникновения аппаратов управления движением с рассмотрения перемещений реснитчатого организма.
Такой «абстрактный организм» имеет вид сферической клетки, покрытой ундулоподиями — респичками или жгутиками (он похож на ныне живущих сферических инфузорий или колониальных сферических водорослей — вольвокса и его родственных форм).
Ясно, что движение может осуществляться с большой скоростью лишь организмами, форма тела которых приспособлена к быстрому перемещению *. Из гидродинамических соображений следует необходимость эволюционного преобразования сферических организмов в более или менее вытянутые эллипсоиды.
У таких активно, но беспорядочно движущихся организмов намного усилится взаимодействие со средой, увеличится сечение захвата пищи, т. е. для них станет возможным преодоление диффузионного барьера. Однако жизнь быстро перемещающихся эллипсоидов может быть достаточно благополучной лишь в среде с равномерным распределением пищи в пространстве, например, в растворах питательных веществ (так живут многие микроорганизмы— бактерии, протозоа). При неравномерном распределении пищи в среде, при существовании пищи в виде дискретных кусков живого или неживого вещества, беспорядочное броунирова- ние организмов оказывается недостаточно эффективным.
Для того чтобы равномерно покрытый ундулоподиями эллипсоид мог перемещаться в направлении дискретных кусков пищи или убегать от нападающего на него, как на жертву, хищника, нужно совершенное управление работой ундулоподий, обеспечивающее целенаправленное перемещение в пространстве.
Наиболее естественный механизм такого управления — прямое действие сигналов, исходящих от цели движения, на ундуло- подии.
Таким образом, первым шагом на эволюционном пути к управлению перемещениями в пространстве должны быть отбор способов изменения двигательной активности ресничек и жгутиков под влиянием химических, акустических или световых сигналов. Это значит, что рецепторами могут служить модифицированные реснички. Как же они смогут обеспечить функции рецепторов? Для этого необходимо либо эволюционное усиление изначально присущей ресничкам чувствительности к свету, к вибрациям (и другим акустическим воздействиям), к химическому составу среды или электромагнитным полям, либо сопряжение работы ресничек с функционированием молекулярных или надмолекулярных комплексов, чувствительных к указанным воздействиям.
Прежде чем углубляться в проблему построения рецепторов, вернемся к нашему реснитчатому эллипсоиду. Представим себе, что его реснички чувствительны к свету, причем освещение вызывает замедление и даже изменение направления — реверсию *. Ясно, что при движении эллипсоид будет поворачиваться к источнику света и медленно приближаться к нему. Ускорение биений под влиянием света приведет к повороту от света и уходу в темноту. То же справедливо для звука, химических сигналов, электрических и магнитных полей. Поведение эллипсоида определяется вынужденными движениями. Учение о вынужденных движениях при воздействии разных факторов на растения и животных создал в конце прошлого века и в нашем веке развивал его уже упоминавшийся мною великий физиолог Ж. Леб. Его книги, среди
! Анализ механизмов управления движением жгутиковых и ресничных простейших дан в работе Ю. А. Лобаса и В. С. Маковского [165].
193
7 С. Э. Шнолькоторых «Организм как целое», «Динамика живого вещества», «Вынужденные движения, тропизмы и поведение животных» [167—169] не только являются замечательными примерами развития биологии тех лет, но полны ценнейших мыслей и наблюдений, не утративших интереса и для современной биологии.
Однако нас интересуют не только тропизмы — вынужденные движения, обусловленные различными полями с очень небольшими градиентами,— но н перемещения в направлении дискретных объектов (или в противоположном направлении). Поэтому следует рассмотреть возможные эволюционные пути возникновения органов чувств.
Возникновение первичных рецепторов из ундулоподий. Биение ресничек вызвано синхронными конформационными колебаниями, обусловленными ферментативными циклами в АТФ-азной реакции. Любые факторы, влияющие на эти процессы, могут использоваться в ходе эволюции в качестве сигналов о состоянии окружающей среды. Среди таких возможных факторов на первом месте стоят химические, а именно, градиенты концентрации различных химических веществ. Вещества, замедляющие биение или вызывающие реверсию биений, вызовут поворот реснитчатых эллипсоидов и их медленное продвижение к источнику данного вещества, т. е. перемещение против градиента концентрации активного вещества. От веществ, ускоряющих биение, эти организмы будут удаляться — двигаться по градиенту концентраций.
Такое реагирование вызывают вещества, дающие эффект одного знака. На самом деле, для биохимических реакций зависимость скорости процесса от концентрации какого-либо вещества обладает по меньшей мере одним экстремумом — обычно максимумом (оптимумы pH и т. п.). Для многих же воздействий при монотонном изменении концентрации какого-либо вещества или силы воздействия характерна неоднократная смена знака эффекта. В этих случаях поведение нашей модели будет более сложным.
Начнем с простого. Какие химические вещества влияют на биение ундулоподий? В общем виде ответ «прост»: субстраты, кофакторы, ионы водорода и гидроксила, специфические конкурентные и аллостерические регуляторы (ингибиторы) АТФ-азной реакции и, кроме того, все химические вещества, влияющие на конформационную лабильность и синхронность конформационных колебаний макромолекул белков. Среди последних нужно прежде всего отметить, вероятно, двувалентные катионы и различные детергентные и гидрофобные молекулы. Так, увеличение концентрации ионов магния ускоряет биение ресничек, а увеличение концентрации ионов кальция вызывает реверсию биения ресничек [165].
Таким образом, грубая химическая ориентация при перемещениях организмов в водной среде может достигаться при пали-
чии в среде различных веществ. Хемочувствительныеундулоподии •служат первичными рецепторами вкуса и запаха у первичных организмов. «Запах» это — вкус воздуха».
Первичные фоторецепторы. Для того, чтобы ундулоподии могли реагировать на свет нужно, чтобы АТФ-азная реакция или сопровождающие ее синхронные конформационные циклы макромолекул фермента были сенсибилизированы к свету. Макромолекулы белков поглощают ультрафиолетовое излучение с длиной волны около 280 нм (за счет остатков ароматических аминокислот), а также более короткое излучение в области 250, 220 нм (за счет тиолов и пептидных связей). Такое короткое излучение несет слишком много энергии, что может вызвать разрушение макромолекул, и поэтому оно мало пригодно для сигнальных функций. Больше пользы в этом отношении можно получить от 'использования «нетравматического» -излучения в диапазоне 400—800 там, т. е. от видимой области.
В качестве сенсибилизаторов двигательного аппарата ундуло- подий к воздействию видимого света эволюция могла бы избрать несколько пигментов — обязательных участников внутриклеточных предельно совершенных биохимических реакций (мы исходим из того, что биохимические процессы уже предельно совершенны). Возможными кандидатами на роль фотохимических сенсибилизаторов могли быть флавины, порф.ирины, пиридин- нуклеотиды, каратиноиды. Почему на самом деле эволюция избрала для этой цели преимущественно каротиноиды, до сих пор не ясно. Много десятилетий назад уже высказывались предположения о возможной роли каротиноидов в качестве обязательных участников окислительных превращений в клетках. Мне кажется, однако, что каротиноиды вошли в процессе эволюции в биохимию в связи с необходимостью защиты от вредного действия ультрафиолета. Хорошо поглощая ультрафиолет, каротиноиды могут «разменивать» в процессе флуоресценции жесткие кванты на безвредные кванты видимого света или осуществлять за счет поглощенной энергии какие-либо полезные сопряженные процессы. Выполняя роль светозащитных очков, каротиноиды оказались удобными пигментами, воспринимающими видимый свет. То обстоятельство, что первоначальная функция каротиноидов состояла в защите от ультрафиолета, объясняет их загадочное обязательное присутствие во в-сех эктодермальных тканях (клетках), а также в листьях растений. Итак, каротиноиды обязательно должны содержаться в ундулоподиях, и поэтому все ундулоподии неизбежно должны обладать чувствительностью к видимому свету *.
Нам осталось теперь «©сего лишь» предложить (механизм фотохимически сенсибилизированного управления циклом процессов, составляющих основу биения и реверсии биений ресничек. Впрочем, нельзя так много требовать от дедукции! Как известно, фотосенсибилизированное изменение АТФ-азной реакции мышечных белков, открытое Сцент-Дьёердьи, изучалось многими авторами (см. например, [186а, 248]). Для понимания возможного механизма фотосенсибилизации АТФ-азы необходимо знать детальный механизм этой реакции. Такого знания еще нет. Пока нам достаточно того, что фотохимические процессы в каро- тиноидах или во флавинах, порфиринах, пиридиновых кофер- ментах способны, в принципе, регулировать биение ресничек (см. [164, 200]).
Первичные механорецепторы. Восприятие акустических сигналов. При рассмотрении механизма движения ансамбля макромолекул, способа синхронизации конформационных циклов всех макромолекул ансамбля в предыдущих главах мы уже обращали внимание на возможную роль в этом сложном кооперативном процессе акустических полей. В принципе, внешние акустические ноля могут способствовать или препятствовать синхронизации конформационных изменений макромолекул при ферментативном катализе в двигательном ансамбле, изменяя тем самым силу или частоту биений ундулоподий. Ясно, что речь может идти об акустических сигналах с частотами, соответствующими диапазону возможных частот при ферментативных реакциях, т. е. я* 0—200 гц.
К сожалению, звуковые колебания с такими частотами соответствуют столь длинным волнам, что градиенты акустического давления не могут восприниматься не только отдельными ресничками, но и целыми клетками. В самом деле, скорость звука в воде равна 1500 м/сек. При частоте 100 гц длина »олны в воде составит 15 м. Легко понять, что посредством столь длинных волн нельзя передавать информацию о локализации источника этих акустических сигналов воспринимающей системе с размерами порядка микрон (линейный размер клетки около 100 мк.)
Вывод о невозможности восприятия акустических сигналов реальных биохимически частот рецепторами с размерами порядка десятков микрон основан на принятии величины скорости звука в воде 1500 м/сек. Возможно ли исходить из других величин? Мы уже отметили, что в работе Б. Б. Кудрявцева и особенно в работах А. П. Сарвазяна и др. [253] показано, что в гелеобразных средах, а также в трубках с податливыми стенками, скорость звука на три порядка меньше, чем в воде, так как в этих условиях нормальная продольная звуковая волна не распространяется. Акустическая энергия переносится короткими поперечными волнами.
Таким образом в гелях звуковые волны характеризуются (при «слышимых» частотах) длиной волны порядка 10-2—1 см. К сожалению, это важное само по себе обстоятельство не создает условий для звукового общения наших микрообъектов — реснитчатых, одноклеточных эллипсоидов. В самих этих организмах выполняются условия Сарвазяна, в них скорость звука и соответственно длина акустической волны малы. Но между организмами находится вода. Медленная звуковая волна не будет возбуждаться в частичках геля, взвешенных в воде *. Следовательно, не может быть собственно акустических (волновых) рецепторов у нашего эллипсоида.
Зато реснички прекрасно приспособлены для восприятия гидродинамических импульсов, т. е. механических возмущений, создаваемых потоками воды. Условия возникновения гидродинамических эффектов при движении микрообъектов в жидкостях отнюдь не просты. Отчасти мы затрагивали их в предыдущей главе при анализе скорости движения посредством ресничек. При очень малых числах Рейнольдса, характерных для таких объектов, потоки жидкости будут быстро замедляться вблизи движущегося объекта. Тем не менее, реснички могут воспринимать и весьма слабые механические возмущения. В высокой степени это восприятие зависит от периода (частоты) следования возмущений. По-видимому, механические колебания, совершаемые не в резонансе с собственными ферментативными циклами ундулоподий, будут нарушать работу ресничек. Чем ближе частота внешних сигналов к собственной частоте ферментативного цикла, тем больше вероятность резонансного усиления биений ресничек. Помимо чисто механического влияния пульсации скорости потоков жидкости на механохимические и регуляторные циклы, возможно и влияние на рецептор посредством изменения концентрации в прилегающем слое каких-либо активных веществ посредством преодоления диффузионных ограничений. В самом деле, вся проблема перемещения в пространстве рассматривается нами в связи с диффузионными ограничениями подвода к клетке питательных веществ или отвода от нее продуктов метаболизма. Таким образом, первичный механорецеп- тор может работать как хеморецептор.
Моделью такого рецептора может служить метод измерения гидродинамических потоков, использованный в работе [454][16]. Этот метод основан ,на зависимости электрического тока в полярографическом электроде или в термисторе от скорости омывающих их потоков жидкости. Ток в полярографическом электроде определяется стационарной концентрацией реагирующего на поверхности электрода вещества (например, кислорода). Ток лимитируется диффузией. Перемешивание приводит к увеличению стационарной концентрации реагирующего вещества. Интенсивность движения жидкости пропорциональна току в электроде. Аналогичным датчиком слабых потоков жидкости может быть термистор, сопротивление (температура) которого (при постоянном нагреве) зависит от скорости теплоотвода — от скорости омывающих его потоков жидкости. Так или иначе, ясно, что первичный механоакустический рецептор может быть создан на основе ундулоподий.
Следовательно, основой органов чувств могут служить универсальные органеллы движения. Специализация и совершенствование этих первичных рецепторов должны привести и приводят в процессе эволюции к высокосовершенным органам чувств животных.
Необходимо отметить, что мысль о универсальности реснитчатого аппарата во всех органах чувств принадлежит не мне. Она возникла в результате обобщения многих весьма тонких морфологических и морфо-физиологических данных о различных рецепторах. Выдающийся вклад здесь принадлежит Я. А. Винникову и его сотрудникам.
Я- А. Винников писал: «Итак, все рецепторные клетки как первично чувствующие у позвоночных, так и первично чувствующие у беспозвоночных, как правило, снабжены особо дифференцированными подвижными антеннами, содержащими в своем составе девять пар периферических и две центральные фибриллы... Фибриллы сокращаются под воздействием АТФ, которая черпается из митохондрий... Таким образом, движение антенн рецепторных клеток ничем не отличается от движения жгутиков и ресничек других одноклеточных или многоклеточных организмов». [51, с. 313].
Следует лишь подчеркнуть, что рецепторы возникли из ресничек при решении в ходе эволюции задач управления движением, перемещением организмов.
«Одиночные каналы». В соответствии с нашим подходом посмотрим, в чем должен состоять предельно совершенный механизм рецепторов?
В самом деле, для рецептора предельное совершенство определяется: 1) предельной чувствительностью; 2) предельной широтой (величиной) диапазона воспринимаемых интенсивностей сигнала и 3) предельной разрешающей способностью различения спектра сигналов. 2-е и 3-е предельные свойства рецепторов достигаются у высших организмов посредством сложного сочетания механизмов и приспособлений, различных для рецепторов, воспринимающих различные сигналы. Мы не можем здесь обсуждать эти проблемы, а ьот предельно высокая чувствительность рецепторов может быть достигнута лишь при эволюции первичных воспринимающих механизмов.
Так, для фоторецептора предельно совершенная чувствительность характеризуется восприятием и реагированием на 1 квант света. Это, в принципе, возможно, если энергия кванта используется лишь для запуска достаточно экзэргонического процесса— типа инициации лавинного перехода из «предварительно заряженного», нестабильного состояния в стабильное. Однако именно так и происходит возбуждение клетки. Сначала создается неравновесное, «напряженное», «инверсное» распределение ионов по обе стороны мембраны. А затем это напряженное состояние «разряжается» — распространяется по мембране волна возбуждения.
Следовательно, один квант света в принципе должен быть способен вызывать лавинную разрядку инверсного распределения ионов в клетке. Для этого нужен микромасштабный рецептор, ключевой механизм. Триггерная, включающая— выключающая система, реагирующая на один квант, может быть лишь молекулярных размеров. Элементарный рецепторный механизм должен представлять собой, например, макромолекулу с двумя конформационным'и состояниями — одно, соответствующее «открыванию», другое — «неоткрыванию» лавинного разряда — запуска волны возбуждения.
Чувствительность такого молекулярного триггера ограничивается тепловым шумом, беспорядочно и напрасно включающего лавинный разряд. Таким образом, триггерная макромолеку- лярная система должна иметь два термодинамически равновероятных состояния, разделенных минимальным кинетическим барьером. Для макроскопической системы эти условия означают область фазового перехода. Мы рассмотрим их подробнее в следующей главе. В механизме возбудимых мембран такие микроскопические, молекулярные триггеры обнаружены несколько лет назад и стали широко известны под названием «одиночные каналы» [383, 399].
При исследовании скачков тока мы впервые получили возможность «визуализации» функционирования отдельных макромолекул. Механизмы функционирования и природа одиночных каналов интенсивно изучаются в настоящее время [5, 158, 172, 383].
Теперь наш гипотетический живой эллипсоид покрыт не просто ресничками, а ресничками, сочетающими в себе свойства органелл движения со свойствами рецепторов химических, световых и механических сигналов. Это странное существо имеет множество глаз, ушей, ноздрей и языков. Ясно, что дальнейшая эволюционная задача — это выбор дискретных зон локализации тех или иных рецепторов. А это связано с проблемой передачи управляющих команд от рецепторов к движителям. Последняя задача вызывает к жизни нервную систему.
Пространственное разделение рецепторов и исполнительных механизмов — двигательных органелл — делает необходимым специализированную связь между ними. Возникает задача преобразования сигналов: специфическая реакция рецепторов на свет, звук и т. п. должна преобразовываться в сигнал, передаваемый по протоплазме к соответствующей ундулоподии. К началу этой стадии эволюции организмы располагают только одним механизмам точного адресования сообщений, а именно, химической специфичностью.
Рецепторы, восприняв специфический сигнал, должны выделять вещество, специфически изменяющее двигательную активность других, не рецепторных ундулоподий. Способ доставки такого вещества адресату — диффузия. Гарантией правильности адреса служит химическая специфичность: адресат сам выбирает вещество, являющееся носителем информации о возбужденном состоянии рецептора. Диффузия в пределах клетки, т. е. организма с размерами порядка сотен микрон, обеспечивает довольно высокую скорость передачи сигнала; промежуток времени между срабатыванием рецептора и поступлением вещества к ресничке порядка секунды.
Однако по уже рассмотренным выше причинам диффузия препятствует строгой направленности сигналов.Ресничек много, все они одинаковые, но расположены на разных участках поверхности нашего эллипсоида. Поэтому при выделении рецептором регуляторного вещества биения всех ресничек будут усиливаться или ослабляться.
Это затруднение можно преодолеть посредством специализации рецепторов ресничек 'И медиаторов или ;при помощи труб.! Нужно проложить трубы от рецепторов к регулируемым ими рес-} ничкам, и тогда регуляторное вещество будет диффундировать в строго заданном направлении. Морфогенная проблема создания полых цилиндров — труб решена в эволюции раньше. Организму достаточно иметь необходимый набор макромолекул белков и трубы нужного диаметра самособерутся из этих морфологических единиц (см. выше, с. 148). Здесь возникает задача «правильной» укладки труб, а именно соединения ими строго определенных ресничек и рецепторов. Как решается эта задача внутри одной клетки — я не знаю. Однако сам факт наличия внутри клеток большого числа микротрубочек установлен и очень занимает внимание цитоморфологов в последнее время. Существование таких трубочек внутри клетки, как мне кажется, обеспечивает необходимую точность адресования сигналов, строго направленную диффузию специфических химических веществ- регуляторов *.
Пытаясь выяснить возможную химическую природу этого вещества-регулятора, мы должны будем использовать уже полученные .нами сведения о механизмах регулирования биений ундулоподий '. Вряд ли в качестве регулятора можно использовать АТФ-рецептор сам потребляет чужую АТФ, поставляемую специализированными биохимическими системами, но им может быть какой-либо регулятор АТФ-азной активности. Опираясь на знания биохимии современных организмов, мы должны были бы поручить эту роль ионам кальция. Диффузия по трубке ионов кальция или связывающих их веществ могла бы служить хорошим способом управления биения ресничек. Правдоподобность такого способа регуляции двигательной активности следует из многих данных о работе различных аппаратов движения. Именно так регулируется сокращение стебелька сувойки. Изменение концентрации ионов кальция регулирует работу мышц всех видов — недаром роли кальция в биологических процессах посвящаются специальные книги (см., например, [64]). Примем пока, что внутри одной клетки регуляторные импульсы от рецепторов передаются двигательным органеллам по специальным трубам в виде порций ионое -кальция или веществ, 'изменяющих состояние кальция в ундулоподиях.
Теперь наш реснитчатый организм выглядит следующим образом. Основные рецепторы специализированы по виду принимаемых ими сигналов и расположены на том конце эллипсоида, который направлен ,в сторону своего движения. Это головной конец эллипсоида, противоположный его конец — хвост. Ундуло- подии покрывают поверхность организма не равномерно, а так, чтобы обеспечить наиболее рациональное их движение в нуж- юм направлении. Рецепторы связаны с ундулоподиями посредством трубочек, передающих химические сигналы.
По-видимому, дальнейшее совершенствование такого одноклеточного организма может идти по пути все большей дифференцированное™ рецепторов, все более рационального их расположения, группировки, взаимодействия, все более совершенной связи с ундулоподиями. Однако никакого существенного увеличения в скорости перемещения к цели этим уже не достичь. Она лимитируется предельной, невысокой скоростью движения организмов посредством ресничек. Управление же движением, совершаемым посредством мышц, требует решения совсем других задач. Как мы уже показали выше, движение при помощи мышц состоит в изменении формы клетки ,в целом, и поэтому направленное перемещение организма может быть осуществлено лишь в .многоклеточном организме. В конце предыдущей главы уже отмечалось, что само возникновение многоклеточных организмов, пс-видимому, обусловлено 'Именно решением в процессе эволюции задачи резкого ускорения управляемого направленного движения. Мы пришли к выводу об обусловленности вознмшовения многоклеточности .именно необходимостью все болышх скоростей перемещения в пространстве. В многоклеточном организме каждая отдельная -мышечная клетка аналогична по своей функции ундулоподии одноклеточного организма. Только совокупность мышечных клеток может обеспечить движение организма в разных направлениях. Управление перемещением такого организма требует решения почти тех же задач, что и в случае реснитчатого эллипсоида. Необходимо, 1Тобы сигналы от рецепторов, сгруппированных в определенны: зонах поверхности тела, поступали в -мышечные клетки и регуляровали частоту и силу их спонтанного ритмического сокращенш (Мы предполагаем, что спонтанное ритмическое сокращение является первичным свойством первичных мышц).
Однако теперь проблема передачи сигналов усложняется. Речь идет 1еперь об их передаче не внутри одной клетки, а от одной клетки к другой. Здесь уже эволюции приходится думать о передаче сигналов через поверхностную мембрану клеток или посредством нее. Теперь нужны морфологические связи, быть может, такие же трубы между клетками. Однако трубы с диффундирующими в них сигнальными химическими веществами уже непригодны: скорость диффузии и в данном случае оказывается узким местом — она -совершенно недостаточна для управления движением мышечных клеток. Нужно время порядка 1 — 10 сек для передачи сигнала диффузионным путем от клетки к клетке .при характерном размере клетки 10~2 см.
Во-первых, резко -возрастают скорости движения, и поэтому управление перемещением должно происходить значительно быстрее. Во-вторых, резко возрастают расстояния, на которые приходится передавать сигналы. Следовательно, порции веществ, диффундирующих по трубам, уже не могут служить сигналами. Нужны другие механизмы распространения сигналов от рецепторов к двигательным клеткам.
Что кроме веществ может распространяться по трубчатым или иным соединениям клеток друг с другом? Химические процессы (перемещается фронт горения или фронт колебательной химической реакции), фронт структурной перестройки (например, фронт кристаллизации или плавления), электрические, акустические сигналы и, наконец, световые импульсы.
Распространение в трубах химических процессов или фронтов фазовых переходов (структурных перестроек) требует заполнения трубы активным веществом и обязательной регенерации его активного состояния после прохождения каждого импульса— это должен быть бикфордов шнур, снова образующийся после каждого взрыва. Распространение электрических, акустических и электромагнитных (световых) импульсов не связано,в принципе, с необходимостью регенерации самого волновода после прохождения каждого импульса.
Итак, речь идет о связи рецепторов и мышечных «леток посредством «нервов».
После работ Ходжкина, Хаксли и других выдающихся исследователей (см. [130, 313, 314]) природу проведения нервного импульса принято считать выясненной. Известно, что возбуждение по нерву распространяется в виде волны деполяризации — волны изменения распределения ионов натрия и калия по обе стороны клеточной мембраны аксона. Об этой волне можно судить по изменению электрического потенциала. Физико-химиче- ским механизмом распространения возбуждения по нерву вероятно служит процесс (волна) конформационных перестроек вещества мембраны. Одна из первых попыток регистрации распространения по нерву волны структурных 'изменений принадлежит Г. М. Франку, Р. Г. Людковской, Л.П. Каюшину, изучавшим возможность регистрации прохождения механических волн по возбуждаемому нерву посредством интерференционного микроскопа [131а, 1866]). В последующие годы исследованиям физических процессов такого рода, проведенным главным образом оптическими методами, было посвящено много работ [22, 365, 410]. В детально разработанном виде концепция, связывающая распространение возбуждения по нерву с волной конформационных изменений, была создана Тасами [281]. Тасаки полагал, что возбуждение мембраны представляет собой быстрый обратимый катионообменный процесс, сопровождающийся переходами между двумя устойчивыми (конформационными) состояниями образующих ее макромолекул... Конформация мембранных макромолекул и свойства катионообменника в основном определяются отношением концентраций одновалентных и двухвалентных катионов в мембране... Причиной распространения процесса возбуждения по мембране аксона служит различие макромолекулярных конформаций в возбужденном состоянии и в состоянии покоя... В конце концов, если начальная возбужденная поверхность была достаточно велика, вся мембрана переходит из устойчивой конформации покоя в устойчивую возбужденную конформацию.
Такое представление о механизме проведения нервного импульса в общем вполне правдоподобно. Оно может быть модифицировано с учетом новых данных о существовании и свойствах одиночных каналов (см. выше) и фазовых переходах в фосфолипидах мембран (см. следующую главу). Но главный критерий естественного отбора, определявший направление эволюционного становления механизмов возбуждения по нерву это скорость распространения нервного импульса.
Правдоподобным может быть лишь механизм, обеспечивающий предельновозможную, ограниченную лишь физико-химическими свойствам-и системы, скорость проведения нервногоимпульса. Скорость распространения нервного импульса .впервые измерил (в 1850 г.) Гельмгольц и получил величину около 30 м/сек. Теперь мы знаем, что скорость распространения нервного импульса обычно близка к этой величине, но в разных нервах может варьировать от 0,1 до 150 м;сек. [358]. Какие факторы ее ограничивают? Если верна гипотеза о распространении волны структурной перестройки и сопряженной с пей вол,ны изменения объема, т. е. акустической волны, то скорость распространения нервного импульса должна зависеть от скорости структурной перестройки (фазового перехода) и от предельно возможной скорости звука в нервном волокне. Здесь мы еще раз должны обратить внимание ,на малую скорость звука в гелях [253] —скорость нервного импульса, по-видимому, неслучайно совпадает по порядку величин со скоростью звука в гелях[17]. Итак, скорость звука при данных значениях модулей упругости и геометрии проводящих структур ограничивает скорость распространения волны конформационных переходов макромолекулярных комплексов мембраны при распространении возбуждения по нерву. Подробнее мы рассмотрим это обстоятельство ниже. Пока же необходимо сказать, что, если распространение волны возбуждения по нерву основано на обратимом фазовом переходе, то оно возможно лишь при условии практической равновероятности двух состояний, т. е. в узкой зоне фазового перехода (см. следующую главу).
Можно все же, не боясь показаться наивным, спросить: а почему бы не снять этот предел, создав посредством естественного отбора иной механизм проведения нервного импульса, например механизм, основанный на распространении электромагнитных волн или прямом проведении электрических сигналов?
Электромагнитные сигналы, видимый свет и нервный импульс. Для управления перемещением необходимы устройства, обеспечивающие строгое безошибочное адресование сигнала. Для этого вполне пригоден «видимый» свет. В случае световых сигналов необходимы источники излучения, а также световоды — нити с такими оптическими свойствами, с такими показателями преломления, чтобы свет распространялся лишь внутри нити, испытывая полное внутреннее отражение на границе световод — внешняя среда. Поглощение света в световоде, в его веществе должно быть небольшим, а на конце световода должно находиться вещество, обладающее высокой фотохимической активностью.
Я не понимаю почему бы, в самом деле, в процессе эволюции не могла возникнуть такая связь рецептора и двигательного аппарата? Источники интенсивного излучения видимого света биохимически реальны (см. [69]). Известно множество различных биохимических механизмов изучения света: от свечения бактерий в результате люцеферазной (окислительной) реакции до свечения белка, выделенного из гребневиков и медуз, светящих- <ся при взаимодействии с ионами кальция [164, 200]. Имеются также данные о влия?ши света на биохимические процессы (см. [437]). Зато какая скорость проведения'Сигналов — скорость света! Насколько можно судить, нервные волокна обладают всеми необходимыми свойствами световодов, и поэтому возможность передачи световых сигналов по нервам не кажется очень фантастической Я думаю, что вполне реален эксперимент для проверки возможной роли видимого света в передаче сигналов по нервам. Электромагнитные волны, соответствующие другим частотным диапазонам, как указывалось выше, по-видимому, не пригодны. Более короткие — в силу жесткости, повреждающегося действия, более длинные — из-за очень сильного поглощения в среде, воде, протоплазме.
Таким образом, в соответствии с современными биохимическими и биофизическими представлениями нельзя считать невозможной связь клеток посредством излучений .в видимой или ультрафиолетовой области электромагнитного спектрй.
Было бы непростительной ошибкой вслед за М. В. Волькенш- тейном a priori квалифицировать такую возможность как «лженауку»[18]. Весь вопрос в получении достоверных фактических данных. Следует заметить, что огромный фактический материал А. Г. Гурвича и его сотрудников [77, 78] по такому влиянию отнюдь не был опровергнут. Его работы не были продолжены ■отчасти в силу трудностей в «объективной» регистрации «мито- генетического» излучения, отчасти по психологическим и другим причинам..
В недавнее время появились сообщения В. П. Казначеева н ■сотр. [123] о взаимовлиянии клеток посредством излучений оптического диапазона. Эти данные заслуживают внимания и экспериментальной проверки.
Наконец, совсем недавно опубликованы результаты чрезвычайно интересных опытов В. А. Мостовникова и И. В. Хохлова 1218] также обнаруживших сильное, специфическое взаимодействие клеток в культуре ткани, посредством излучаемого или видимого света[19].
Можно надеяться, что в близком будущем количество экспериментальных данных станет достаточным для определенных выводов.
Существует хорошо известный способ передачи сообщений посредством телеграфа. Почему же эволюция не использовала этот принцип? В самом деле, для чего эволюция создала столь ■сложные механизмы проведения нервного импульса — посредством волны деполяризации, волны изменения проницаемости, волны конформационной перестройки? Мало того, что они сложны,они еще и не могут обеспечить большую скорость. Ответ очень прост — прямое прохождение электрического сигнала по нерву невозможно в силу непреодолимых биохимических препятствий; нервы можно было создать в ходе эволюции лишь биохимическим путем, т. е. из водных растворов солей, белков и т. д., заключенных в фосфолипидную оболочку. Сопротивление даже самого электропроводного раствора соли на 7—8 порядков (!) больше сопротивления металла. Поэтому телеграфный принцип передачи сообщений, а именно, передача низковольтного, слабого импульса тока по проводу от источника к приемнику здесь не годится. Не годится по нескольким причинам.
Одна из них — невозможность создания импульса с достаточно высоким напряжением, способного пробиться по жидкостному проводнику — трубе с протоплазмой. Дело не в технической неосуществимости высоковольтного разряда — биофизические батареи электрических рыб показывают, что в принципе это возможно. Разности электрических потенциалов, превышающие 0,4 в, уже не годятся из-за электрохимических эффектов — электрохимического восстановления кислорода и других веществ, а затем при несколько большей разности потенциалов и из-за электролиза воды.
Будучи жестко ограничены верхним пределом разности потенциалов, мы сталкиваемся с невозможностью уменьшить в должной степени сопротивления и путем увеличения толщины проводника. Удельное сопротивление протоплазмы порядка 100 ом/см. Следовательно, протоплазматический проводник диаметром порядка 10 мк будет иметь удельное сопротивление порядка 100 Мом/см. Поскольку по аналогии с телеграфом сопротивление принимающего аппарата должно быть значительно больше сопротивления провода, в клетке должны были бы содержаться вещества с сопротивлением слоя толщиной в несколько сот А порядка 108—1010 ом, т. е. вещества, являющиеся очень хорошим изоляторам.
Таким образом, прямое проведение сигнального электрического импульса по нервному волокну с диаметром клеточного масштаба невозможно. Можно лишь пожалеть об этом; ведь по телеграфу сообщения передаются очень быстро.
Значит (пока вопрос о нервах-световодах не решен) мы можем рассматривать лишь одну возможность, а именно, проведение сигналов по нерву посредством волны структурной перестройки. Ясно, что и здесь под давлением искусственного отбора рано или поздно должно достигаться предельное совершенство, т. е. предельно возможная скорость проведения незатухающего сигнала по нерву.
Скорость проведения у позвоночных животных оказывается достаточно высокой вследствие миелинизации нервного волокна с перехватами Ранвье через каждые 1 —2 мм, а у беспозвоночных животных, нуждающихся в возможно более быстром прове-
дении нервного импульса,— вследствие образования предельно толстых (до 1 мм) нервных волокон.
Как отмечалось, в механизме проведения сигналов по нерву посредством волны деполяризации источником необходимой энергии служит неравновесное состояние градиентов концентрации катионов— калия, натрия, кальция. Скорость проведения нервного импульса зависит от скорости изменения этих градиентов, т. е. от быстроты конформационной перестройки мембранных компонентов. По мнению Катца [130], «скорость распространения волны деполяризации зависит от продольной электропроводности внутренней области аксона и поэтому она тесно коррелирует с толщиной волокна. Как известно, один из способов увеличения скорости передачи сигналов состоит в уменьшении осевого сопротивления кабеля путем увеличения диаметра волокна. Именно это решение избрала природа для удовлетворения потребности в высокоскоростной передаче импульсов у некоторых беспозвоночных. Например, быстрое движение кальмара при бегстве от опасности контролируется небольшим числом гигантских аксонов, иннервирующих обширную мускулатуру его мантии, которая действует как реактивный двигатель. Однако для животного, которому помимо быстроты реакции необходимо и наличие огромного числа каналов для передачи множества сенсорных сообщений и обеспечения двигательных реакций, гигантские аксоны оказались бы непригодными. Ясно, например, что в зрительном нерве, где должно находиться больше миллиона параллельных аксонов для передачи зрительной информации просто не хватило бы места для большого числа гигантских волокон. Решением, найденным нервной системой позвоночных, явилось образование миелинизиро- ванного аксона, в котором кабельные потери сильно снижены благодаря миелиновой оболочке» (130, с. 114).
Поскольку механизм проведения нервного импульса рассмотрен в трех книгах выдающихся исследователей этого процесса — Тасаки [281], Катца [130], а также в классической книге А. Ходжкина [313], я не буду излагать сведения о механизме генерации и проведения нервного импульса. Сказанное выше можно просуммировать следующим образом.
Наиболее совершенный способ связи рецептора и двигательного аппарата, которые обязательно должны быть пространственно разделены, состоит в передаче сигналов по нервному волокну. Скорость передачи сигналов по нерву (нервных импульсов), осуществляемых по принципу структурных перестроек, не может превысить скорость звука и фактически лимитируется подвижностью ионов в электрическом поле. Возможность передачи световых сигналов по нервному волокну не исследована.
Нервы возникли в эволюции вследствие необходимости преодоления пространственной разобщенности механизма, воспринимающего сигналы из внешней среды, и механизма перемещения в пространстве; сни необходимы, кроме того, из-за медленностии беспорядочности направлений диффузий химического регулятора. Однако непосредственным способом управления биохимическим механизмом перемещения в пространстве может стать только образование в мышечной клетке специфических веществ. Только такие вещества способны изменить скорость ферментативного процесса расщепления АТФ, инициировать процесс конформационных изменений макромолекул, приводящих к преобразованию свободной энергии химического процесса, в механическую работу.
Следовательно, нервный импульс, дойдя до цели, до конца нервного волокна, должен вызвать образование специфического химического регулятора сокращения мышцы (что и происходит на самом деле). Сейчас мы знаем, что роль такого медиатора играют ионы кальция. Освобождение ионов из связанного состояния (в структурах саркоплазматического ретикулума) и соединение его с белками комплекса актомиозин — тропомио- зин — тропонин — условие начала сокращения миофибриллы, начала движения нитей актина и миозина навстречу друг другу. Связывание кальция служит причиной прекращения ферментативного расщепления АТФ, прекращения энергетического обеспечения сокращения миофибрилл, т. е. условием расслабления, сопровождающегося при нагрузке растяжения миофибриллы, например, при действии эластических сил коллагеновых волокон или груза, или же под действием реципрокяых (тянущих в противоположную сторону) мышц. Однако количество ионов кальция, непосредственно поступающих в протоплазму в результате прихода нервного импульса, очень невелико, и .поэтому на нервных окончаниях действует механизм химического усиления, т. е. увеличения количества кальция, происходящего посредством медиаторов. Под влиянием нервного импульса выделяется химический медиатор — ацетилхолин (обеспечивающий регуляцию быстрых мышц) или адреналин (регулирующий относительно длительный тонус специализированных мышц в стенках кровеносных сосудов). Эти медиаторы запускают процессы, приводящие к появлению больших количеств кальция в иннервируемом органе.
Эволюционное совершенствование связей двигательных аппаратов (мышц) с рецепторами посредством нервов является залогом дальнейшего прогресса в решении основной проблемы эволюции на данном этапе — проблемы совершенного управления перемещением организма. Мы уже видели, что управление движением становится предельно совершенным лишь при достижении совершенного анализа траектории произвольного движущегося тела (жертвы или хищника). Анализ произвольной траектории и управление движением собственного организма основаны на функциях восприятия, запоминания, обобщения, сопоставления образов, в частности абстрактного образа — модели траектории с реально осуществляющейся траекторией.
Все это должно завершиться возникновением управляющей сигнальной системы, определяющей движение организма в нужном направлении по соответственно построенной траектории. Выполнение таких сложных задач не обеспечивается элементарной связью рецептора и двигательного аппарата — простейшей |рефлекторной дугой. Непосредственная связь рецепторов с двигательным аппаратом становится принципиально недостаточной для управления движением в случае взаимодействия двух движущихся организмов. Это объясняется тем, что абсолютные скорости двух взаимодействующих организмов в принципе могут быть одинаковыми. И тогда хищник никогда не догонит жертву: они будут двигаться с равными скоростями, сохраняя постоянный интервал. И, наоборот, даже небольшое преимущество в скорости у хищника лишит жертву всех надежд на спасение. При близости скоростей перемещения в пространстве взаимодействие в системе жертва — хищник возможно лишь при произвольных траекториях движений. Это значит, что управление перемещением посредством простых рефлекторных механизмов не пригодно. Хищник не должен бежать непосредственно к тому месту, где он увидел жертву. Пока он добежит, жертвы там уже не будет. (Зенитчики стреляют, целясь не в летящий самолет, а вынося прицел далеко вперед, т. е. целясь ,в то место, куда при правильной экстраполяции самолет должен прилететь в момент долета туда снаряда). Итак, нужен анализ воспринятых рецепторами сигналов, необходима предварительная переработка информации и лишь затем должны возникать сигналы управления движением.
Глава 11
ТЕПЛОКРОВНОСТЬ И ТЕПЛОУСТОЙЧИВОСТЬ
Теплокровность, повышение температуры тела относительно температуры среды не связаны непосредственно со скоростью биохимических процессов.
Теплокровность необходима для высокосовершенного функционирования нервной системы.
Необходимо соответствие теплоустойчивости белков температуре среды обитания.
Температура среды — важнейший физический фактор эволюции. Приспособление к той или иной температуре среды, т. е. выработка механизмов терморегуляции, играет чрезвычайно важную роль при определении кинетического (биологического) совершенства живых организмов. Есть два способа приспособлений организмов к разной температуре среды.
Первый способ состоит в поддержании постоянных скоростей биохимических и физиологических процессов посредством компенсирующей регуляции этих скоростей при разных температурах ■среды обитания, а следовательно, и тела организмов (пойкило- термные организмы). Второй способ состоит в поддержании постоянной температуры тела при разных температурах среды (гомойотермные организмы). Тем самым он обеспечивает независимость существования организма от температуры среды.
Априори совсем не очевидно, что второй способ «лучше» первого. Более того, лично мне кажется более «удобной» пойкило- термия —• она не требует столь тонких и сложных механизмов терморегуляции. Нам, людям («голым обезьянам») приходится тратить огромное количество угля, нефти, газа, электроэнергии на согревание жилищ. А многие пойкилотермные организмы, например океанические рыбы, ведут весьма активную жизнь при температуре тела, близкой к 0°. Более того, если теплокровность по каким-то причинам (например, для обеспечения автономии от колебаний условий среды) и оказалась выгодной, то почему же наиболее высокоорганизованные животные — млекопитающие и птицы — поддерживают температуру своего тела в узком диапазоне вблизи 40°? Ведь здесь дорог каждый градус — куда экономнее было бы иметь температуру 25 или 30°.
Нужно сказать, что хотя проблемам терморегуляции посвящены капитальные исследования [117, 134, 151, 273, 300, 330, -364], вопрос о физико-химических причинах возникновения в эволюции теплокровных организмов и выбора температуры тела вблизи 40°, как мне кажется, не анализируется с должной строгостью.
При анализе этих вопросов сразу же становится очевидным, что необходимость повышения температуры тела по сравнениюсо -средой -возникает в эволюции не сразу, а лишь после преодоления диффузионного ба-рьера. В самом деле, скорость химических реакций увеличивается с ростом температуры значительно быстрее скорости диффузии. В самом деле, ихдл ~ ехр {—Е^/кТ), tW~y7\ Поэтому для систем, неприспособленных к активному преодолению диффузионного барьера, повышение температуры скорее вредно, чем полезно: ухудшается возможность регулирования скоростей разных химических реакций, и с приближением к диффузионному пределу он.и уравниваются. Итак, первый вывод: увеличение температуры способствует кинетическому совершенству лишь активно перемещающихся животных. Как известно, £акт ферментативных реакций мало, т. е. скорость ферментативных процессов слабо зависит от температуры, однако активность ферментов сильно зависит от других факторов—присутствия активаторов, особенностей аминокислотного состава макромолекулы и т. п. Это заставляет усомниться в правильности распространенного мнения о повышении температуры тела, как непосредственном способе ускорения биохимических процессов. Таков второй вывод. Наконец, принято думать, что при высоких температурах возможность жизни высокоорганизованных животных ограничена в -силу все -возрастающей с повышением температуры денатурации белков. Это также весьма спорно — большинство белков и нуклеиновых кислот денатурируют при температурах, значительно превышающих 36—40°. Кроме того, молекулы белков и нуклеиновых кислот могут в процессе естественного отбора приобрести относительно высокую теплоустойчивость (термофильные бактерии). Значит, возможность жизни сложных организмов при высоких температурах ограничена не денатурацией макромолекул, а другими причинами. Таков наш третий вывод.
Итак, хорошо было бы попытаться выяснить, почему для наиболее высоко организованных животных необходима температура тела в диапазоне 36—40°.
Узость температурного диапазона активной жизнедеятельности теплокровных более всего напоминает ограниченность температурных интервалов, соответствующих каким-либо фазовым переходам. Более того, узость зоны теплокровности свидетельствует о том, что для активной жизнедеятельности необходимы условия, соответствующие именно переходу одного состояния в другое, а не преимущественному нахождению в каком-либо одном состоянии. О каких фазовых переходах может здесь идти речь?
Вот один из возможных ответов на поставленный вопрос: речь идет о переходе гель — золь в протоплазме. В самом деле, мы уже пришли к выводу, что протоплазма должна быть жидкой, чтобы не затруднять движение в клетке метаболитов и макромолекул. Однако для активного перемещения, для движения протоплазмы^ для поддержания и изменения формы клеток их содержимое должно быть гелеобразным. Тогда «наилучшей» будет жизнь в условиях, соответствующих переходу гель — золь. О каких золях игелях можно говорить в этом случае? Как известно, гели делятся на экзо-, эндо- п изотермические [405]. Экзотермические гели, возникающие главным образом вследствие взаимодействия полярных групп, плавятся при нагревании (желатина, агар). Эндотермические гели, образующиеся по преимуществу вследствие гидрофобных взаимодействий, плавятся при охлаждении (акто- миозиновые гели); изотермические гели не изменяют своего агрегатного состояния при изменении температуры. Следовательно, нагревание способствует ускорению внутриклеточных биохимических процессов лишь в том случае, если внутриклеточные структуры образуют экзотермический гель. Вполне вероятно, что в клетке преобладает именно такой гель — трехмерная сеть типа фибриллярных коллагеновых молекул. Превращение этого геля в золь — плавление геля, способствует резкому усилению перемешивания внутриклеточного содержимого, ускорению биохимических процессов. Однако в большинстве клеток присутствуют и белки актомиозинового комплекса, образующие эндотермический гель. Впрочем, последний особенно эффективно плавится не при охлаждении, а при действии АТФ.
Мысль о том, что переход гель — золь служит универсальным регулятором общей скорости биохимических процессов, была чрезвычайно популярна уже более 30 лет назад. Потом в связи с грандиозными успехами биохимии, выявлением -и изучением множества отдельных ферментативных реакций и их систем взгляд на протоплазму как .на единую коллоидную систему был не то, чтобы скомпрометирован, а просто стал казаться .наивным. Тем не менее, вполне соответствующим совокупности современных данных можно считать предположение о ключевой .роли в клетке коллагеноеого или, быть может, тубулянового геля. Белки такого типа необходимы для придания клетке специфической формы. Упорядоченные сети, структуры из волокон составляют каркас, поддерживающий оболочки животных клеток. Ясно, что движение протоплазмы, перемешивание внутриклеточного содержимовозможны лишь .при полужидком состоянии, т. е. в диапазоне температур перехода гель — золь. Более того, течение внутриклеточного содержимого может осуществляться лишь при подвижной оболочке клеток. Следовательно, и оболочка должна быть полужидкой. Итак, мы как будто бы пришли к .выводу, что температурные оптимумы определяются температурами переходов гель — золь коллагенов или иных образующих экзотермический гель компонентов клетки.
Увы, это так и не так. Исследования и обобщения ряда авторов [96, 97, 166] показали, что справедливо обратное соотношение, а именно, температуры структурных переходов коллагенов соответствуют температурам среды обитания клеток и определяются вариациями аминокислотного состава коллагенов. Иными словами, физико-химические свойства коллагенов «приспосабливаются» в ходе эволюции к разным температурным условиям, а не наоборот. Эволюции не приходится подстраиваться под прихоти коллагенов.
Более вероятно, что температура тела, необходимая для гомой- отермных животных, определяетеяпаличием липидов возбудимых мембран клеток. Мысль об ответственности липидов за температурную адаптацию животных, была высказана Л. Гейльбруном 1396], а затем ее развивали многие исследователи (см., например, [364] ). Доводом в пользу такой гипотезы служило соответствие температуры плавления жиров организма температуре среды обитания или (у гомойотермных животных) температуре тела. Как известно, температура плавления жиров тем ниже, чем больше в их составе ненасыщенных жирных кислот. Количество ненасыщенных связей измеряют количеством иода, реагирующего с этими связями (так называемое йодное число). Корреляция йодного числа жиров с температурой обитания организма обычно очень хороша, причем интересно, что эта корреляция прослеживается для теплоустойчивости организма как целого, а не для теплоустойчивости отдельных клеток. Последнее обстоятельство, по мнению Б. П. Ушакова [300], не свидетельствует в пользу липидной теории теплоустойчивости клеток. Относительно клеток он, возможно, прав. Но нас-то интересует организм — живое, многоклеточное существо, объединенное в единое целое нервной системой.
Есть и другие основания для недовольства липидной теорией в ее классическом виде. В соответствии с ней температурные характеристики липидов (как и коллагенов) организма легко адаптируются к температуре среды обитания вида. А мы ищем инвариант — непреодолимые по физико-химическим соображениям обстоятельства, делающие температурный интервал 36—40° универсальной оптимальной зоной условий существования наиболее совершенных животных. Такой системой, свойства которой не подгоняются к температуре существования, а, в известном смысле, определяют ее, являются, по моему мнению, не вообще липиды, а липопротеидные комплексы возбудимых мембран. Состав и свойства предельно совершенных возбудимых мембран клетки жестко детерминированы их главными функциями — способностью к созданию асимметричного распределения ионов и к проведению волны возбуждения. Волны структурной перестройки, волны конформационных изменений в мембране, обеспечивающие проведение возбуждения, с наибольшей легкостью осуществляются именно в зоне равной вероятности двух состояний, т. е. в зоне фазового перехода. Следовательно, скорость распространения возбуждения будет наибольшей при температурах и при других условиях., соответствующих зоне фазового перехода. Эта скорость резко падает при температурах, меньших и больших оптимальной.
Итак, возможный ответ на вопрос о причинах выбора в ходе эволюции температурного оптимума жизни животных формулируется следующим образом: область 36—40° соответствует диапазону температур фазового перехода возбудимых мембран[20]. Свойства мембран определяются их основной функцией—возбудимостью. Эволюция всех биохимических процессов и систем в возбудимых клетках направлена на совершенное обеспечение этой основной функции. Из сказанного следует, что у холоднокровных скорость проведения нервного импульса должна быть, как правило, существенно меньше, чем у теплокровных (при температуре тела, меньшей 36—40°).
Для подтверждения правильности предположения об обусловленности температурного режима многоклеточных животных физико-химическими свойствами липопротеидных комплексов их возбудимых мембран можно привести разные факты. Так, в работах В. Б. Ушакова [301] было показано, что при тепловой инактивации мышц лягушки сначала инактивируются компоненты возбудимой системы. Мышечное волокно в целом гибнет до того, как наступают заметныефизико-химические изменения сократительных белков. В работе Т. А. Джамусовой [88] показано, что необратимая потеря возбудимости при нагревании мышц травяной лягушки начинается при 36° и происходит с заметной скоростью. Завершается она при 42°. Эти температуры отнюдь не соответствуют температуре среды обитания лягушек, а являются, по-видимому, свидетельством инвариантных свойств возбудимой мембраны, определяемых зоной фазового перехода ее липопроте- идного комплекса. У теплокровных мембраны работают в условиях, максимально способствующих появлению волны конформационных перестроек.
Как мы видим, предполагаемое предельное совершенство клеточной мембраны именно как возбудимой системы детерминирует определенный температурный режим существования соответственно совершенных животных. Итак, фактором, определяющим эволюцию в направлении теплокровности, является скорость распространения волны возбуждения по нерву, а не скорость ферментативных реакций. Теплокровные победили холоднокровных благодаря большим скоростям реакций, большим скоростям выбора траекторий движения в ходе преследования жертвы или бегства от хищников. Это становится ясным при наблюдении, например, за рептилиями. Скорость реагирования и перемещения змей и ящериц столь явно изменяется с температурой, что более строгие доказательства кажутся излишними.
Итак, время, затрачиваемое на переработку сигналов от рецепторов и проведение сигналов, является критерием в естественном отборе, приводящим к появлению теплокровности. Конечно же, на самом деле все может быть существенно сложнее. И здесь возможны удивительно тонкие «обходные маневры». Так, холоднокровные лягушки никогда бы не успевали поймать муху, еслибы их сетчатка не была устроена особым образом. Восприятие сетчаткой круглого темного движущегося предмета с угловыми размерами мухи или небольшого жука вызывает автоматическую реакцию захвата добычи посредством прямой, почти не заходящей в головной мозг, связи сетчатки с соответствующей группой эффекторов.
Однако платой за скорость в данном случае оказывается отсутствие центрального аппарата анализа, а следовательно, и невозможность эволюционного развития мозга в направлении способности к экстраполяции траектории быстро перемещающихся объектов *.
У теплокровных сетчатка не замкнута накоротко на исполнительные органы, и существует мощнейший центральный аппарат анализа зрительных образов. Это становится возможным именно в результате теплокровности.
После того, как было написано изложенное выше, мне стали .известны факты, указывающие на большую вероятность предположения об обусловленности диапазона 36—40° у теплокровных фазовым переходам в фосфолипидах возбудимых мембран.
Относительно недавно было установлено, что в системе фосфолипиды— вода и в самом деле наблюдается резкий фазовый переход в узком диапазоне невысоких температур (см. литературу к статье [443]). Этот переход состоит в превращении кристаллической структуры в жидко-кристаллическую и осуществляется как в искусственных бислойных фосфолипидных мембранах, так и в естественных мембранах. Температура фазового перехода зависит в основном от структуры цепей жирных кислот — для жирных кислот с одной двойной связью и транс-конфигурацией температура перехода значительно выше, чем для таких же кислот с цис-ко.нфигурацией.
О биологическом значении фазовых переходов свидетельствуют и данные работы [443], выполненной на мутантах Е. coli, не способных самостоятельно синтезировать жирные кислоты с длинной цепью. Мутантов выращивали на средах, содержащих разные жирные кислоты, используемые этими микроорганизмами для построения фосфолипидов клеточных мембран.
Оказалось, что при добавлении в культуральную среду жирной кислоты, состоящей из 18 атомов С и находящейся в трансконфигурации относительно двойной связи углеродного атома в положении 9 (транс — Д9С18:1), скорость переноса веществ через мембрану резко увеличивалась при температуре, превышающей 37°. При добавлении в среду той же кислоты в цис-кон- фигурации скорость переноса резко возрастала при 16°, при добавлении транс — A9C1S:1 — при 31°.
Анализ различными физическими методами выделенных из клеток фосфолипидов, клеточных мембран, а также целых клеток показал, что температуры, соответствующие резкому изменению скорости трансмембранного переноса, лежат вблизи температур фазового перехода кристалл — жидкий кристалл для соответствующих препаратов фосфолипидов (в основном— фосфатидилэтаноламина) [422]. При температурах, меньших температуры перехода, мембраны состоят из молекул липидов, упакованных в гексагональную кристаллическую решетку. В такие мембраны утоплены молекулы белков-перенос- чиков, и транспорт через них весьма затруднителен. При температуре фазового перехода происходит резкое увеличение подвижности углеводородных цепей, мембрана становится жидкой, трансмембранная диффузия и активный перенос веществ оказываются облегченными (см. в частности [143]).
Из сказанного выше следует, что в возбудимых мембранах теплокровных наиболее существенную роль играют фосфолипиды, содержащие транс — AeC,8;i — жирные кислоты. В зоне фазового перехода этих фосфолипидов при температурах около 38° становится возможным распространение волны возбуждения по нерву. Итак, весьма вероятно, что наиболее совершенные го- мойотермные животные поддерживают температуру тела около 38°, поскольку именно при этой температуре обеспечиваются наилучшие условия функционирования возбудимых мембран.
КОРРЕЛЯЦИЯ ТЕПЛОУСТОЙЧИВОСТИ БЕЛКОВ С ТЕМПЕРАТУРОЙ СРЕДЫ ОБИТАНИЯ ПОИКИЛОТЕРМНЫХ животных
Итак, предельно совершенные, активно перемещающиеся животные должны быть теплокровными, поскольку именно в диапазоне температур 36—42° обеспечивается максимальная скорость проведения возбуждения по нерву. Этот диапазон температур задан свойствами фосфолипидов возбудимых мембран. Сам фосфолипидный состав детерминирован необходимостью максимально совершенного выполнения функций возбуждения и проведения возбуждения. Приходится идти на заботы о высокосовершенной системе терморегуляции, поддерживающей необходимую температуру тела.
Как живут холоднокровные, пойкилотермные животные и растения?
Существует удивительное явление — корреляция термоустойчивости (температуры денатурации) внутриклеточных белков и средней температуры среды обитания животных. Большой вклад в эти проблемы и обстоятельные обзоры этого феномена осуществили В. Я- Александров [6, 7] и Б. П. Ушаков [300] (см. также [465]). Что удивительно: коррелирует температура денатурации белков, соответствующая диапазону температур 50—100°, и температура среды обитания, соответствующая, например, температурам 15—25°. В силу наших причин ферменты типа миокиназы денатурируют у озерной лягушки (средняя температура среды обитания 18° С) при температуре 78°, а у травяной лягушки (средняя температура среды обитания 13°) при 71°? Причем эта корреляция наблюдается, по-видимому, для большинства белков клетки [7].
Было предложено несколько объяснений этой удивительной корреляции (см. [7]). Мне представляется наиболее интересной интерпретация этого явления Н. Г. Есиповой [100], следующим образом объясняющей физические причины (факторы), приводящие в ходе эволюции к соответствию, теплоустойчивость белков с температурой среды обитания пойкилотермных животных *: «Теплоустойчивость — температура денатурации — белков определяется ях аминокислотным составом.
Белки—полимеры и, следовательно, в структурах их макромолекул возможны дефекты двух сортов: 1) «динамические» дефекты, как в обычных атомных 'И молекулярных кристаллах и 2) «статические» дефекты, специфические для полимеров.
Динамические дефекты — нарушение кристаллической структуры вследствие теплового движения атомов в узлах решетки. Это типичные тепловые флуктуации. Динамические дефекты равно быстро возникают и исчезают независимо друг от друга в разных частях кристалла. Концентрация динамических дефектов растет с ростом температуры.
Статические дефекты (понятие, вводимое Н. Г. Есиповой) обусловлены связью отдельных узлов решетки друг с другом посредством полимерной цепи. В результате такой связи небольшие тепловые флуктуации могут распространяться по ходу главных валентностей и сохраняться относительно долгое время «неразряженными», образуя статические дефекты. Парадоксальным образом концентрация статических дефектов уменьшается с ростом температуры, возрастает вероятность «разрядки» на динамическом дефекте.
Макромолекула белка, как показано в работах [98, 99], аналогична кристаллу. Поскольку концентрация статических дефектов в кристалле уменьшается с ростом температуры, кристаллическая структура макромолекулы становится все более совершенной при увеличении температуры до температуры денатурации. Отсюда следует, что, поскольку совершенное функционирование связано с совершенной структурой, для функционирования белков желательна возможно более высокая температура. Однако с ростом температуры растет концентрация динамических дефектов. Для каждого белка и любого другого полимера должна существовать оптимальная температура функ-
! Далее с согласия Н. Г. Есиповой, приведен краткий вариант ее доклада на нашем семинаре.
ционирования в биологических системах, соответствующая минимальной концентрации, статических и динамических дефектов.
Кривая, характеризующая зависимость концентрации дефектов от температуры, определяется типом его структуры, поэтому для разных белков минимальная концентрация дефектов соответствует разным температурам денатурации.
Особый случай представляют собой коллагены, определяющие структуру клеток и морфологию многоклеточных организмов. Минимум дефектов у коллагенов (вследствие особенностей их структуры и особой роли воды в ее поддержании) практически совпадает с температурой денатурации. Поэтому структура коллагенов максимально совершенна на границе тепловой денатурации. Этим объясняется удивительный факт — точная подгонка температуры денатурации коллагенов к температуре среды обитания, достигаемая изменениями аминокислотного состава [100].
О ВОЗМОЖНОСТИ СУЩЕСТВОВАНИЯ ДИСКРЕТНЫХ ТЕМПЕРАТУРНЫХ ЗОН ЖИЗНИ
В интерпретации Н, Г. Есиповой .необходимость соответствия теплоустойчивости белков температуре среды обитания объясняется существованием экстремума — минимума дефектов структуры макромолекулы белка. Однако известно, что макромолекулы белков, вероятно, существуют в нескольких конформациях и их функционирование состоит в переходах из одной конформации в другую.
Представления о возможности существования белков в виде нескольких дискретных конформационных форм — конформе- ров — уже давно обсуждаются в литературе. Мне эти представления стали казаться правдоподобными после опытов сраствора- ми актомиозина [341]. Весьма интересные сведения о температурных переходах конформеров были получены С. В. Коневым и его сотрудниками [148]. Из .их данных следует, что большинство белков резко кооперативно переходит из одного состояния в другое в узком интервале относительно невысоких, заведомо неденатурационных температур (12—13°, 23—25°). Эти состояния различаются, в частности, ферментативной активностью и характером упаковки полипептидных цепей в макромолекуле.
В наших исследованиях [338] выяснилось, что белки существуют в нескольких различных конформациях, -переходящих одна в другую. Отсюда, возможно, следует наличие нескольких температурных зон —разных для разных конформаций, соответствующих -совершенному функционированию. Кроме того, эти температурные зоны могут определяться не самим по себе аминокислотным составом белков, а также и взаимодействием с водою. В этом случае температурная «зона жизии» будет сильно зависеть от свойств воды. В самом деле, мы уже отмечали, что почти любое кон формационное изменение макромолекул белков сопровождается изменением соотношения гидрофобно-гидрофильных групп на поверхности макромолекулы. Следовательно, любое такое изменение должно быть связано с перестройкой, изменением ориентации молекул воды вблизи макромолекул. Это значит, что конформационные изменения последних должны весьма сильно зависеть от легкости перестройки «структуры» воды. В диапазоне условий (содержание электролитов и неэлектролитов, температура, <рН), соответствующим изменениям состояния воды, конформационные изменения макромолекул будут происходить легче, чем вне этого диапазона. Можно довольно легко представить себе «подгонку» свойств полипептидных цепей к свойствам воды посредством изменений аминокислотного состава. В то же время вовсе не очевидно, что и свойства воды могут быть «подогнаны» под любые температурные условия посредством изменения концентраций в ней различных веществ. Вполне может быть, что организмы должны приспосабливаться к неизменяемым свойствам воды.
Тогда может оказаться, что высокосовершенная жизнь может осуществляться лишь при некоторой, определяемой свойствами воды, температуре. Более того, может оказаться, что имеется лишь несколько дискретных температурных зон, совместимых с интенсивной жизнедеятельностью. Существует ли ряд дискретных значений температур, благоприятных для изменений состояния воды? Я говорю здесь о большом числе таких температур. Две из них общеизвестны — 0 и +100°. Предположение о существовании каких-то температур, соответствующих изменению структуры, состояния воды в интервале 0—ТОО0, содержит в себе допущение полиформизма жидкой воды. Само словосочетание «полиморфизм жидкости» парадоксально, так как оно означает «квазикристалличность» жидкой воды. По этому поводу было столько бурных дискуссий, что я не рискую далее затрагивать подобные вопросы. Давно уже дело не в умозрительных общих соображениях, а в надежных фактах (см. [111]).
На протяжении многих лет все данные, способные помочь решению вопроса о полиморфизме жидкой воды, о множественности фазовых переходов в температурном интервале 0—100°, собирал В. Дрост-Хансен [381, 382]. После интенсивного обсуждения этого вопроса [439] был сделан вывод, что в чистой жидкой воде нет полиморфных переходов, множества переходящих при нагревании одно в другое дискретных состояний (структур) воды. Но почти одновременно с таким обескураживающим умозаключением появились данные о полиморфизме воды, находящейся в непосредственной близости от гидрофобных поверхностей, в частности, близ мембран и макромолекул. Я имею в виду прежде всего работу Дрейера и его сотрудников [380], в которой исследовалась температурная зависимость коэффициента диффузии тиомочевины, взятой в низкой концентрации и известной своим слабым влиянием на свойства воды. Авторы цитируемой работы обнаружили ступенчатое возрастание коэффициента диффузии, причем ступеньки начинались при 15, 28, 43 и 60°. Они показали также, что при тех же температурах аномально изменяется и активность ферментов. Вот что .они пишут: «Структурные аномалии воды проявляются в тепловом движении молекул воды ,или в подвижности низкомолекулярных веществ вблизи твердых поверхностей... поверхность макромолекул для низкомолекулярных веществ подобна макроскопической поверхности. Следовательно, .подвижность субстратов в окружении белков должна изменяться в результате структурных аномалий воды». Исходя из этого заключения, авторы цитируемой работы, исследовали температурную зависимость скорости восстановления пирувата, катализируемого лактатдегидрогеназой. Они обнаружили скачкообразное увеличение скорости восстановления вблизи 15° (начало ступеньки) при насыщающей концентрации пирувата и резко аномальное ее изменение в той же области температур при малой концентрации пирувата (порядка константы Михаэлиса Км)-
Последнее обстоятельство представляется особенно интересным. При малой концентрации субстрата общая скорость ферментативной реакции лимитируется диффузией. По млению авторов, структура воды вблизи макромолекул фермента при температуре 14° .препятствует .диффузии субстрата—вода застудневает, а затем .при повышении температуры до 15° и выше она плавится и диффузия облегчается.
Данные об аномальном ходе изменений свойств воды с ростом температуры вблизи твердых поверхностей были получены также в работе [444], в которой измерялась вязкость воды в слоях толщиной 300, 500, 700 и 900 А между двумя кварцевыми пластинками при разных температурах и получались поли- экстремальные зависимости. Обширный материал, свидетельствующий о полиэкстремальном характере зависимости различных биохимических и физико-химических показателей от разных факторов, был получен в работах нашей лаборатории [341, 342]. Наиболее убедительным мне представляется объяснение этих фактов соответствующими изменениями свойств воды. Итак, есть основания для предположения о наличии множества дискретных значений температур, благоприятных для жизни и определяемых свойствами воды.
В соответствии с принятым в книге принципом осталось немного— узнать, есть ли в природе такая дискретность температурных зон жизни. Факты, свидетельствующие о множественности температурных оптимумов для жизни бактерий, в литературе имеются [375, 425, 442]. Однако этих фактов еще недостаточно для общих выводов.
Меня давно занимает мысль о грандиозной статистической обработке данных о температурах, оптимальных для обитания всех пойкилотермных живых существ на Земле. Ясно, что такая статистика даст правильное представление о предмете лишь при прочих равных условиях, а это практически нереально. Можно было бы плыть по морям и океанам на воображаемом корабле от полюса до полюса, измерять температуру и изучать морских обитателей. Более того, после 100 лет морских биологических экспедиций такое путешествие можно совершать, не выходя из библиотеки. Но жизнь сложна — в течение такого путешествия изменяется не только температура морской воды, но и ее минеральный состав, насыщенность газами и освещенность [277а]. Таким образом, получить такие данные можно будет лишь в результате широких исследований 'Искусственно культивируемых живых организмов.
Глава 12
МОЗГ. СПОСОБНОСТЬ к АБСТРАКЦИИ. СОЗНАНИЕ
Невозможность целенаправленных взаимосвязанных перемещений двух произвольно движущихся организмов (жертва — хищник) без экстраполяции взаимных траекторий.
Связь решения в ходе эволюции этой задачи с образованием в мозге аппаратов запоминания, ассоциаций, сопоставления образов, контроля новизны ситуации, принятия решений.
Предел чисто биологической эволюции.
Способность накапливать и использовать сначала случайный, а затем систематический опыт как решающий критерий естественного отбора.
Как следует из сказанного в предыдущей главе, центральная нервная система формируется и совершенствуется при совершенствовании в ходе эволюции аппаратов управления перемещениями в пространстве. Глубокая связь деятельности центральной нервной системы с управлением перемещениями в пространстве была выявлена в трудах И. М. Сеченова [263], Н. А. Бершнтей- на [26], Л. В. Крушинского [159— 161]. Так, Сеченов в своей знаменитой книге «Рефлексы головного мозга» писал: «Все бесконечное разнообразие внешних проявлений мозговой деятельности сводится окончательно к одному лишь явлению — мышечному движению» [263, с. 71].
Замечательны взгляды на эту проблему Б. Спинозы. Старинный язык, геометрический стиль 'изложгния затрудняет в наше время восприятие его идей. Поэтому я приведу изложение взглядов Спинозы, осуществленное с большим искусством Э. В. Ильенковым: «Мыслящее тело активно строит (конструирует) форму (траекторию) своего движения в пространстве сообразно с формой (с конфигурацией >и положением) другого тела, согласовывая форму своего движения (своего действия) с формой этого другого тела, причем любого... Сгособность активно строить свое собственное действие по форм; любого другого тела, активно согласовывать форму своего движения в пространстве с формой и расположением всех другие тел Спиноза и считает отличительным признаком мыслящего -ела, специфической чертой того действия, которое называется «мышлением», «разумом» [119, с. 33 — 34].
Задачу экстраполяции траекторий при управлении перемещениями организма нельзя решить при помощи непосредственной связи рецептора (глаза, например) i эффектора (мышцы).
Более того, наличие подобной связи несовместимо с решением такой задачи. В самом деле, экстраполяция очень часто не совпадает с очевидностью. Я уже приводил пример зенитчика, который должен стрелять не в самолет, а в точку воображаемой траектории его полета, опережающей его видимое положение на много «корпусов». Точно также сокол, пикируя на голубя или утку, должен делать соответствующий «вынос».
Ясно, что в этой сложной задаче орган зрения не должен передавать свои сигналы непосредственно мышцам. Здесь нужно поступать вопреки очевидности. Сигнал зрительного рецептора должен перерабатываться и направляться, вероятно, совсем не «очевидной» группе эффекторов. Должен существовать центральный анализатор сигналов, поступающих от рецепторов. Его функция подобна функции диспетчера. Для принятия решения диспетчер (центральный анализатор) должен заранее знать функции различных эффекторов и успеть (до принятия решения) получить информацию об их состоянии — занятости, работоспособности и т. п.
Следовательно, в сложной многоэтапной системе управления скорость принятия решений зависит от скорости поступления информации (сигналов), сообщаемой всеми потенциально существенными элементами системы управления — другим,и центральными анализаторами (акустическим, тактильным, сигнализирующем об уровне глюкозы в крови), а также возможными эффекторами и пр. Скорость эта будет, естественно, определяться расстоянием, проходимым многочисленными сигналами, необходимыми для принятия решений.
Отсюда ясно, что в ходе естественного отбора должно осуществляться объединение в один орган всех центральных анализаторов и центров управления эффекторами, и таким органом является мозг. Объединение нервных центров в компактные образования — ганглии, а затем, ганглиев в мозг было прослежено в эволюции животных в прошлом веке и получило название цефализации или в греческом произношении кефализации (см. [49, 91, 229]).
Итак, первая стадия принятия решений в задаче управления движением — опрос всех систем, учет состояния всех рецепторов (экстра- и интрарецепторов). Для этого необходима общая команда, приводящая все центральные анализаторы в активное состояние (общий тонус). Такая генерализация тонуса служит условием быстрого принятия решений. В мозге позвоночных тонус создается ретикулярной формацией [60, 219]. Общая мобилизация мозга в связи с решением данной задачи соответствует понятию доминанты, введенному А. А. Ухтомским [299]. Сколько ни ускорять этот процесс опроса, он неизбежно длится заметное время, растущее по мере усложнения системы. Дальнейшее ускорение решения задачи экстраполяции траектории движениядостигается созданием стандартных программ действия при данном сочетании сигналов от рецепторов.
Такой способ экстраполяции имеет вероятностый характер и основан на следующем предположении: если раньше данное действие (перемещение) при данном сочетании сигналов от рецепторов неоднократно приводило к правильному результату, то велика вероятность того, что такие же действия при данном сочетании сигналов от рецепторов окажутся правильными и впредь. Реализация решений, основанных на этом способе прогнозирования, и есть, по-видимому, то, что И. П. Павлов назвал условным рефлексом (динамическим стереотипом). Такой способ подобен предсказанию погоды по аналогии: прогноз составляют на основании многолетних наблюдений. Если неоднократно ранее при определенном сочетании температуры, давления, скорости и направления ветра и пр. на следующий день бывала ясная солнечная погода, значит, при том же их сочетании и завтра можно с некоторой вероятностью ожидать такой же погоды.
Выработка условных рефлексов резко ускоряет принятие решений. Хорошо тренированный охотник, не размышляя, выносит прицел на семь корпусов вперед от летящей утки и, побеждая в естественном отборе неопытного охотника, жарит утку вечером на привале. Выработка стандартной программы, т. е. условнорефлекторного поведения не только резко ускоряет совершение целесообразных действий, но .и делает ненужной общую мобилизацию мозга при каждом акте принятия решений. Такая мобилизация, вдохновение, остается необходимой лишь при выработке новых программ. Следовательно, должен существовать механизм, определяющий новизну ситуации, устанавливающий, что для данного сочетания сигналов от рецепторов готовых программ действия нет. Ситуация 'новизны должна вызывать общую мобилизацию анализаторов, приводить их в активное, я бы сказал, творческое состояние. Механизмы оценки .новизны и те структуры мозга, которые осуществляют оценки, весьма успешно исследуются О. С. Виноградовой [52], к ее работам и следует обращаться читателю.
Мне необходимо лишь подчеркнуть, что функционирование аппарата различения новых ситуаций от бывших ранее играет решающую роль в кинетике поведения животных. Чем быстрее задача будет сведена к предыдущей, тем быстрее будет принято решение, тем успешнее окажется экстраполяция траекторий движения. В самом деле, застывший в недоумении и нерешительности охотник не успевает выстрелить и справедливо называется раззявой.
Такое же значение имеет скорость принятия новых решений, т. е. скорость создания новых программ действия, новой схемы подключения эффекторов. Вопрос этот очень сложен, однако, мне представляется, что в большинстве случаев новые решения принимаются по аналогии с принимавшимися ранее. Чем большенакопленный опыт, чем богаче память, тем вероятнее нахождение аналогий. Скорость возникновения ассоциаций, установления аналогий зависит от развития ассоциативных связей между различными центральными анализаторами.
Таким образом, совершенство мозг.а определяется совершенством разрешающей способности центральных анализаторов, сигналов от периферических рецепторов, скоростью создания стандартных программ действий, емкостью памяти (склада готовых программ), совершенством аппарата различения новое — старое и ассоциативного аппарата, быстротой передачи сигналов эффекторам.
При всей соблазнительности более детального анализа механизмов, обеспечивающих функцию мозга из общих соображений, я не решусь на это. Как было бы интересно представить себе, например, роль света, передаваемого по нервам-световодам, в возникновении ассоциаций. Тут бы и стало физически реальным поэтическое понятие «озарение». Как заманчиво рассмотрение возможных биохимических механизмов регулирования эмоций, тонуса различных отделов мозга, анализ возможных механизмов памяти [4, 11, 52, 60, 224, 324, 469]. Но действительные механизмы функционирования мозга еще далеко не выяснены и поэтому у нас не будет должного критерия, контроля верности дедуктивных построений.
Предельное совершенство мозга предельно совершенных хищников и жертв — предел чисто биологической эволюции. Люди — это организмы, способные перерабатывать вещества и использовать энергию не только при помощи биологических процессов, но и посредством небиохимических и нефизиологических механизмов. Эволюция человека основана на отборе по признаку все более совершенного использования сначала случайного, а затем систематического опыта.
Рассмотренные нами абстрактные предельно совершенные жертвы и хищники — предел чисто биологической эволюции. Они предельно совершенны биохимически, т. е. скорость биохимических процессов у них предельно велика в той степени, в какой это допускают физические и химические свойства возникших в эволюции систем. Они обладают предельным совершенством в основных физиологических процессах. Они движутся по произвольным траекториям с предельной, лимитируемой все теми же физическими и химическими законами скоростью и предельно совершенно управляют своим перемещением в пространстве.
Такие организмы предельно совершенны еще в одном, чрезвычайно существенном отношении, установленном в 1922 г. А. Н. Северцовым [254]. Возникшая под давлением естественного отбора высоко совершенная нервная система обусловливает новый способ приспособления к условиям существования. Этот способ состоит в соответствующем изменении поведения.
Северцов отмечает, что известно три основных типа психической деятельности — поведения, а именно: рефлекторное, инстинктивное и деятельность «разумного» типа. У членистоногих преимущественное значение в ходе эволюции приобрело поведение, основанное на инстинктах (см., однако, [188]). \ хордовых преимущественное развитие приобрела психика «разумного» типа.
При инстинктивной и рефлекторной деятельности наследуются сами эти действия, т. е. последовательность определенных движений. Они эволюционируют, как и прочие признаки, т. е. (что подчеркивает Северцов) -крайне медленно, посредством отбора благоприятных мутаций. Так обеспечивается приспособление к медленным изменениям внешней среды. При деятельности «разумного» типа наследуются не сами действия, а лишь определенный уровень психической организации, структурной и функциональной организации нервной системы.
Северцов отмечает, что при быстрых изменениях условий существования в неблагоприятную сторону у животных с высоко организованной психикой «выживут и приспособятся те, которые окажутся способными быстро и целесообразно изменять свое поведение и выработать новые привычки, особи с потенциально более высокой .психикой, т. е. животные наиболее умные и наиболее способные; говоря метафорически, выживут «изобретатели новых способов поведения». Другими словами, при эволюции этим путем повышается потенциальная психика, причем дело здесь идет уже о наследственном повышении .психических способностей» [254, с. 43]. Эволюция .при этом резко ускоряется. В относительно недавнее время в этом намеченном Северцовым направлении появились новые исследования [124, 159, 160, 181а].
Мы, естественно, не могли обсудить здесь все направления биохимического и физиологического прогресса. Моя задача состояла в выявлении ведущего физико-химического или чисто биологического критерия, фактора биологической эволюции. Однако и при таком схематическом рассмотрении можно придти к выводу, что после возникновения предельно совершенных жертв и хищников все в биосфере, все запасы вещества и энергии, все что может превращаться в вещества, входящие в состав этих предельно совершенных организмов, будет использоваться с предельно возможной скоростью. Я не говорю о ее величине — я только подчеркиваю, что по достижении предельной скорости наступает физико-химический предел чисто биологической эволюции
Однако предел чисто биологической эволюции не означает вообще предела увеличения скорости превращения неживой материи в живую. Новый этап резкого ускорения превращения веществ в «живую материю» начинается с возникновения человека. Человек— прямой наследник организмов, обладающих в принципе предельна говершенной центральной нервной систе
мой. Способность к анализу произвольных траекторий — к восприятию, к ассоциациям, к сопоставлению образов, а также большая емкость памяти, наличие высокосовершенных анализаторов— обусловливают возможность выхода на новый этап прогресса.
Этап этот основан на использовании (наряду с чисто биохимическими и физиологическими) небиологических механизмов переработки веществ и преобразования энергии. Для лаконичности можно произнести одно слово «огонь». В этом смысле Прометей и был первым человеком.
Огонь впервые был использован случайно, в случайном опыте, но теперь в естественном отборе будут побеждать организмы, способные все более совершенно использовать случайный опыт, сделать его систематическим, проводить, по существу, активное исследование окружающей природы.
Последнее обстоятельство весьма важно. До применения огня материал для естественного отбора поставляли только случайные изменения, мутации наследственных молекул. Теперь критерием совершенства мутации, их полезности стала способность «мутантов использовать случайный опыт». С некоторого уровня биологического совершенства роль мутаций оказывается совсем незначительной и, быть может, прогресс в целом определяется не столько накоплением полезных мутаций, сколько накоплением полезного опыта [27, 93].
Организмы предельно совершенных жертв или хищников в принципе приспособлены для использования случайного опыта. На этом, как мы видели, основана их повседневная жизнь, требующая решения задач экстраполяции траекторий, способность к обучению и т. п. Их мозг .в принципе годится « Прометею. Однако степень совершенства мозга еще далеко не достаточна. Использование случайного и, тем более систематического, опыта для интенсификации жизнедеятельности требует мощного развития мозга и особенно его ассоциативных центров.
Так оно 'И происходит в эволюционном становлении человека.
Возрастание роли жизненного опыта, умения, знаний, а также науки, искусства в биологической судьбе популяции или вида приводит к еще одному замечательному обстоятельству. Отобранные в естественном отборе прогрессивные мутации передаются при размножении организмов, но накопленный жизненный опыт таким путем не передается — он сохраняется вследствие обмена опытом в популяции, вследствие обучения. Носителями этого жизненно необходимого опыта оказываются не отдельные особи, а популяции — коллективы. Поэтому люди образуют не чисто биологические популяции, а социальные коллективы, и предметом естественного отбора становится и совершенство обмена опытом, знаниями, совершенство обучения. И раньше, до появления человека такой обмен имел биологический смысл, но теперь он приобрел решающее значение в интенсификации превращения веществ (интенсификация является автоматическим следствием давления отбора).
В результате социально-биологической эволюции человека вырабатываются небиохимические методы превращения веществ— от варки пищи до химической промышленности; небиохимические, небиофизические способы преобразования энергии— от сжигания веществ (огонь) до использования электричества и атомной энергии; нефизиологические механизмы перемещения в пространстве — от телеги до спутника; нефизиологические способы хранения, передачи и увеличения информации— письменность, библиотеки, радио, телеграф, телевидение, вычислительные машины.
Иными словами, в принципе все чисто биохимические, биофизические, физиологические механизмы перестают быть определяющими факторами увеличения интенсивности преобразования неживой материи в живую.
В сущности и в социально-биологической эволюции человека ведущим фактором оказывается кинетическое совершенствование, измеряемое интенсивностью преобразования веществ и энергии окружающей среды. Это прослеживается в смене каменного века железным, века пара веком электричества и т. д. Но подобные вопросы далеко выходят за пределы нашей задачи и мы не будем их рассматривать дальше [174].
На этом можно было бы завершить наш очерк эволюции. Стоит, однако, еще раз подчеркнуть, что всякий новый ароморфоз приводит к обновлению биосферы и заполнению всех возможных ареалов обладателями новых принципиальных достоинств. Все организмы, принадлежащие к предыдущим эволюционным этапам, остаются существовать в биосфере лишь в количествах, совместимых с существованием более совершенных организмов. Наличие множества сложных реальных ситуаций (паразитизм, безразличное сосуществование) не изменяет принципиальной верности этого положения.
Глава 13
ОБЩИЙ ИТОГ. ДЕТЕРМИНИРОВАННОСТЬ ТРАЕКТОРИЙ БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ — СЛЕДСТВИЕ ЕСТЕСТВЕННОГО ОТБОРА
Общий очерк эволюционного процесса показывает, что на всем его протяжении действуют критерии естественного отбора, основанные на величине <гкинетического совершенства», т. е. все большейитоговой скорости увеличения массы вещества данного вида. Кинетическое совершенство на разных этапах эволюции определяется разными факторами эволюции.
Эволюция на каждом этапе совершается до исчерпания эволюционного потенциала очередного фактора эволюции.
Основные этапы биологической эволюции детерминированы физическими и химическими свойствами эволюционирующей системы.
В предыдущих главах мы совершили восхождение от первичных макромолекул, способных к конвариантному воспроизведению, до человека. При этом мы пытались выявить физико-химические критерии естественного отбора на разных этапах биологической эволюции. Анализ физико-химических факторов биологической эволюции отнюдь не был исчерпывающим. Не были рассмотрены такие важные для эволюции ряда таксономических групп критерии, как механическое совершенство организмов — совершенство рычагов и способов приложения сил в аппаратах перемещения в пространстве, конструкции скелета, стеблей, стволов. Не обсуждались с должной тщательностью гидро- и аэродинамические характеристики приспособлений для активного и пассивного полета или плавания. Этот перечень можно продолжить. Однако нашей задачей и не являлось рассмотрение всех таких факторов. Важно было лишь показать принципиальную возможность анализа физико-химических факторов в качестве критериев, определяющих направления биологического эволюционного процесса.
Направление эволюции определяется, как отмечено во 2-й главе, и собственно биологическими факторами. Эти факторы, критерии естественного отбора, сами возникают в процессе эволюции. Совершенствование по критериям собственно биологических факторов обусловливается главным образом выбором траектории, движение по которой в направлении роста биологического (кинетического) совершенства идет с наибольшей скоростью. Скорость возрастания биологического совершенства является .мерой биологического прогресса (см. гл. 2, с. 29).
Выступающие в определенной последовательности физико-химические и собственно биологические факторы делают процесс биологической эволюции строго детерминированным. Речь идет, конечно, о детерминированности основных этапов. Реальные траектории в сильной степени зависят от множества «внешних» обстоятельств — геологических и космических в первую очередь.
Тем не менее основные этапы эволюции представляются детерминированными настолько, что они могут быть получены посредством общих соображений. При этом для выявления основных этапов нет необходимости прибегать к помощи ЭВМ. Достаточны приближенные оценки, основанные на принципе предельного совершенства.
Однако для оценки скорости эволюции, времени, необходимого для достижения определенной величины биологического совершенства, нельзя обойтись без построения теории скоростей биологической эволюции, построения математических моделей и их просчета посредством ЭВМ (см. (12, 50, 122, 128, 139, 203— 205, 243а, 305, 308, 319, 321, 372]).
При анализе направлений эволюционного совершенствования соответственно разным физико-химическим факторам выявляется общая схема эволюционного процесса—из множества возможностей в процессе естественного отбора выбирается в пределе одна. Общий образ эволюции состоит, следовательно, в выборе уникальной траектории в многомерном пространстве . Так происходит при становлении системы биохимических реакций и при совершенствовании каждой биохимической реакции в отдельности. Так же идет выработка механизмов управления перемещениями организмов. И в самом общем виде — так происходит при отборе уникального полинуклеотидного (и полипептидного) текста из безграничного множества возможных сочетаний нуклеотидов или аминокислот.
Следовательно, естественный отбор — основа дарвинизма — причина детерминированности биологической эволюции
Существует мнение, что такой детерминизм нереален, поскольку каждый эволюционный шаг в пространстве параметров возможных направлений исключает другие шаги, а этот каждый шаг случаен (Л. А. Блюменфельд [29]).
Весь вопрос в степени необратимости эволюции, т. е. в конце концов в необходимом времени для достижения определенной степени биологического совершенства. В истории Земли нам известны примеры чрезвычайно быстрой эволюции, равно как и примеры застывания в неизменном виде на протяжении многих сотен миллионов лет. В лабораторной практике удается получить новые наследственные формы микроорганизмов с очень большой
скоростью. Достаточно быстро получаются и «обратные мутанты». Так что мнение о полной необратимости эволюции может быть и неверно.
С другой стороны, в случае различных далеко зашедших в определенных направлениях эволюционных траекторий под действием одинаковых факторов возникают лишь функционально сходные организмы. Это явление конвергентного сходства иллюстрирует высокую степень детерминированности эволюционного процесса. Примеры биологической конвергенции приведены в Приложении.
Итак, в ограниченном пространстве (ареале), при наличии источника свободной энергии, конвариантно воспроизводящиеся (матричные) молекулы полиморфных полимеров с неизбежностью вступают в конкуренцию друг с другом за вещества и энергию, необходимые для построения их копий. Происходит естественный отбор мутантов по признаку кинетического совершенства— все большей итоговой скорости превращения веществ окружающей среды в вещества данного вида. Конвариантно воспроизводящиеся матричные полимерные молекулы — предмет естественного отбора — становятся объектом эволюционного развития. Такая эволюция называется биологической эволюцией. Ее объекты — живые существа. Процесс их существования — жизнь.
Направление биологической эволюции определяется физикохимическими и биологическими факторами. Эволюционное развитие направлено в сторону все большего кинетического (биологического) совершенства. Как следствие этого, за счет сопряженной траты термодинамического потенциала системы, объекты эволюции развиваются в направлении все большего усложнения, все большего удаления от термодинамически вероятной случайной последовательности мономеров в полипептидной цепи.
Кинетическое совершенство на первых этапах биологической эволюции достигается посредством улучшения в ходе Естественного отбора механизмов матричного воспроизведения и формирования ферментов — макромолекулярных полимерных катализаторов.
Система предельно совершенных ферментов обеспечивает протекание все более сложных биохимических процессов, приводящих ко все более интенсивному преобразованию веществ среды в вещества данного вида.
Чисто биохимическая эволюция ограничивается скоростью диффузии метаболитов. В качестве приспособлений, уменьшающих диффузиционные ограничения и расход вещества, возникают «органеллы» — митохондрии и хлоропласта.
Кинетическое совершенствование обусловливает дискретность «живого вещества» — образование дискретных комплексов оптимальных размеров, содержащих необходимый и достаточный набор матричных молекул, ферментов, метаболитов. Существование этих комплексов — организмов становится возможным при фор
мировании поверхностных мембран, препятствующих растеканию комплексов.
Необходимость поддержания целостности поверхностных мембран, быстрого реагирования на их повреждение обусловливает развитие сигнальной системы, срабатывающей посредством нарушения ионного градиента. Ионный градиент — неравновесное распределение одновалентных катионов — в системе организмов (клетка — внешняя среда) активно поддерживается биохимическими процессами. Нарушение этого градиента, изменение внутриклеточной .концентрации одновалентных катионов активирует процессы синтеза биохимических 'компонентов мембраны.
Диффузионные ограничения кинетического совершествования преодолеваются двумя путями — посредством по преимуществу морфологического прогресса и посредством развития аппаратов активного перемещения в пространстве. Первый путь в основном соответствует эволюции автотрофов, живущих в векторизованных потоках вещества и энергии (растения).
Второй путь — эволюция гетеротрофов. Гетеротрофы развиваются в силу способности заполнять ареалы, не ограниченные непрерывным поступлением векторизованных потоков вещества и энергии. Эволюция гетеротрофов с активным перемещением в пространстве определяется совершенствованием аппаратов, обеспечивающих такое перемещение. В связи с необходимостью (под давлением Естественного отбора) увеличения абсолютной скорости перемещений в пространстве у.ндулоподии ( жгутики, реснички) заменяются, в качестве органов движения, мышечными волокнами. Мышечные волокна обеспечивают движение посредством из;менений формы клеток. Совершенствование перемещений в пространстве состоит далее в создании аппаратов управления направленным движением в среде. Такое управление основывается на развитии системы рецепторов, сигналы от которых поступают к органам движения. Первоначально рецепторами были сами органы движения (ундулоподии). Связь специализированных рецепторов со специализированными органами движения осуществляется посредством нервных волокон.
Переход от ундулоподий к мышечным волокнам, при развитии аппаратов управления движением, обусловил возникновение многоклеточных организмов, состоящих из специализированных дифференцированных клеток.
Совершенствование управления перемещениями, движениями в среде многоклеточных организмов приводит к совершенствованию нервной системы, к возникновению мозга.
Мозг достигает .предельного качественного совершенства при формировании аппаратов управления движением «предельно — совершенных жертв и хищников». Существование этих организмов основано на способности определения необходимой произвольной траектории движения в пространстве в связи с независимой произвольно?' траекторией другого организма, равного поабсолютной скорости движения. Способность построения таких траекторий основана на способности восприятия, анализа, памяти, сопоставления образов, принятия решений и их реализации. «Предельно —совершенные жергвохищники» являются пределом чисто биологической эволюции.
Дальнейшая интенсификация процессов превращения веществ среды в вещества данного биологического вида возможна лишь при использовании также и небиохимических, небиофизических и нефизиологических механизмов. Такое использование «огня» может быть основано сначала на случайном, а затем систематическом опыте. Эволюционное развитие в этом направлении определяется количественным совершенствованием мозга. Организмы, эволюционно развивающиеся в этом направлении, — люди. Необходимость накопления и сохранения «жизненного опыта» в поколениях, определяющая победу в Естественном отборе, обусловливает социальный характер этих организмов.
ПРИЛОЖЕНИЕ
ПРИМЕРЫ БИОЛОГИЧЕСКОЕ КОНВЕРГЕНЦИИ, ИЛЛЮСТРИРУЮЩИЕ ДЕТЕРМИНИРОВАННОСТЬ НАПРАВЛЕНИЙ БИОЛОГИЧЕСКОЕ ЭВОЛЮЦИИ УСЛОВИЯМИ ЗАДАЧ, РЕШАЕМЫХ ПОД ДАВЛЕНИЕМ ЕСТЕСТВЕННОГО ОТБОРА [21]
Биологическая конвергенция — образование сходных признаков на разных траекториях эволюционного процесса — важное свидетельство детерминированности биологической эволюции. Анализ имеющихся данных показывает, что конвергенция является скорее правилом, чем исключением, и захватывает не только морфологию, но и физиологию и биохимию неродственных организмов. Неоднократное возникновение в принципе одинаковых приспособлений на разных эволюционных направлениях означает существование предельно совершенного решения. К этому решению стремятся эволюционные траектории, берущие начало в разных местах филогенетического древа.
Ниже приводятся примеры биологической конвергенции разного рода, свидетельствующие о могуществах естественного отбора.
Строение органов чувств животных особенно явно определяется физическими факторами. Поэтому эти органы часто обладают конвергентным сходством. Неоднократно на разных эволюционных направлениях возникали глаза. Наиболее примитивные глаза способны лишь отличать свет от темноты. Однако независимо несколько раз в ходе эволюции возникают настоящие гла
за, в которых создается более или менее детальное изображение окружающего мира. На рис. 25 показано строение глаза гидромедузы (кишечиополостиое), кольчатого червя (червь), каракатицы (головоногий моллюск) и человека (млекопитающее). Мы видим везде прозрачную наружную стенку глаза, стекловидное тело, заполняющее пространство глазной камеры, линзу — хрусталик, расположенные в один слой светочувствительные клетки, отходящий от глаза зрительный нерв. Все глаза (кроме глаза медузы) способны к аккомодации; у кольчатого червя и головоногих моллюсков (а также у рыб) оиа осуществляется посредством перемещения хрусталика, у человека — путем изменения формы хрусталика. Наблюдаемое различие вызвано оптическими свойствами среды обитания.
Гидродинамические законы обусловили конвергентное сходство формы тела у акул, ихтиозавров и дельфинов (рис. 26).
Мы уже рассматривали проблему движения воды при обсуждении происхождения цветковых растений. Скорость проведения воды от подземной чаек» растения к фотосинтезирующим органам неоднократно становилась «узким местом» в процессе увеличения биологического совершенства разных групп растений. Наиболее эффективное преодоление этой трудности — создание труб,

РИС. 25. Конвергентное сходство строения глаз различных животных [14] а — эпителиальный глаз гидромедузы;
—глаз кольчатого червя; в —глаз головоногого моллюска; г — глаз человека; 1— нервные отростки зрительных клеток (в а) или зрительный нерв; 2—зрительные клетки н светочувствительные палочки на их концах (в а) или палочки у остальных; 3— стекловидное тело; 4 — хрусталик; 5 — радужная оболочка; 6 — передняя камера;
— роговица
РИС. 26. Конвергентное сходство формы тела акулы (рыба) (а), ихтиозаз- ра (рептилия) (б), дельфинов (млекопитающие) (в, г) [333]
по которым вода движется, не встречая препятствий (клеточных стенок). Такое решение было найдено независимо пятью группами, виды, имеющие водопроводные трубы, встречаются почти у всех высших растений, а именно у плау- новидиых, членистых, папоротниковидных, голосеменных и покрытосеменных. Доказано многократное возникновение водопроводных труб у разных видов покрытосеменных. Способы образования сосудов в разных группах высших растений неодинаковы. У одних проводящая воду полость образуется в результате разрушения первичных проводящих клеток (хвощи), у других, проводящие клетки становятся члениками сосудов — трахей, отверстия в их стенках образуются за счет редукции замыкающих пластинок пор. Наконец, у третьих проводящие клетки превращаются в сегменты сосудов (как и в предыдущем случае), но отверстия образуются в результате разрыва клеточной оболочки (рис. 27).
Внешнее строение большинства активных животных характеризуется двусторонней симметрией. Асимметричное животное ие способно к эффективным направленным перемещениям в пространстве. В большинстве случаев двусторонняя симметрия разных видов может рассматриваться как признак, унаследованный ими от общего предка. Известно, одиако, несколько случаев, когда внешняя двусторонняя симметрия возникала у потомков асимметричных животных в результате их возвращения к активному образу жизни.
В процессе эволюции брюхоногие моллюски выработали спирально закрученную раковину и стали асимметричными. Мы не будем останавливаться н» причинах этого явления. Здесь важно другое. Несколько семейств из классу брюхоногих, объединяемые названием голые слизни, и две группы морских брюхоногих (голожаберные и часть крылоногих) утратили раковину и стали более активными, причем их строение вновь приобрело двустороннюю симметрию (рис. 28). Одиако расположение внутренних органов вернулось к исходной симметрии лишь у некоторых голожаберных, да и то не полностью. Что касается всех остальных, то они полностью сохранили внутреннюю аснммет-, рию своих предков, или даже (крылоногие) усложнили ее.
Сходный процесс произошел и среди ракообразных. Известны раки-отшельники, живущие в раковинах брюхоногих моллюсков и ставшие из-за этого асимметричными. Один из потомков раков-отшельников, вылезший из раковины и перешедший к более активному образу жизии, изображен на рис. 29. Части тела этого рака, существенные в активном движении, двустороине- симметричиы.
Конвергенция происходит под действием ие только биофизических, но и чисто биологических факторов.
Для многих животных характерны различные защитные приспособления. На рис. 30 изображены представители трех различных типов животных: мидия — моллюски, класс двустворчатых моллюсков, остракода тип членистоногих, класс ракообразных, подкласс ракушковые рачки и представитель типа плеченогих. Все оии выработали сходные по строению двустворчатые раковины. У двустворчатых моллюсков и остракод створки раковины находятся по бокам тела, а у плеченогих сверху и снизу. Интересно, что и у плеченогих, и у двустворчатых моллюсков имеются формы, имеющие специальное приспособление (замок), служащее для скрепления створок раковины, но имеются и такие,.



РИС. 27. Конвергентное сходство разных форм члеников водопроводных сосудов покрытосеменных растений (а) и члеников сосудов плауиовидного растения селагинел- лы (б) [282]
РИС. 28. Двусторонне-симметрич ные моллюски — потомки асимметричных предков [ЮЗ] а — голожаберный моллюск Dendrono- ius frontosus; б — крылоногий моллюск Clione lltna- cina (морской ангел); в — слизень Troglolestes sp.

РИС. 29. Двусторонне-симметричиый рак Lytodes maja — потомок асимметричных раков-отшельников [10]
Асимметрия сохранилась в одностороннем расположении брюшных ножек
у которых замка нет. Замечателен случай образования двустворчатой раковины у брюхоногого моллюска, изображенного на рис. 31.
Эволюция двух групп живых существ-покрытосеменных растений и членистоногих животных буквально пронизана конвергенцией, причем, именно эти группы характеризуются наибольшим числом, разнообразием видов.
Перед растениями, вышедшими на сушу, встает задача обеспечения встречи мужской и женской гамет, произведенных желательно разными особями. Эта задача решалась по-разному. Отметим поэтому лишь конвергенцию, связанную с одним из возможных решений. Хорошо известно, что цветки большинства покрытосеменных опыляются насекомыми. Исходно насекомых привлекала сама пыльца, а затем растения научились выделять особую сладкую жидкость — нектар. Для улавливания переносимой насекомыми пыльцы образовался специальный орган—пестик (точнее его рыльце) или несколько стило- диев. Оказывается аналогичные образования возникли и у некоторых ржавчинных грибов (дальнейшее относится к виду Пукциния граминис). В жизненном цикле этого гриба чередуются две формы. Одна из иих, назовем ее первой, диплоидна (дикарионна) и живет на хлебных злаках. Другая, вторая форма, гаплоидна и живет иа листьях барбариса. Вторая форма развивается из гаплоидных спор, образуемых первой формой в результате редукционного деления. Первая форма в свою очередь развивается из диплоидных спор, возникающих в ходе жизни второй формы. Для формирования диплоидных спор у второй формы необходимо оплодотворение. Оно может происходить при встрече мицелиев разных полов (точнее разных знаков — полы здесь ие различимы морфологически). Такая встреча возможна, если мицелии разных знаков вы-

РИС. 30. Конвергентное возникновение двустворчатой раковины у представителей трех различных таксономических типов животных [103]
а — двустворчатый моллюск Mytilus edulus; б — ракушковын рачок Cyprls pubera; в — плеченогие

РИС. 31. Конвергентное возникновение двустворчатой раковины у брюхоногого моллюска Bertelinia sp. [103].

росли иа одном листе. Но если они выросли на разных листах, оплодотворение все равно возможно. Вторая форма образует иа листе барбариса особые органы — пикииды, аналогичные цветкам покрытосеменных растений (рис. 32). Наружу из пикииды высовываются пучки гиф и выделяется пахучая сахаристая жидкость — иектар, в которую погружены пикноспоры, играющие роль
РИС. 32. Пикниды ржавчинного гриба Puccinia graminis, находящиеся на листе барбариса и имеющие конвергентное сходство с цветком
Из пикниды высовываются пучкн гиф и выделяется пахучая сахаристая жидкость — нектар, привлекающий насекомых
пыльцы. Пикноспоры переносятся привлеченными нектаром насекомыми и улавливаются гифами, аналогичными пестикам.
В эволюции покрытосеменных четко прослеживается несколько направлений. Об одном из них, а именно, об эволюции от бессосудистых к сосудистым, мы уже упоминали. Другой характерный пример — переход от видов, со свободнолепестным венчиком к видам со спайнолепестным венчиком. Раньше этот признак использовали в систематике для выделения двух классов — Свободнолепестных и Спайнолепестных. Теперь же выяснилось, что Спайнолепестные не класс, а сборная группа, так как эволюция многих групп раздельнолепестных кончалась образованием спайнолепестного венчика.
Другой пример всемогущества конвергентной эволюции — превращение на разных направлениях эволюции радиально-симметричных цветков в двусторон- не-симметричные (актиноморфиых в зигоморфные). На рис. 33 изображены зигоморфные цветки представителей разных семейств, от лютиковых до губоцветных.
Характерным примером сложной конвергенции может служить образование травянистых растений из древесных предков. Такой переход деревьев в травы неоднократно происходил независимо в разных группах.
В эволюции членистоногих замечательно конвергентное возникновение наружного покрова ■— кутикулы. Многократно присходила дифференцировка исходной гомогенной кутикулы с образованием нескольких слоев, близких по функциям и строению в разных филах. Так, М. С. Гиляров пикнет:' «Эти слои у разных форм, если они одинаково располагаются, можно сравнивать, считая их соответственно эидо-, экзо- и эпикутикулой, но при этом их следует рас- сматривать как образования аналогичные (конвергентные или параллельные), но не как гомологичные» [60].
Для большинства членистоногих, живущих иа суше, характерно дыхание трахеями — сообщающимися с атмосферой трубками, содержащими воздух н пронизывающими тело животного. Происхождение трахей у всех членисто-

РИС. 33. Конвергентное образование двустороннесимметричных венчиков цветков в разных семействах цветковых растений в результате приспособления к опылению насекомыми [187, 222]
/ — Караганник (Бобовые); 2— Недотрога (Бальзаминовые); 3—Хохлатка (Дымянковые); 4— Буквица (Губоцветные); 5 — Льняика (Норичниковые); £—Ятрышник (Орхидные); 7—Живокость (Лютиковые); 8 — Шандра (Губоцветные); 9 — Синяк (Бурачниковые); 10— Заразиха (Заразиховые)


РИС. 34. Сопоставление строения брюшного мозга стрекозы (а) и спиниого мозга человека (б)
Функционально сходные структуры изображены сходным образом. Сильно упрощенно [108]
ногих в принципе одинаково: трахеи представляют собой видоизмененные впя- чиваиия утонченных участков кутикулы, несших вначале дыхательную функцию. Но возникали они конвергентно более чем на 10 направлениях эволюции. Гиляров пишет по мому поводу: «Трахеи разных групп членистоногих даже в случае глубокого сходства их структуры, гомологичны друг другу только как производные поьровов, выполняющих функцию дыхательных органов. Одиако как специалАЗированные системы органов трахейные системы разных групп наземных членистоногих аналогичны, их сходство определяется сходными условиями и принципами функционирования.» [66].
Можно было бы привести пример с конвергенцией выделительного аппарата членистоногих [независимое образование мальпигиевых сосудов), а так
же ряд других, ио мы ограничимся перечисленными выше, и отошлем читателя к специальной литературе [67].
Конвергентное решение близких эволюционных задач в разных филогенетических направлениях очень характерно также для тканевого уровня организации. Детерминированность конечного результата условиями задачи проявляется здесь с особой яркостью А. А. Заварзин писал: «Нервная система при сравнительном изучении позволила обнаружить основную закономерность, господствующую в гистологии, которая казалась на первых порах парадоксальной, и которая заключается в том, что даже очень сложные и специализированные тканевые системы, несмотря на огромные анатомические и филогенетические отличия тех органов, которые ими образованы, оказываются очень сходными друг с другом даже в деталях» [108]. Заварзин иллюстрирует это положение, сопоставляя строение спинного мозга человека и брюшного мозга стрекозы. Соответствующие поперечные разрезы свидетельствуют о замечательном конвергентном сходстве обоих образований (рис. 34).
Другим важным примером гистологических параллелизмов может служить конвергентная утрата ядер эритроцитами. Безъядерные эритроциты в ходе эволюции появлялись независимо четыре раза: у немертин, кольчатых червей, иглокожих и млекопитающих. Заметим, кстати, что сама кровеносная система у немертин, кольчатых червей и хордовых возникала совершенно независимо.
Приведенные выше примеры имеют главным образом морфологический и физиологический характер. Это ие значит, что биохимическая конвергенция менее распространена. Она просто меиее изучена.
Широко известна способность насекомоядных растений переваривать пойманных насекомых посредством внешнего пищеварения с помощью желудочного сока, аналогичного ферментам животных. Яркой иллюстрацией биохимической конвергенции служит независимое неоднократное возникновение в ходе эволюции одних и тех же пигментов крови [150]. Особенно, на наш взгляд, замечательно наличие голубой крови, переносящей кислород посредством медьсодержащего пигмента гемоцианина, у брюхоногого моллюска-прудовика и красной крови, содержащей гемоглобин, у столь же распространенного брюхоиогого моллюска — катушки.
Наконец, примером биохимической конвергенции может служить наличие; гемоглобина в корневых клубеньках бобовых растений — так называемого леггемоглобииа. Как показал недавно посредством рентгеноструктурного анализа В. В. Борисов [34], молекула этого растительного гемоглобина даже по своей пространственной структуре практически неотличима от гемоглобина эритроцитов животных.
В заключение приведем слова Э. Майра: «Животный мир (так же как и растительный) полон конвергенций, когда сходные требования среды вызывают сходный фенотипический ответ у неродственных или, по крайней мере, не близкородственных организмов... Поговорка «все дороги ведут в Рим» также справедлива в применении к эволюции, как и в житейских делах» [189].
ЛИТЕРАТУРА
Абиогенез и начальная стадия эволюции жизни. М.: Наука, 1968.
Агол И. И. Диалектический метод и эволюционная теория. М.: ГИЗ, 1930.
Агол В. И. О системе вирусов.— Усп. соврем, биол., 1974, т. 77, с. 9.
Азарашвили А. А. Изменение продолжительности хранения приобретенных навыков при «разрушении» диссоциированных состояний.— Журн. высш. нервн. деят., 1974, т. 24, с. 1189.
Александров А. А., Берестовский Г. Н„ Волкова С. П., Востриков И. Я., Жерелова О. М., Кравчик С., Луневский В. 3. Реконструкция одиночного кальций-натрневого канала клетки в липидном бислое.— Докл. АН СССР,
т. 227, с. 723.
Александров В. Я. О связи между теплоустойчивостью протоплазмы и температурными условиями существования.— Докл. АН СССР, 1952, т. 73, с. 149.
Александров В. Я. Клетки, макромолекулы и температура. М.: Наука, 1975.
Аллен А. О. Радиационная химия воды и водных растворов. М.: Госатом-
издат, 1963.
Антонов А. С. Мы похожи, но насколько? М.: Знание, 1975.
Ароннет Н. И. Мышечные и клеточные сократительные (двигательные) модели. М.: Наука, 1971.
Ашмарин И. П. Загадки и откровения биохимии памяти. Л.: ЛГУ, 1975.
11а. Ашмарин И. П. Молекулярная биология. Л.: ЛГУ, 1977.
Базыкин А. Д. Отбор и генетическая дивергенция в системах фкальных популяций и популяциях с непрерывным ареалом (математическая модель).— В кн.: Проблемы эволюции, 1973, т. 3, 231 с.
Бауэр Э. С. Теоретическая биология. М.: ВИЭМ, 1935.
Беклемишев В. Н. Основы сравнительной анатомии беспозвоночных. М.: Наука, 1964.
Беклемишев К• В. О возможных и осуществленных направлениях эволюции беспозвоночных.—Жури. общ. биол. 1974, т. 35, с. 209.
Белозерский А. Н. Нуклеиновые кислоты и их связь с эволюцией, филогенией и систематикой организмов.—В кн.: Материалы IV Междунар. био- физ. конгр. М.: Наука, 1973. 709 с.
Белозерский А. Н., Карпинская Р. С. Молекулярная биология и эволюционное учение. М.: Наука, 1970.
Белоусов Л. В. Проблема эмбрионального формообразования. М.: МГУ,
1971.
Белоусов Л. В., Гурвич А. А., Залкинд С. Я-, Каннигесер Н. Н. Александр Габрилович Гурвич, 1874—1954. М.: Наука, 1970.
Бендолл Дж. Мышцы, молекулы и движение. М.: Мир, 1970.
Берг Л. С. Труды по теории эволюции. М.: Наука, 1977.
Берестовский Г. Н„ Луневский В. 3. Электрооптические эффекты в мембранах (обзор).— В кн.: Биофизика мембраи. Каунас, 1971, ч. 1, с. 111.
Бернал Дж. Молекулярные матрицы живых систем.— В кн.: Происхождение предбиологических систем. М.: Мир, 1966. 76 с.
А. И. Опарин. Предисловие,—В кн.: Бернал Дж. Возникновение жизни. М.: Мир, 1969.
Бернхард С. Структура и функции ферментов. М.: Мир, 1971.
Бернштейн Н. А. Очередные проблемы физиологии активности.—В кн.: Проблемы кибернетики, 1961, т. 6.
Биологическое и социальное в развитии человека /Под ред. Б. Ф. Ломова и др. М.: Наука, 1977.
Блюменфельд Л. А., Темкин М. И. О возможном механизме образования АТФ в ходе окислительного фосфорилирования.— Биофизика, 1962, т. 7, с. 731.
Блюменфельд Л. А. Проблемы биологической физики. М.: Наука, 1974.
Блюменфельд Л. А. О некоторых физических аспектах внутриклеточной трансформации энергии.— Биофизика, 1976, т. 21, с. 946.
Бляхер Л. Я. Развитие представлений о материальной основе живых структур с древности до начала XX в.— В кн.: Физические и химические основы жизненных явлений (исторические очерки). М.: Изд-во АН СССР, 1963.
Богданов А. А. Современное состояние проблемы нуклеиново-белковых взаимодействий.— Усп. соврем, биол. 1974, т. 77, с. 48.
Болдырев А. А., Твердислов В. А. Молекулярная организация и механизмы функционирования Na-Hacoca. М.: ВИНИТИ. Сер, Итоги науки. Биофизика, 1978, т. 10.
33а. Бойченко Е. А., Опарин А. И. Участие поливалентных металлов в эволюции оксидоредуктаз.— Журн. эволюц. биохим. физиол., 1977, т. 13, с. 541.
Борисов В. В. Пространственная структура молекулы леггемоглобииа при разрешении 5А°. Канд. дне. М.: Ии-т кристаллографии АН СССР. 1975.
Браше Ж- Биохимическая эмбриология. М.: ИЛ, 1961.
Б ржевская О. Н., Каюшин Л. П., Неделина О. С. О существовании свободных радикалов в реакции гидролиза АТФ актомиозином.— Биофизика, 1966, т. 11. с. 213.
Брода Э. Эволюция биоэнергетических процессов. М.: Мир, 1978.
Брусков В. М., Полтев В. И. Узнавание ферментами комплементарных пар азотистых оснований и усиление специфичности взаимодействий в процессах матричного синтеза.— Докл. АН СССР, 1974, т, 219, с. 231.
Брусков В. И. Об узнавании оснований нуклеиновых кислот аминокислотами и пептидами с помощью водородных связей.— Молек. биол., 1975, т. 9, с. 304.
Букатина А. Е., Дещеревский В. И. Циклический механизм актомиозино- вой АТФ-азы.— Биофизика, 1972, т. 17, с. 738.
Букатина А. Е., Морозов В. Н., Шноль С. Э. Механохимическне преобразования в белках.— В ки.; Движение немышечных клеток и их компонентов. М.: Наука, 1976.
Букатина А. Е., Морозов В. Н., Шноль С. Э. Механохимическне преобразования в ферментах.— В кн.: Молекулярная и клеточная биофизика. М.: Наука, 1977.
Вавилов Н. И. Роль Дарвина в развитии биологических наук.— В кн.: Учение Дарвина н марксизм-ленинизм. М., 1932.
Вавилов Я. И. Закон гомологических рядов в наследственной изменчивости: Лнннеевский вид как система. М.: Наука, 1967.
Вазина А. А., Готберг М. И., Железная Л. А., Франк Г. М. Надмолекулярные структурысократительных белков.— В кн.: Биофизические основы и регуляция процесса мышечного сокращения. Пущино-иа-Оке, 1972. 204 с.
Ван-Ша шен. Скорость восходящего тока у древесных пород.— Докл. АН СССР, 1964, т. 156, с. 706.
Васильев Ю. М., Маленков А. Г. Клеточная поверхность и реакция клеток. М.: Медицина, 1968.
Вепринцев Б. Н. О связи электрической активности нервной клетки с синтезом г ней РНК и роль клеточной мембраны в регулировании биосинтеза РНК в клетке. Канд дис. Пущино-на-Оке: Ин-т биофизики АН СССР, 1970.
Вернадский В. И. Химическое строение биосферы Земли и ее окружения. М.: Наука, 1965. 316 с.
Виленкин Б. Я. Взаимодействующие популяции.— В ки.: Математическое моделирование в экологии. М.: Наука, 1978.
50а. Виленкин Б. Я-, Виленкина М. Н. Колебания воды, вызванные активностью морских дониых беспозвоночных.— Докл. АН СССР, 1971, т. 196, с. 214.
Винников Я- Л. Мехаиохимические и структурные основы функции рецепторов органов чувств.— В кн.: Молекулярная биология. Проблемы и перспективы. М.: Наука, 1964. 304 с.
Виноградова О. С. Гиппокамп и память. М.: Наука, 1975.
Возникновение органического вещества в солнечной системе/Под ред. Г. П. Вдовыкииа. М.: Мир, 1969.
Волькенштейн М. В. Физика ферментов. М.: Наука, 1967.
Волькенштейн М. В. Нарушения генетического кода.— Вестн. АПН. Научная мысль, 1968, вып. 4.
Волькенштейн М. В. Регуляция ферментами и регуляция ферментов.— Биофизика, 1970, т. 15, с. 215.
Вольтерра В. Математическая теория борьбы за существование. М.: Наука, 1976.
Воробьев В. И. Коиформационные превращения белковых комплексов как основа подвижности клеток и клеточных структур: докт. дис. М.: Ии-т цитологии АН СССР, 1975.
Воронцов Н. Н., Яблоков А. В. К 70-летию Н. В. Тимофеева-Ресовско- го.— Бюл. МОИП. Отд-ние биол., 1970, № 5.
Вулдридж Д. Механизмы мозга. М.: Мир, 1965.
Газиев А. И. ДНК-лигазы.— Усп. соврем, биол., 1974, т. 78, с. 171.
Гахова Э. Н. Трансмембраниые ионные потоки как возможный механизм регулирования синтеза РНК и белка. Канд. дис. Пущино-на-Оке: Ии-т биофизики АН СССР, 1978.
Гейз С. Образование нервных связей. М.: Мир, 1972.
Гейльбрун Л. Динамика живой протоплазмы. М.: ИЛ, 1957.
Геодакян В. А. Дифференциальная смертность и норма реакции мужского и женского пола. Онтогенетическая и филогенетическая пластичность.— Журн. общ. биол., 1974, т. 35, с. 376.
Гиляров М. С. Закономерности приспособления членистоногих к жизни на суше. М.: Наука, 1970.
Гиляров М. С. Закономерности и направления филогенеза.— Журн. общ. биол., 1970, т. 31, с. 31.
Гинецикский А. Г. Физиологические механизмы водносолевого равновесия. М.: Наука, 1964.
Гительзон И. И., Чумакова Р. И. Биохимические основы биолюминесценции.— Усп. соврем, биол., 1975, т. 3, с. 79.
Голенкин М. И. Победители в борьбе за существование. М.: Учпедгиз, 1959.
Гончарова Н. В., Евстигнеев В. Б. Образование АТФ в модельной системе при участии каталазы и H202.— Докл. АН СССР, 1975, т. 226, с. 970.
Гончарова Н. В., Евстигнеев В. Б. О фотофэсфорилироваиии сенсибилизированном хлорофиллом в адсорбированном состоянии.— Биохимия, 1975, т. 40, с. 622.
Гончарова Н. В., Евстигнеев В. Б. Фосфорилирование при кислотио-ос- новном переходе в модельной системе, содержащей хлорофилл.— Докл. АН СССР, 1977, т. 236, с. 220.
Гончарова Н. В., Евстигнеев В. Б. Фотофосооршшрование, сенсибилизированное хлорофиллами айв, феофитином и Р-каротином в модельной системе,—Биохимия, 1977, т. 42, с. 963.
Гохлернер Г. Б. Развитие аэробной жизни и проблемы клеточной эволюции.— Природа 1977. № 6.
Гурвич А. Г. Теория биологического поля. М.: Сов. наука, 1944.
Гурвич А., Гурвич Л. Введение в учение о митогенезе. М.: Изд-во АМН СССР, 1948.
Гурвич А. А. Проблема митогенетического излучения как аспект молекулярной биологии. Л.: Медицина, 1968.
Гурский Г. В., Туманян В. Г., Заседателев А. С., Жузе А. А., Гроховский С. Л., Готтих Б. П. Код, управляющий специфическим связыванием регуляторных белков с ДНК, и структура стереоспецифичных участков регуляторных белков.— Молек. биол., 1975, т. 9, с. 635.
Дадингтон К.. Эволюционная ботаника. М.: Мир, 1972.
Движение и поведение одноклеточных животных. Л.: Наука, 1978.
Движение немышечиых клеток и их компонентов. Л.: Наука, 1977.
82а. Дегерменджи А. Г. «Надежность» процесса микроэволюции стабильных и флуктуирующих популяций в открытых системах.— Журн. общ. биол., 1977, т. 38, с. 423.
Дельбрюк М. Обновленный взгляд физика иа биологию.— УФН, 1971, т. 105, 393; Волькенштейн М. В. Несколько слов о лекции Дельбрюка.— УФН, 1971, т. 10, с. 401.
Дещеревский В. И. Две модели мышечного сокращения.— Биофизика,
т. 13. с. 928.
Дещеревский В. И. Теория сокращения асинхронных мышц.— Биофизика,
т. 15, с. 112.
Дещеревский В. И. Математические модели мышечного сокращения. М.: Наука, 1977.
Дещеревский В. И., Букатина А. Е., Сидоренко Н. П. Исследование меха нохимического цикла по ферментативной кинетике актомиозииовых систем.— В кн.: Молекулярная и клеточная биофизика. М.: Наука, 1977.
Джамусова Т. А. Тепловая контрактура и необратимая потеря возбудимости мышц в связи с теплоустойчивостью мышечной ткани.— В кн.: Теплоустойчивость клеток животных. М.: Наука, 1965. 61 с.
Дженкс В. П. Катализ в химии и энзимологии. М.: Мир, 1972.
Диксон М., Уэбб Э. Ферменты. М.: Мир, 1966.
Догель В. А. Зоология беспозвоночных. М.: Учпедгиз, 1939.
Догель В. А. Общая протистология. М.: Сов. наука, 1961.
Дубинин Н. П., Шевченко Ю. Г. Некоторые вопросы биосоциальной природы человека. М.: Наука, 1976.
Евреинова Т. Н. Концентрирование веществ и действие ферментов в коа- церватах. М.: Наука, 1966.
Евстигнеев В. Б. О возможной роли кислотно-основного равновесия как фактора регуляции фотохимических реакций хлорофилла in vivo и in vitro.— Биофизика, 1970, т. 15, с. 239.
Есипова Н. Г. О характере водородных связей в коллагене.— Биофизика, 1957, т. 2, с. 461.
Есипова Н. Г. Условия существования и факторы стабилизации структур коллагенового типа. Канд. дне. Пущиио-на-Оке: Ии-т биофизики АН СССР, 1974.
Есипова Н. Г., Макаров А. А. и др. К проблеме тепловой устойчивости решетки кристаллов белков.— Докл. АН СССР, 1976, т. 227, с. 981.
Есипова Н. Г., Макаров А. А., Монаселидзе Д. Р., Мгеладзе Г. Н., Волкова Г. А. Физическое состояние белков в кристаллах. III. О структуре кривых теплового нагрева.— Биофизика, 1976, т. 21, с. 615.
Есипова Н. Г. О роли воды в биополимерах в связи с проблемой струк- турообразоваиия в биологических системах.— Биофизика, 1978, т. 23.
Жаботинский М. А., Заикин А. Н. Пространственные эффекты в автоколебательной химической системе.— В кн.: Колебательные процессы в биологических и химических системах. Пущино-на-Оке, 1971, т. 2. 279 с.
Жаботинский А. М. Концентрационные автоколебания. М.: Наука, 1974.
Жизнь животных. М.: Просвещение, 1968. Т. 1, кн. 2.
Заалишвили М. М. Физико-химические основы мышечной деятельности. Тбилиси: Мецниереба, 1971.
Завадский К.. М. Вид и видообразование. М.: Наука, 1968.
105а. Завадский К. М. Развитие эволюционной теории после Дарвина. Л.: Наука, 1973.
Завадский К- М., Георгиевский А. Б. К оценке эволюционных взглядов Л. С. Берга. В кн. [21].
Завадский К М., Колчинский Э. И. Эволюция эволюции. М.: Наука, 1977.
Заварзин А. А. Очерки по эволюционной гистологии нервной системы.— Избр. труды. М.: Изд-во АН СССР, 1950, т. 3.
Закономерности прогрессивной эволюции. Л.: Наука, 1972.
Захваткин А. А. Сравнительная эмбриология низших беспозвоночных: (Источники и пути формирования индивидуального развития многоклеточных). М.: Сов. наука, 1949.
Зацепина Г. Н. Свойства и структура воды. М.: МГУ, 1974,
Зенкевич Л. А. Очерки эволюции аппарата движения животных.—I. Некоторые общие аспекты эволюции движения животных.— Журн. общ. биол. 1944, т. 5, с. 129; Он же. III. Конечности насекомых.— Труды Л> нингр. об-ва естествоиспыт. Секция зоол. 1957, т. 73, с. 19; Он же. Гидравлический способ движения животных.— Природа, 1964, № 6, с. 89.
ИЗ. Иберт Дж. Взаимодействующие системы в развитии. М.: Мир, 1968.
Иванов А. В. Происхождение многоклеточных животных: Филогенетические очерки. М.: Наука, 1968.
Иванов А. В. Trichoplax adhaereus — фагоцителлообразиое животиое.— Зоол. журн., 1973, т. 52, с. 1117.
Иванов В. И. Изучение механизма действия фермента аспартаттрансамн- иазы оптическими методами. Каид. дис. М.: МГУ им. Ломоносова, 1968.
Иванов К■ П. Биоэнергетика и температурный гомеостазис. М.: Наука,
1972.
Иванцов Н. А. Факторы эволюции. М.: Госиздат, 1923.
Ильенков Э. В. Диалектическая логика. М.: Политиздат, 1974.
Ильичева И. А„ Кистер А. Э., Дашевский В. Г., Есипова Н. Г., Туманян В. Г. «В»-форма как абсолютный энергетический минимум ДНК- Энтропийная природа А—Б-перехода.— Биофизика, 1978, т. 23.
Ильюченок Р. Ю. Фармакология поведения и памяти. М.: Наука, 1972.
Иорданский Н. Н. Неравномерность темпов макроэволюции и ключевые ароморфозы.— Природа, 1977, № 6, с. 36.
Казначеев В. П., Шурин С. П., Михайлова Л. П., Игнатович Н. В. О межклеточных дистантных взаимодействиях в системе двух тканевых культур, связанных оптическим контактом.— В кн.: Сверхслабые свечения в биологии. М.: Наука, 1972.
Кайданов Л. 3. К вопросу о роли поведения как фактора микроэволюции.— В кн.: Исследования по генетике. Л.: ЛГУ, 1967, вып. 3, с. 27.
Калиниченко Л. П., Христова М. Л., Шноль С. Э. Влияние D2O, глицерина, алифатических спиртов и возможная роль гидрофобных взаимодействий в транспорте электронов в митохондриях.— Биофизика, 1967, т. 12, с. 284.
Кальвин М. Химическая эволюция. Молекулярная эволюция, ведущая к возникновению живых систем на Земле и на других планетах. М.: Мир,
1971.
127а. Камшилов М. М. Фенотип и генотип в эволюции.— Проблемы эволюции,
т. 2, с. 28.
1276. Камшилов М. М. Эволюция биосферы. Л.: Наука, 1974.
Карев Г. П., Ляпунов А. А., Тресков С. А. О детерминированном и вероятностном подходах к эволюционным задачам в математической теории популяций.— Проблемы эволюции, 1975, т. 4, с. 5.
Карпинская Р. С. Материалистическая диалектика о закономерностях развития органического мира. М.: Знание, 1975.
Катц В. Нерв, Мышца, Синапс. М.: Мир, 1968.
131а. Каюшин Л. П., Людковская Р. Г. Изменение объемио-упругих свойств нерва при проведении возбуждения и связь их с биопотенциалами.— Труды Ин-та биофизики АН СССР, 1955, т. 2, с. 40.
1316. Каюшин Л. П. и др. К вопросу о передаче энергии гидролиза АТФ сократительному белку.— Биофизика, 1961, т. 6, с. 20.
Кеньон Д., Стейман Г. Биохимическое предопределение. М.: Мир, 1972.
Киселев JI. Л. В кн.: Молекулярные основы биосинтеза белков. М.: Наука, 1971, с. 7.
Клетка и температура среды.— Труды Междунар. симпоз. по цитоэкологии. М.: Наука, 1964.
Клотц И. Энергетика биохимических реакций. М.: Мир, 1970.
Кобозев Н. И. Общие принципы каталитической активности. 2. Принципы агравации в катализе.— Учен. зап. МГУ, 1946, т. 86, № 13.
Кобозев Н. И. О валентном и энергетическом механизме катализа.— В ки.: Катализ в высшей школе. М.: МГУ, 1962, вып. 1, ч. 1, с. 43.
Кобозев Н. И. Адсорбционные катализаторы и теория активных центров.— В кн.: Современные проблемы физической химии. М.: МГУ, 1968, т. 3, с. 3.
Ковров Б. Г., Косолапова Л. Г. Дискретная математическая модель процесса видообразования.— Журн. общ. биол., 1977, т. 38, с. 359.
Кокшайский Н. В. Очерк биологической аэро- и гидродинамики (полег и плаванье животных). М.: Наука, 1974.
Колли А. А. Процессы брожения. М., 1876.
Колли А. А. Микроорганизмы с химической точки зрения: (Речь, произнесенная 1.1.1893 г. в общем собрании 9-го съезда естествоиспытателей и врачей). М., 1894.
Кольтовер В. К-, Блюменфельд Л. А. Термические конформационные переходы в электрои-переносящих биологических мембранах.— Биофизика, 1973, т. 18, с. 827.
Кольцов Н. К■ Организация клетки.: Сборник эксперим. исслед. статей и речей, 1903—1935 гг. М.: Биомедгиз, 1936.
Комиссаров Г. Г., Шумов Ю. С. О возможности фотоэлектролиза воды в моделях, содержащих фотосинтетические пигменты, и в зеленом листе.— Докл. АН СССР, 1968, т. 182, с. 1226.
Кондрашова М. Н. Метаболические состояния митохондрий и основные физиологические состояния живой ткани.— В кн.: Свойства и функции макромолекул и макромолекулярных систем. М.: Наука, 1969, с. 135.
146а. Кондрашова М. Н. Регуляция дыхания митохондрий при усиливающемся воздействии иа клетку.— Биофизика, 1970, т. 15.
Кондрашова М. Н., Миронова Г. Д. Необходимость кислорода для фос- форилирования АДФ в условиях цианидного блока.— Биохимия, 1971, т. 36, с. 864.
Конев С. В., Аксенцев С. П., Черницкий Е. А. Кооперативные переходы белков в клетке. Минск: Наука и техника, 1970.
Коникова А. С., Крицман М. Г., Самарина 0. П. Исследование условий включения аминокислот в «индивидуальные» белки и белковые комплексы.— Биохимия, 1954, т. 19, с. 440.
Коржуев П. А. Эволюция дыхательной функции крови. М.: Изд-во АП СССР, 1949.
Корниенко И. А., Маслова Г. М., Гохблит И. И. О роли бурого жира в химической терморегуляции иа разных стадиях онтогенеза крыс.— Журн. эволюц. биохим. и физиол., 1971, т. 7, с. 385.
Котельникова А. В. Эволюция механизмов превращения химической энергии у живых организмов.— В кн.: Материалы 1-го Всесоюз. биохим. съезда: Тез. докл. М.: Изд-во АН СССР, 1963, т. 1, с. 5.
Котельникова А. В., Соломатина В. В. Исследование способности к электролитическому восстановлению некоторых нуклеотидов.— Биохимия
т. 30, с. 816.
Кочетков Н. К■ и др. Органическая химия нуклеиновых кислот. М.: Химия, 1970.
Крамер П. И., Козловский Т. Т. Физиология древесных растений. М.: Гос- лесбумиздат, 1963.
Красновский А. А. Преобразование энергии при фотосинтезе. Молекулярные механизмы. М.: Кчука, 1974.
Кребс Г., Корнберг Г. Превращения энергии в живых системах. М.: ИЛ, 1959.
Кремлев И. Н. Потенциал-зависимые реакции и регуляция мембранной проводимости. Автореф. канд. дис. М.: МГУ нм. Ломоносова, 1976.
Крушинский Л. В. и др. Экстраполяционный рефлекс и его роль в эволюции поведения животных.— В кн.: Проблемы кибернетики. М.: Наука, 1966, т. 16, с. 91.
Крушинский Л. В. Сравнительно физиологические аспекты изучения сложных форм поведения животных.— Журн. эволюц. биохим. и физиол., 1973, т. 9, с. 278.
Крушинский Л. В. Биологические основы рассудочной деятельности. М.: МГУ, 1977.
Кулаев И. С. Биохимия высокомолекулярных полифосфатов. М.: МГУ, 1975.
Курсанов Л. И. и др. Определитель низших растений. Водоросли. М.: Сов. наука, 1953, т. 1.
Лабас Ю. А. К проблеме происхождения и функции биолюминесценций у простейших и низших.— В кн.: Биологическая спектрофотометрия и фотоактинометрия. Красноярск, 1973, с. 59.
Лабас Ю. А., Маковский В. С. Биопотенциалы, связанные с мерцательным движением, как начальный этап эволюции возбудимых структур,—В кн.: Немышечные формы подвижности/Под ред. Г. М. Франка. Пущино-на- Оке: НЦБИ АН СССР, 1976.
Лазарев Ю. А., Есипова Н. Г. Спектральные исследования структурной упорядоченности белков коллагеиового типа.— Докл. АН СССР, 1973, т. 209, с. 478.
Леб Ж■ Динамика живого вещества. Одесса, 1910.
Леб Ж. Организм как целое. М.; Л., 1926.
Леб Ж■ Вынужденные движения, тропизмы и поведение животных.
Лебедев А. И., Киселев Б. А., Макаров А. Д. Исследования восстановления АДФ в нейтральной области pH электрохимическим методом.— Докл. АН СССР, 1971, т. 201, с. 1489.
Лев А. А. Ионная избирательность клеточных мембран. М.: Наука, 1975.
Лев А. А. Моделирование ионной избирательности клеточных мембран. М.: Наука, 1976.
Левонтин Р. Генетические основы эволюции. М.: Мир, 1978.
Лем С. Сумма технологии. М.: Мир, 1968.
Ленинджер А. Связанные с дыханием механохимические изменения в митохондриях.— В кн.: Горизонты биохимии. М.: Мир, 1964. 326 с.
Ленинджер А. Биохимия. М.: Мир, 1974.
Либерман Е. А. и др. Транспорт ионов и электрический потенциал митохондриальных мембран.— Биохимия, 1969, т. 34, с. 1083.
Либерман Е. А. Мембраны (ионная проницаемость, возбудимость, управление).— Биофизика, 1970, т. 15, с. 278.
179а. Лифшиц И. М. Некоторые вопросы статистической теории биополимеров,— ЖЭТФ, 1968, т. 55, с. 2408.
1796. Лифшиц И. М. Диаграмма состояний полимерной глобулы и проблема ее пространственной структуры—ЖЭТФ, 1973, т. 65, с. 2399.
180. Лишко В. К Натриевый насос биологических мембран. Киев; Наукова думка, 1977.
181а. Лобашев М. Е. Сигнальная наследственность.— В кн.: Исследование по генетике. Л.: ЛГУ, 1961, вып. 1, с. 3.
1816. Лобышев В. И., Калиниченко Л. П. Изотопные эффекты D20 в биологических системах. М.: Наука, 1978.
182. Лопашов Г. В. Механизмы развития зачатков глаз в эмбриогенезе позвоночных. М.: Изд-во АН СССР, 1960.
.183. Лукин Е. И. Проблема типов животного мира и теория ароморфозов.— Проблемы эволюции, 1972, т. 2, с. 124.
Я84. Любимова М. Н., Энгельгардт В. А. Адепозиитрифосфатаза н миозин мышц,—Биохимия, 1939, т. 4, с. 716.
185а. Любищев А. А. Проблемы систематики.— Проблемы эволюции, 19681,. т. 1, с. 7.
1856. Любищев А. А. К логике систематики.— Проблемы эволюции, 1972, т. 2, с. 45.
185в. Любищев А. А. О постулатах современного селектогенеза.— Проблемы эволюции, 1973, т. 3, с. 31.
185г. Любищев А. А. К классификации эволюционных теорий.— Проблемы эволюции, 1975, т. 4, с. 206.
185д. Любищев А. А. Редукционизм и развитие морфологии и систематики.— Журн. общ. биол., 1977, т. 38, с. 245.
186а. Людковская Р. Г., Каламкарова М. Б. О механизме усиления сократимости глицеринизироваииых мышечных волокон в присутствии красителей,—В кн.: Биофизика клетки. М.: Наука, 1965, с. 115.
1866. Людковская Р. Г., Емельянов В. Б., Лемажихин Б. К■ Исследование оптических свойств гигантского аксона кальмара в покое и в разные фазы возбуждения.— Цитология, 1965, т. 7, с. 520.
Маевский П. Ф. Флора средней полосы Европейской части СССР. М.: Колос, 1964.
Мазохин-Поршняков Г. А. Информационная организация и механизмы поведения насекомых.— Журн. общ. биол., 1975, т. 36, с. 8.
Майр Э. Зоологический вид и эволюция. М.: Мир, 1968.
189а. Майр Э. Популяция, виды и эволюция. М.: Мир, 1974.
Маленков А. Г. Иоиный гомеостаз и адгезивные свойства нормальных и опухолевых клеток. Докт. дис. Пущиио-на-Оке: Ии-т биофизики АН СССР. 1974.
Маленков А. Г., Модянова Е. А. Прочность межклеточных контактов и пролиферативный пул в мышиных гепатомах.— Цитология, 1968, т. 710, с. 1088.
Маленков Г. Г. Кто кого?—Химия и жизиь, 1974, № 10, с. 3—9.
Малиновский А. А. Пути теоретической биологии. М.: Знание, 1969.
Малиновский А. А., Порошенко Г. Предисловие к русскому изданию.— В кн.: Шрёдингер Э. Что такое жизнь? М.: Атомиздат, 1972.
Манойлов С. Е., Вовси Б. А., Полосова Р. Г., Сидорова Н. Д. Влияние каталазы на процессы сопряженного окислительного фосфорилирования и на состояние аденнловой системы в печени белых крыс.— Биохимия, 1966, т. 31, с. 613.
Манойлов С. Е. Первичные механизмы биологического действия проникающей радиации. М.: Медицина, 1969, с. 46—56.
196а. Манойлов С. Е. О биологической роли каталазы.— Труды Ленингр. хим.- фармакол. ин-та, 1969.
Манойлов С. Е. Биохимические основы злокачественного роста. М.: Медицина, 1970.
Маркин В. С. Механизм проводимости искусственных фосфолипидных мембран в присутствии переносчиков ионов.— Биофизика, 1969, т. 14, с. 256.
Мартынов Л. А. Оптико-механические свойства и деформации клеточной оболочки ацетабулярий.— Биофизика, 1973, т. 18, с. 935.
Матвеев Б. С. Происхождение и пути эволюции позвоночных животных по современным данным.— Проблемы эволюции, 1968, т. 1, с. 82.
Машанский В. Ф„ Лабас Ю. А., Снигеревская Е. С. Природа фотогенных гранул и пусковые механизмы активируемой кальцием биолюминесценции у гребневиков.— Цитология, 1974, т. 16, с. 1459.
Медников Б. М. О реальности высших систематических категорий позвоночных животных.— Журн. общ. биол., 1974, т. 34, с. 516.
Медников Б. М. Дарвинизм в XX в. М.: Сов. Россия, 1975.
Медников Б. М. Дивергенция геномов и некоторые вопросы эволюционной, теории. Докт. дис. М.: Ин-т биол. развития им. Н. К. Кольцова, 1977.
Медников Б. М., Меншуткин В. В. Опыт моделирования эволюции нуклеотидных последовательностей на ЭВМ.— Журн. общ. биол., 1977, т. 37„ с. 198.
Меншуткин В. В. Опыт имитации эволюционного процесса на вычислительной машине.— Журн. эволюц. биохим. и физиол., 1977, т. 13, с. 545.
Мейен С. В., Соколов Б. С., Шрейдер Ю. А. Неклассическая биология, феномен Любищева.— Вести. АН СССР, 1977, т. 10.
Микельсаар X. Н. Концепция археореляции аминокислот о возникнове- нни жизни и генетического кода.— Журн. общ. биол., 1975, т. 36, с. 3.
Моисеева Л. Н., Слободская В. П., Пасынский А. Г. Сопряжение процессов диффузии и химических реакций и эволюция открытых биохимических систем.— В кн.: Абиогенез и начальные стадии эволюции жнзни. М,; Л.: Наука, 1968, с. 56.
Молчанов А. М. Термодинамика и эволюция.— В кн.: Колебательные пре- цессы в биологических и химических системах. М.: Наука, 1967, 292 с.
Молчанов А. М. Возможная роль колебательных процессов в эволюции.— В кн.: Колебательные процессы в биологических и химических системах. М.: Наука, 1967. 274 с.
Моно Ж, Жакоб Ф. Телеономические механизмы в клеточном метаболизме, росте и дифференцировке.— В кн.: Регуляторные механизмы клетки. М.: Мир, 1964. 477 с.
Мора П. Несостоятельность вероятностного подхода.— В кн.: Происхождение предбиологических систем. М.: Мир, 1966. 47 с.
Моран П. Статистические процессы эволюционной теории. М.: Наука,
1973.
Морозов В. Н., Букатина А. Е., Шноль С. Э. Механохимическпе преобразования в белках.— В кн.: Движение немышечных клеток и их компонентов. Л.: Наука. 1978.
Москона А. Как клетки объединяются друг с другом.— В кн.: Живая клетка. М.: Мир, 1966. 111 с.
Мосолов Л. Н., Монахова М. А., Нилзова Л. Р. О возможном участии центромерного кольца в формировании ядер половых клеток.— Цитология и генетика, 1976, т. 10, с. 8.
Мосолов В. В. Протеолитические ферменты. М.: Наука, 1971.
Мостовников В. А., Хохлов И. В. Взаимодействие клеток человека с помощью электромагнитных волн оптического диапазона: Препринт Л» 128. Минск: Ии-т физики АН БССР, 1977.
Мэгун Г. Бодрствующий мозг. М.: ИЛ, 1961.
Насонов Д. Н. Местная реакция протоплазмы и распространяющееся возбуждение. М.: Изд-во АН СССР, 1962.
Насонов Д. Н„ Александров В. Я. Реакция живого вещества на внешние воздействия. М.: Изд-во АН СССР, 1940.
Нейштадт М. И. Определитель растений средней полосы европейской части СССР. М.: Учпедгиз, 1963.
Николаев Л. А. Биокатализаторы и их модели. М.: Высшая школа, 1968.
Нуцубидзе М. А. Эмоциональная и замыкательная функция лимбической системы. Тбилиси: Мецниереба, 1969.
Овчинников Ю. А. Макроциклические депсипептидные антибиотики и транспорт ионов через мембраны.—Усп. соврем, биол., 1974, т. 77, с. 103.
Овчинников Ю. А., Иванов В. Т., Шкроб А. М. Мембрано-активные комп- лексоны. М.: Наука, 1974.
Опарин А. И. Происхождение жизни. М.: Моск. рабочий, 1924.
Опарин А. И. Жизнь, ее природа, происхождение и развитие. М.: На\ка, 1959.
'229. Орбели Л. А. Вопросы высшей нервной деятельности. М.: Изд-во АН СССР, 1949.
Основные аспекты геохимии нефти. М.: Недра, 1970.
Остроумов Г. А., Серавин Л. Н. К вопросу о гидромеханике плавания водных животных. 1. Гидромеханические модели, объясняющие движение инфузорий.— Вестн. ЛГУ, 1974, № 9.
.232. Патти X. Наследственная упорядоченность в примитивных химических системах.— В кн.: Происхождение предбиологических систем. М.: Мир, 1969. 385 с.
'233. Паулинг Л. Природа химической связи. М.: Гос. хим. изд., 1947.
Первухина Н. В. Проблемы морфологии и биологии цветка. М.: Наука, 1970
Печуркин Н. С., Терское И. А. Автоселекциониые процессы в непрерывной культуре микроорганизмов. М.: Наука, 1973.
Поглазов Б. Ф. Сократительные системы фагов в [82].
Поглазов Б. Ф. Самосборка биологических структур. М.: Наука, 1970.
Полторак О. М., Чухрай Е. С. Физико-химические основы ферментативного катализа. М.: Высшая школа, 1971.
Пондер Э. Цит. по кн.: Теоретическая и математическая биология. М.: Мир, 1968. 155 с.
Проблема возникновения и сущности жизни/Под ред. А. И. Опарина. М.: Наука, 1973.
Проблемы эволюции/Под ред. Н. Н. Воронцова. Новосибирск: Наука, 1968—1975.
Происхождение нефти и газа и формирование их месторождений. М.: Недра, 1972.
Происхождение предбиологических систем/Под ред. А. И. Опарина. М.: Мир, 1966.
243а. Ратнер В. А. Математическая теория эволюции менделевских популяций.— Проблемы эволюции, 1973, т. 3, с. 151.
Рогинский С. 3., Шноль С. Э. Изотопы в биохимии. М.: Изд-во АН СССР, 1963.
Романовский Ю. М., Степанова Н. В., Чернавский Д. С. Математическое моделирование в биофизике. М.: Наука, 1975.
Рубинштейн Д. Л. Общая физиология. М.: Медгиз, 1947.
Рубинштейн Д. Л. Выступление по докладу Д. Н. Насонова иа конференции в Гаграх в 1947 г.: Гагрские беседы. Тбилиси, 1949, с. 34.
Рубинштейн Д. Л., Туточкина Л. БГрищенко Е. Д. Фотодинамическое действие эритрозина на сократительные белки скелетных мышщ.— Докл. АН СССР, 1951, т. 76, с. 243.
Руденко А. П. Теория саморазвития открытых каталитических систем. М.: МГУ, 1969.
Руттен М. Происхождение жизни. М.: Мир, 1973.
Сабинин Д. А. Физиологические основы питания растений. М.: Изд-во АН СССР, 1955.
Савостьянов Г. А. О некоторых элементарных актах и законах биологического развития. Элементы структурной биологии.— Журн. общ. биол.,
т. 38, с. 167.
Сарвазян А. П., Пасечник В. П., Шноль С. Э. Низкая скорость звука в гелях и протоплазматических структурах. Возможное биологическое значение этого явления.— Биофизика, 1968, т. 13, с. 587.
Северцов А. Н. Эволюция и психика. М.: Изд-во братьев Сабашниковых. 1922.
Северцов А. Н. Главные направления эволюционного процесса. М.: Био- медгнз, 1934.
Северцов А. Н. Морфологические закономерности эволюции. М.: Изд-во АН СССР, 1939.
Сельков Е. Е. Два альтернативных автоколебательных стационарных состояния в метаболизме тиолов—два альтернативных типа размножения клеток: нормальный и злокачественный.— Биофизика, 1970, т. 15, с. 1065.
Сельков Е. Е. Исследование механизма гликолитических колебаний. Релаксационная модель.— В кн.: Математические модели биологических систем. М.: Наука, 1971, с. 5.
Серавин Л. Н. Двигательные системы простейших. М.: Наука, 1967.
Серавин Л. Н. Некоторые гидродинамические аспекты движения ресничных дифузорий.— Вести. ЛГУ, 1870, К» 21. 41 с.
Серебровская К■ Б. Коацерваты и протоплазма. М.: Наука, 1971.
Сериков А. 3. Фотодииамические эффекты в органных культурах.— Журн. общ. биол., 1978, т. 39, с. 304.
Сеченов И. М. Рефлексы головного мозга. Избр. филос. и психол. произв. М.: ОГИЗ, 1947.
Симпсон Дж. Г. Темпы и формы эволюции. М.: ИЛ, 1948.
Скулачев В. П. Роль адеииннуклеотидов в фосфорилирующем дыхании и физиологические функции окислительных процессов.— Усп. соврем, биол., 1964, т. 6, с. 180.
Скулачев В. П. Аккумуляция энергии в клетке. М.: Наука, 1969.
Скулачев В. П. Трансформация энергии в биомембранах. М.: Наука, 1972.
Скулачев В. П. Механизм окислительного фосфорилирования и некоторые принципы биоэнергетики.— Усп. соврем, биол., 1974, т. 77, с. 125.
Скулачев В. П. АТФ и трансмембранный потенциал ионов водорода —две конвертируемые и транспортабельные формы энергии в живой клетке.— Усп. соврем, биол., 1977, т. 84, с. 165.
Скулачев В. П., Денисович Л. И. Окислительно-восстановительные свойства производных аденина.— Биохимия, 1966, т. 31, с. 132.
Скулачев В. П., Козлов И. А. Протонные аденозинтрифосфатазы. Молекулярные биологические генераторы тока. М.: Наука, 1977.
Слейгер Р. Водный режим растений. М.: Мир, 1970.
Слоним А. Д. Экологическая физиология животных. М.: Высшая школа-, 1971.
273а. Современныйдетерминизм. Законы природы. М.: Мысль, 1973.
Соннеборн Т. М. Морфогенез у ресничных инфузорий и его отношение к клеточному морфогенезу в целом.— Цитология, 1974, т. 16, с. 1069.
Спирин А. С. О структурных основах функционирования рибосом.— Усп. соврем, биол., 1974, т. 77, с. 155.
Спирин А. С., Гаврилова Л. П. Рибосома. М.: Наука, 1971.
Спитковский Д. М. Об одной возможной причине движения протоплазмы и некоторые следствия тейно-химического принципа.— Биофизика, 1966, т. 11, с. 1022.
277а. Степанов В. Н. Мировой океан. М.: Знание, 1974.
Сургучев А. П. Для чего нужна рибосомная РНК?—Природа, 1977, № 7, с. 128.
Сыркин Я■ К-, Дяткина М. Е. Химическая связь и строение молекул. М.: Госхимиздат, 1946.
Съюорд А. Ч. Века и растения, М., 1936.
Тасаки И. Нервное возбуждение. М.: Мир, 1971.
Тахтаджян А. Л. Высшие растения. М.: Изд-во АН СССР, 1956, т. 1.
Тахтаджян А. Л. Происхождение покрытосеменных растений. М.: Изд-во АН СССР, 1961.
Тахтаджян А. Л. Основы эволюционной морфологии покрытосеменных. М.: Наука, 1964.
Твердислов В. А. Параметрический насос и активный транспорт ионов в клетках. Канд. дис. М.: МГУ им. Ломоносова, 1973.
Темкин М. И. Кинетика стационарных сложных реакций.— В ки.: Механизм и кинетика сложных каталитических реакций. М.: Наука, 1970, с. 57.
Теренин А. Н. Фотоника молекул красителей. М.: Наука, 1967.
Тимофеев-Ресовский Н. В. О механизмах авторепродукции элементарных клеточных структур.— Цитология, 1960, т. 2, с. 45.
Тимофеев-Ресовский Н. В., Воронцов Н. Н., Яблоков А. В. Краткий очерк теории эволюции. М.: Наука, 1969.
Тимофеев-Ресовский Н. В., Яблоков А. В. Микроэволюция, элементарные явления, материал и факторы эволюционного процесса. М.: Знание, 1974.
Токин Б. П. Общая эмбриология. М.: Высшая школа, 1970.
Третьяк Н. Н., Костенко М. А. Зависимость синтеза РНК и образования отростков у изолированных нейронов легочных моллюсков от ионного состава и осмотической силы питательной среды.— Цитология, 1978, т. 20 с. 643.
Тюньков И. В. Колебания АТФ-азной активности в растворах актомиози- на.— В кн.: Материалы конф. Всесоюз. хим. об-ва Д. И. Менделеева. Иркутск, 1967, с. 175.
Уголев А. М. Мембранное пищеварение. Полисубстратные процессы, организация и регуляция. Л.: Наука, 1972.
.295. Уоддингтон К. Организаторы и гены. М.: ИЛ, 1947.
295а. Уоддингтон К.. Морфогенез и генетика. М.: Мир, 1964.
296. Уоддингтон К. Основные биологические концепции.— В кн.: На пути к теоретической биологии. М.: Мир, 1970, т. 1, с. 11.
.297. Уотсон Дж. Молекулярная биология гена. М.: Мир, 1967.
298. Уотсон Дж. Двойная спираль. М.: Мир, 1969.
.299. Ухтомский А. А. Доминанта. М.: Наука, 1966.
-300. Ушаков Б. П. Современное состояние вопроса о механизме теплового повреждения и причинах изменения теплоустойчивости клеток.— В кн.: Теплоустойчивость клеток животных. М.: Наука, 1965, с. 5.
•301. Ушаков В. Б. Теплоустойчивость возбудимой и сократимой систем мышечного волокна.— В кн.: Теплоустойчивость клеток животных. М.: Наука, 1965, с. 55.
Уэстли Дж. Ферментативный катализ. М.: Мир, 1972.
Ферстер Г. О самоорганизующихся системах и их окружений.— В кн.: Самоорганизующиеся системы. М.: Мир, 1964, с. 113.
■304. Филипченко Ю. А. Эволюционная идея в биологии. М.: Наука, 1977.
Фогель Л., Оуэнс А., Уолш М. Искусственный интеллект и эволюционное моделирование. М.: Мир, 1969.
Фокс К■ Молекулярная эволюция и возникновение жизни. М.: Мио, 1975.
Франк Г. М. Клеточные структуры и функция клетки.— Биофизика, 1970, т. 15, с. 298.
307а. Франк Г. М. Проблемы изучения биологических мембран.— Вестн. АН СССР, 1970, т. 9, с. 36.
•308. Фрисман Е. Я; Шапиро И. П. Избранные математические модели дивергентной эволюции популяций. М.: Наука, 1977.
Фролов И. Т. Генетика и диалектика. М.: Наука, 1968.
■309а. Хенох М. А., Николаева М. В. Абиогенный фотохимический синтез аминокислот в водной среде, содержащей углеводы и нитраты.—Журн. эвол. биохим. физиол., 1977, т. 13, с. 105.
Химические факторы регуляции активности и биосинтеза ферментов/Под ред. В. Н. Ореховича. М.: Медицина, 1969.
Хлебович В. В. Критическая соленость биологических процессов. М.: Наука, 1974.
Хлебович В. В., Орфеев Ю. В. О вероятном экологическом стимуле формирования протозойиой колониальности и становления многоклеточности.— В кн.: Проблемы эволюции, 1975, т. 4, с. 11.
Ходжкин А. Нервный импульс. М.: Мир, 1965.
Ходоров Б. И. Проблема возбудимости. М.: Медицина, 1969.
Холдейн Дж. Возникновение жизни [в 19], с 295.
Холдейн Дж. Факторы эволюции. М.: Биомедгиз, 1935.
Хориути Дж. Как найти кинетическое уравнение обратной реакции?— В кн.: Проблемы физической химии. М.: Госхимиздат, 1959, вып. 2, с. 39.
Хургин Ю. И., Чернавский Д. С., Шноль С. Э. Молекула белка-фермента как механическая система.— В кн.: Колебательные процессы в биологических и химических системах. М.: Наука, 1967, с. 54.
Чайковский Ю. В. Некоторые проблемы дарвинизма и возможности машинного моделирования.— Жури. общ. биол., 1972, т. 33, с. 347.
Чайковский Ю. В. Генетическая интеграция клеточных структур как фактор эволюции.— Жури. общ. биол., 1977, т. 39, с. 823.
Чайковский Ю. В. Выживание мутантного клоиа. Сообщение I. Качественный анализ конкуренции двух клонов. Сообщение II. Мажорирующий анализ судьбы мутанта в полиморфной колонии.— Генетика, 1977, т. 13, с. 1467.
Чернавский Д. С., Чернавская Н. М. Проблема возникновения жизни.— В кн.: Теоретическая и экспериментальная биофизика. Калининград: Изд-во Калининград, ун-та, 1973, вып. 4, с. 36.
Четверикова Е. П. Колебания активности креатинкиназы, выделенной из скелетных мышц.— Биофизика, 1968, т. 13, с. 864.
Черкашин А. Н., Азарашвили А. А. Формакологические исследования памяти.— Журн. ВНД 1972, т. 22, с. 504.
Шапвиль Ф. Энни А.—Л. Биосинтез белка. М., Мир, 1977.
Шапошников Г. X. Направленность эволюции.— Журн. общ. биол., 1977, т. 38, с. 649.
Шапошников Г. X. Динамика клонов, популяций и видов и эволюция.— Журн. общ. биол., 1978, т. 39, с. 15.
Шароян С. Г., Федькина В. Р., Шноль С. Э. О возможной роли макромолекулы белка в ферментативном катализе: Анализ концепций Хиишел- вуда и Касселя.— Биофизика, 1972, т. 17, с. 960.
Шароян С. Г. Развитие представлений о природе ферментативного катализа. М.: ВИНИТИ, деп. № 4215, 1973, с. 72.
Шилов И. А. Регуляция теплообмена у птиц (эколого-физиологический очерк). Докт. дис. М.: МГУ им. Ломоносова, 1964.
Шилов Н. А. О сопряженных реакциях окисления. М., 1905.
Шмальгаузен И. И. Факторы эволюции. М.: Наука, 1968.
Шмальгаузен И. И. Проблемы дарвинизма. М.: Наука, 1969,
Шноль С. Э. О связывании меченых аминокислот белками: Канд. дис. М.: Цент, ин-т усовершенствования врачей, 1956.
Шноль С. Э. О самопроизвольных синхронных переходах молекул акто- миозина в растворе из одного состояния в другое.— Вопр. мед. хим. 1958, т. 4, с. 443.
Шноль С. Э. Синхронные конформационные колебания молекул актина, миозина и актомиозина в растворах.—В кн.: Молекулярная биофизика. М.: Наука, 1965, с. 56.
Шноль С. Э. Конформационные колебания макромолекул.— В кн.: Колебательные процессы в химических и биологических системах. М.: Наука,
с. 22.
Шноль С. Э. Спонтанные обратимые изменения «конформационные колебания» препаратов мышечных белков. Докт. дис. Пущино-иа-Оке: Ин-т биофизики АН СССР, 1970.
Шноль С. Э. Физико-химический аспект процесса биологической эволюции. Возможная детерминированность основных этапов эволюции физическими свойствами эволюционирующей системы.— Журн. общ. биол.
т. 34, с. 331.
Шноль С. Э. Физико-химические механизмы и биологическая специфичность.— В кн.: Современное естествознание и материалистическая диалектика. М.: Наука, 1977. 350 с.
Шноль С. Э., Кондрашова М. Н., Шольц X. Ф. О многофазном характере зависимости АТФ-азной активности актомиозина и миозина от разных воздействии.— Вопр. мед. химии, 1957, т. 3, с. 54.
Шноль С. Э., Христова М. Л., Калиниченко Л. П. Влияние алифатических спиртов на амплитуду конформационных колебаний актомиозина и на скорость поглощения кислорода митохондриями.— В кн.: Свойства и функции макромолекул и макромолекулярных систем. М.: Наука, 1969, с. 89.
Шноль С. Э., Ермакова Е. А., Франк Г. М. Диффузные ограничения и эволюционный смысл образования внутриклеточных структур.— В кн.: Методологические и теоретические проблемы биофизики. М.: Наука, 1979.
Шредингер Э. Что такое жизнь? М.: Атомиздат, 1972.
Шубин В. Н„ Кабакчи С. А. Теория и методы радиационной химии воды. М.: Наука, 1969.
Шулейкин В. В. Очерки по физике моря. 4-е изд. М., Изд-во АН СССР,
1962.
Шулейкин В. В., Лукьянова В. В,, Стась И. И. Сравнительная динамика морских животных.— Докл. АН СССР, 1939, т. 22, с. 424.
Шульман С. С. Проблема происхождения Metazoa.— В кн.: Теоретические вопросы систематики. М.: Наука, 1974, с. 47.
Штерне Карус. Эволюция мира. М.: Т-во «Мир», 1909, т. 1.
Эйген М. Самоорганизация материи и эволюция биологических макромолекул. М.: Мир, 1973,
Эйдус Л. X. О механизме мышечного сокращения.— Биофизика, 1962, т. 7, с. 683.
Энгельгардт В. А. Специфичность биологического обмена веществ.—В кн.: О сущности жизни. М.: Наука, 1964, с. 35.
Энгельгардт В. А. Пути химии в познании явлений жизни. М.: Наука, 1965.
Энгельгардт В. А., Любимов М. Н. К механохимии мышц.— Биохимия, 1942, т. 7, с. 205.
Энгельс Ф. Диалектика природы. М.: Госполитиздат, 1946, с. 158, 245, 246.
Энгельс Ф. Анти-Дюринг. М.: Госполитиздат, 1953, с. 77.
Эсау К. Анатомия растений. М.: Мир, 1969.
Юньев Г. С. Скорость распространения возбуждения в центральной нервной системе. Минск: Мин-во высш. и среди, спец. и проф. образования,
1963.
Яблоков А. В. О разных формах прогрессивного развития в органической природе.— Проблемы эволюции, 1968, т. 1. с. 98.
Ягужинский Л. С., Богуславский Л. И., Волков А. Г., Рахманинова А. Б. Синтез АТФ, сопряженный с функционированием мембранных протонных помп на границе раздела октан/вода.— Докл. АН СССР, 1975, т. 221, с. 1465.
360а. Yaguzhinsky L. S., Boguslavsky L. I., Volkov A. G., Rakhmaninova A. B. Synthesis of ATP coupled with action of membrane protonic pumps at the octane-water interface.—Nature, 1976, v. 259, p. 494.
Allen N. S. Endoplasmic filaments generate the motive force for rotational streaming in Nitella.—J. Cell. Biol., 1974, v. 63, p. 270.
Allen R. D., Taylor D. L. The molecular bases of amoeboid movement.—In: Molecular and cell movement/Ed. S. Inone, R. E. Stephens. N. Y.: Raven Press, 1975. 239 p.
Amos W. B. Contraction and calcium binding in the vorticellid ciliates.— In: Molecules and cel! movement/Ed, S. Inone, R. E. Stephens. N. Y.: Raven Press, 1975, 411 p.
Belehradek J. Temperature and living matter.— Protoplasma Monograph., Berlin, 1935, v. 8.
Berestovski G. N., Frank G. М., Lunesvski V. Z., Raszin V. D. Electroopti- cal phenomena in bimolecular phospholipid membrane.— Biochim. et biop- hys. acta, 1970, v. 219, p. 263.
Berg H. C. How bacteria swim.-—Sci. Amer. Aug., 1975, p. 36.
Berkner L. V., Marshall L. C. On the origin rize of oxygen concentration in the Earth’s atmosphere.—J. Atmos. Sci., 1965, v. 22, p. 225.
Blake J. Hydrodynamic calculations on the movements of Cilia and Fla- gella. II. Opalina.—J. Theor. Biol., 1975, v. 52, p. 67.
Botre C. et al. The generation of spikes by lipo-collagen membranes.—Biochim. et biophys. acta, 1-970, v. 219, p. 283.
Brown H., Escombe F. Static diffusion of gases and liquids in relation to the assimilation of carbon and translocation in plants.—Phil. Trans. Roy. Soc. London, 1900, v. 193B, p. 223.
Carter C. W., Kraut J. A proposed model for interaction of polypeptides with RNA.—Proc. Nat. Acad. Sci. USA, 1974, v. 71, p. 283.
Crow J. E., Kimura M. Evolution in sexual and asexual populations.—Amer. Natur., 1965, v. 99, p. 439.
Crow I. E., Kimura M. Evolution in sexual and asexual populations: a reply.— Amer. Natur., 1969, v. 103, p. 89.
Danon A., Stoekenins W. Photophosphorylation in Halobacterium halobi- um.—Proc. Nat. Acad. Sci. USA, 1974, v. 1, p. 1234.
Davey С. B., Miller R. J. Correlation of temperature dependent water properties and the growth of bacteria.—Nature, 1966, v. 209, p. 638
Deshcherevskii V. I. A kinetic theory of striated muscle contraction—Bior- heology, 1971, v. 7, p. 147.
Dickerson R. The structure of cytochrome and the rates of molecular evolution.— J. Mol. Evol., 1971, v. 1, p. 26.
Delbruck М., Zimmer K. G., Timofeeff-Ressovskii N. W. Uber die Nature der Genmutation und der Genstruktur (цит. no [344]).
Dobzhansky T. Genetics and the origin of species 3rd. N. Y.: Columbia' Univ. Press, 1951.
Dreyer G. et al. Ober Strukturanomalien des Wassers.— Naturwissenschaf- ten, 1969, Bd. 11, S. 558; Stud, biophys., 1971, Bd. 27, S. 2.
Drost-Hansen W. Temperature anomalies and biological temperature optima in the process of evolution.—Naturwissenschaften 1956, Bd. 43, S. 512; Millero F. J., Drost-Hansen W„ Korson L. Relative viscosity and apparent molal volume of aqueous sodium sulfate at various temperatures.—J. Phys. Chem., 1968, v. 72, p. 2251.
Drost-Hansen W. Structure and properties of water at biological interface.— Prepr. Cell interface, 1970.
Ehrenstein G., Lecar H. Electrically gated ionic channels in lipid bilayers.— Quart. Revs Biophys., 1977, v. 10, N 1, p. 1—34.
Eigen М., Schuster P. The hypercycle. A principle of natural selforganization.—Naturwissenschaften, 1977, Bd. 64, H. II; 1978, Bd. 65, H. 1; 1978, Bd. 75, H. 7.
Exobiology/Ed. C. Ponnamperuma. Amsterdam, 1972.
Felsenstein J. The evolutionary advantage of recomb'nation.—Genetics, 1974, V. 78, p. 737.
Fisher R. A. The genetical theory of natural selection. Oxford: Clarendon' Press, 1930.
Fox S. W. Origins of biological information and the genetic code — Mol. and Cell. Biochem., 1974, v. 3, p. 129.
Garay A. S. Origin and role of optical isomery in life.—Nature, 1968, v. 219, p. 338.
Garay A. S. et al. Differences in the annihilation of positrons in optical isomers.—Chem. Phys. Lett., 1973, v. 23, p. 549.
Gause G. F. Problems of evolution.— Trans. Connecticut Acad. Arts and" Sci., 1947, v. 137, p. 17.
Goldman R. D. The distribution of actin in non-muscle cells.— Exp. Cell’ Res., 1975, v. 90, p. 333.
Goldman R. D., Knipe D. Functions of cytoplasmic fibers in non-muscle ceTi motility.— Cold Spring Harbor Symp. Quant. Biol., 1973, v. 37, p. 523.
Gulik-Krzywiski Т., Rivas E„ Luzzati V. Structure et polymorphisme des- lipides: etude par diffraction des rayons X du systeme forme de lipides de* mitochondries de coeur de boeuf.—J. Mol. Biol., 1967, v. 27, p. 321.
Gutfreund H. Enzymes: Physical principles. N. Y.: Wiley, 1972.
Heilbrunn L. V. The colloid chemistry of protoplasma.—Amer. J. Physiol.,. 1924, v. 69, p. 190.
Heilbrunn L. V. An outline of general physiology. Philadelphia, 1952.
Heine R. Hydraulic conductivity in trees.—J. Exp. Bot., 1971, v. 22, p. 503:
Hille B. Ionic channels in nerve membranes.— Progr. Biophys. and Mol. Biol., 1970, v. 21, p. 1.
Hinshelwood C. N. On the theory of unimolecular reactions.— Proc. Roy. Soc. London, 1926, v. 27A, N 113, p. 230.
Horiuti I. —J. Res. Inst. Catal. Hokkaido Univ., 1957, v. 5, p. 1.
Huang B., Mazia D. Microtubules and filaments in ciliate contractility.— In: Molecules and cell movement/Ed. S. Inone, R. E. Stephens. N. Y.: Raven' Press, 1975, p. 389.
Huber B. Untersuchungen fiber die Gesetze der Porenverdunstung.— Z. Bot.., 1930, Bd. 23, S. 839.
Jagendorf A. Т., Uribe E. ATP-formation caused by acid base transition nb spinach chWnplaft.—Proc. Nat. Acad. Sci. USA, 1966, v. 55, p. 170.
Johnson F. H., Eiring H., Polissar M. The kinetic basis of molecular biology. N. Y.: Wiley, 1954.
Kabat D. Potentiation of hemoglobin messenger ribonucleic acid.—J. Biol. Chem., 1975, v. 250, p. 6085.
Kalckar H. Biological phosphorylations. Development of concepts. N. Y.: Englewood Cliffs, 1969.
Katchalsky A. Prebioctic synthesis of biopolymers on inorganic templates.— Naturwissenschaften, 1973, Bd. 60, S. 215.
Katchalsky A. et al. Elementary mechanochemical processes.—In: Contractile polymers. London: Pergamon Press, 1960.
Keynes R. D. Excitable membranes.—Nature, 1972, v. 239, p. 29.
Kimura M. Evolutionary rate at the molecular level.—Nature, 1968, v. 217, p. 624.
Kimura М., Ohta T. Theoretical aspects of polulation genetics. Princeton; New York, 1971.
Kimura M„ Ohta T. Protein polymorphism as a phase of molecular evolution.—Nature, 1971, vol. 228, p. 467.
Kimura М., Ohta T. On the rate of mocular evolution.—T. Mol. Evol., 1971, v. 1, p. 1.
Kno.wles A. F., Racker E. Formation of ATP from H3PO4 and ADP by purified Ca ATP-ase.—J. Biol. Chem., 1949, 1975. v. 250.
Krasnovskii A. A. The models of the evolution of photochemical electron transfer.— In: Chemical evolution and origin of life/Ed. R. Buvet, C. Pon- namperuma. 1971. 279 p.
Ladbrooke B. D. et al. Physical studies of myelin.— Biochim. et biophys. acta, 1968, v. 164, p. 101.
Larsen S. H. et al. Chemomechanical coupling without ATP: the source of energy for motility and chemotaxis in bacteria.—Pros. Nat. Acad. Sci. USA,
v. 71, p. 1239.
Liberman E. A. et al. Mechanism of coupling of oxidative phosphorylation and the membrane potential of mitochondria.—Nature, 1969, v. 222, p. 1076.
■420. Lipmann F. Metabolic generation and utilization of phosphate bond energy.—Adv. Enzym., 1941, v. 1, p. 99.
Lipmann F. et al. Polypeptide synthesis on protein templates. The enzymatic synthesis of gramicidin S and tyrocidine.—Adv. Enzym., 1971, v. 35, p. 1.
Luzzati V., Husson F. The structure of the liquid-crystalline phases of lipid water systems.—J. Cell. Biol., 1962, vol. 12, p. 207; Reiss-Russon F., Lussa- ti V.—Adv. Biol, and Med. Phys. 1967, v. 11, p. 87.
Martynov L. A. et al. Some features of morphogenesis Acetabularia mcdi- terrianea.—Protoplasma, 1974, v. 80, p. 175.
Maynard Smith I. What use is sex?—J. Theor. Biol., 1971, v. 30, p. 319.
424a. Maynard Smith J. The evolution of Sex. Cambr. Univ. Press, 1976 Ldn,
N. Y.
Mitchell H. K., Houlahan М. B. Neurospora, IV. A temperature sensetive riboflavinless mutant.—Amer. J. Bot., 1946, v. 33, p. 31.
Mitchell P. Coupling of phosphorylation to electron and hydrogen transfer by a chemiosmotic type of mechanism.—Nature, 1961, v. 191, p. 144.
Mitchell P. Chemiosmotic coupling in oxidative and photosynthetic phosphorylation.—Glynn Res., 1966.
427a. Mitchell P. Chemiosmotic coupling and energy transduction.—Theor. and Exp. Biophys., 1969, v. 2, p. 159.
Mitchell P., Moyle J. Photon-transport phosphorylation: some experimental tests.—In: Biochemistry of mitochondria. Acad. Press, 1967. 53 p.
Monod I. Le hasard et la necessity. Essai sur la phylosophie naturelle de la biologie moderne/Ed. du Seuil. Paris, 1970.
Morozow V. N., Bucatina A. E., Shnoll S. E. MechanochemicEi! interaction in enzymes. I. Effect of structural deformation of muscle fibres fixed by glutaraldehyde on their ATP-ase activity.— Stud, biophys., 1974, v. 42, p. 99.
Morozow V. N.. Bucatina A. E. Mechanochemical interaction in enzymes. III. The model of enzyme mechanochemical transducer.— Stud, biophys., 1976.
Mueller H. I. Some genetic aspects of sex.—Amer. Natur., 1932, v. 66, p. 118.
Mueller H. I. The relation recombination to mutational advance.—Mutat. Res., 1964, v. 1, p. 2.
Mueller P., Rudin D. Resting and action potentials in experimental bimole- cular lipid membranes.—J. Theor. Biol., 1968, v. 18, p. 222.
Mueller P., Rudin D. Translocators in bimolecular lipid membranes, their role in dissipative and conservative bioenergy transductions.— Curr. Top. Bioenerg., 1969, v. 3. 157 p.
Netter H. Theoretical biochemistry. Physicochemical principles of vital processes. London: Oliver and Boyd, 1969.
Ninneman H., Butter W. L., Epel B. L. Inhibition of respiration in yeast by light.—Biochim. et biophys. acta, 1970. v. 205, p. 499.
Oesper P. Sources of the high energy content in energy rich phosphates.— Arch. Biochem., 1950, v. 27, p. 255.
O’Neil J., Adami L. The oxygen isotope partition function ratio of water and the structure of liquid water.—Phys. Chem. 1969, v. 73, p. 1553.
Oplatka A., Tirosh R. Active streaming in actomyosin solutions.—Biochim. et biophys. acta, 1973, v. 305, p. 684.
Oplatka A. et al. Demonstration of mechanochemica! coupling in systems containing actin, ATP and non aggregating active myosin derivatives.—J. Mechanochem. and Cell Motility, 1974, v. 2, p. 295.
Oppenheimer С. H., Drost-Hansen W. A relationship between multiple temperature optima for biological systems and the properties of water.—J. Bacterid. 1960, v. 80, p. 21.
Overath P., Trouble R. Phase transitions in cells, membranes, and lipids of E. coli. Detection by fluorescent probes, light scattering, and dllatometry.— Biochemistry, 1973, v. 12, p. 2625.
Peschel G., Adlfinger К. H. Temperaturabhangigkeit der Viskositat sehr danner Wasserschichten zwischen Quarzglassoberflachen.—Naturwissenscraf- ten, 1969, Bd. 11, S. 558.
Philipson P. E. On the possible importance of relaxation processes in enzyme catalysis.—J. Mol. Biol., 1968, v. 31, p. 319.
Pirie N. W. The meaninglessness of the therms life and living.—In: Perspectives in biochemistry. N. Y.: McMillan, 1937.
Ponnamperuma C„ Sagan C., Mariner R. Synthesis of ATP under possible- primitive each conditions.—Nature, 1963, v. 199, p. 222.
Ponnamperuma C. Primordial organic chemistry and the origin of life.— Quart. Revs Biophys., 1971, v. 4, p. 77.
Porcellabi G. Lipid metabolism and its regulation in brain tissue.—In: Centra! neurons systems studies of metabolism regulation and function/Ed. E. Qenazzatti, H. Herkew, 1974, p. 12.
Rashevsky N. Contributions to the theory of organismic sets. Why arc there- only two sexes?—Bull. Math. Biophys., 1970, v. 32, p. 293.
Reuter H. Divalent cations as charge carriers in exitable membranes.— Progr. Biophys. and Mol. Biol., 1973, v. 26.
Rutten M. G. The history of atmospheric oxygen.— Space Life Sci., 1970, v. 2, p. 5.
Saxinger C., Ponnamperuma C. Experimental investigation on the origin of the genetic code.—J. Mol. Evol., 1971, v. 1, p. 63.
Sawyer L. I. E. A technique for recording the filtering activity of marine invertebrates.—J. Cons, intern, explor. mer., 1972, v. 34, p. 308.
Shikama K., Nakamura K. J. Standard free energy maps for the hydrolysis of ATP as a function of pH and metal ion concentration. Comparison of metal ions.—Arch. Biochem. and Biophys., 1973, v. 157, p. 457.
Shine /., Dalgarno L. The 3-terminal sequence of E. coli 16S rRMA: complementary to nonsense triplets and ribosome binding sites.—Proc. Nat. Acad. Sci. USA, 1974, v. 71, p. 1342.
Shirom M„ Stein G. Excited state chemistry of the ferrocyanide ion in aqueous solution. Formation of the hydrated electron.—J. Chem. Phys., 1971,. v. 55, p. 3372.
Shnoll S. E., Chetverikova E. P. Synchronous reversible alterations in enzy matic activity (conformational fluctuations) in actomyosin and creatine kinase preparations.— Biochim. et biophys. acta, 1975, v. 403, p. 89.
Skou J. C. The (Na+, K+)-activated enzyme system and its relation to transport of sodium and potassium.—Quart. Revs Biophys., 1975, v. 7, p. 401.
Spiegelman S. An approach to the experimental analysis of precellular evolution.—Quart. Revs Biophys., 1971, v. 4, p. 213.
Stefan I. Versuche fiber die Verdampfung.—Sitzungsber. Bayer. Akad. Wiss. Math.-naturwiss. KU 1873; Bd. 68, S. 385. Cber die Diffusion der Fliissig- keiten.—Ibid., 1878, Bd. 78, S. 957; Ober die Verdampfung aus einem Kreis oder elliptisch begrenzten Becken.—Inbid., 1881, Bd. 82, S. 943.
Takenana Т., Inonue I., Ishima Y., Horie H. Exitability of surface membrane of protoplasmic drop produced from protoplasm in Nitella.—Proc. Jap. Acad.
v. 47, p. 554.
Taniguchi K-, Post R. L. Synthesis of ATP and exchange between H3PO4 and ATP in Na, К ion transport ATP-ase.—J. Biol. Chem., 1975, v. 250, p. 3070.
Taylor G. Analysis of the swimming of microscopic organisms.—Proc. Roy. Soc. London, 1951, v. 17, A, p. 209.
Thermobiology/Ed. H. Rose. Acad. Press, 1967.
Thomas B. D. Origin of the genetic code.— Biochem. and Biophys. Res. Communs, 1970, v. 40, p. 1289.
Timofeeff-Resovsky N. W. The experimental production of mutations.— Biol. Revs Cambridge. Phil. Soc., 1934, v. 9, p. 411.
Tifnofeeff-Resovsky N. W. Eine biophysikalische Analyse des Mutations Vor- ganges.—Nova acta leopold., 1940, v. 9, p. 232.
Ungar G. The problem of molecular coding of neural information. A critical review.—Naturwissenschaften, 1973, Bd. 60, S. 307.
Uribe E. G., Li В. C. Y. Stimulation and inhibition of membrane dependent ATP synthesis in chloroplast by artificially induced K+ gradients.—Bioenergetics, 1973, v. 4, p. 435.
Vennesland B. Steric specifity of hydrogen transfer in pyridine nucleotide dehydrogenase reactions.—J. Cell, and Comp. Physiol., 1956, v. 47, Suppl. 1, p. 201.
Wu T. Y. Hydromechanics of microorganism propulation.—Navig. Res , 197-3, v. 26, p. 12—28.
Yano M. Observation of steady streamings in a solution of Mg ATP and acto-heavy meremyosin from rabbit skeletal muscle.—J. Biochem., 1978, v. 83, p. 1203.
Yarus M. Recognition of nucletodie sequences.—Annu. Rev. Biochem., 1969, v. 38, p. 841.
ОГЛАВЛЕНИЕ
Предисловие ответственного редактора 3
Предисловие автора 5
Глава 1. Физико-химические механизмы и биологическая специфичность 9
Глава 2. Кинетические критерии естественного отбора и физико-химические и биологические факторы
биологической эволюции 19
Глава 3. Естественный отбор матричных ^макромолекул,
способных к конвариантному воспроизведению 44
Глава 4. Ферментативный катализ 65-
Глава 5. Кинетический смысл и возможные способы структурной организации биохимических про- ,
цессов ■ 77
Глава 6. Мембраны, ионная асимметрия 88
Глава 7. Биохимические механизмы превращения энергии 103
Глава 8. Проблемы морфогенеза 142
Глава 9. Перемещения в пространстве. Механо-химиче- ские преобразования энергии. Происхождение
многоклеточности 167
Глава 10. Управление движением. Нервная система ... 191
Глава 11. Теплокровность и теплоустойчивость 210
Глава 12. Мозг. Способность к абстракции. Сознание . . 222 Глава 13. Общий итог. Детерминированность траекторий биологической эволюции — следствие естественного отбора 229
Приложение. Примеры биологической конвергенции, иллюстрирующие детерминированность направлений биологической эволюции условиями задач, решаемых под давлением естественного
отбора 234
Литература 244
Симон Эльевч Шно. *
ФИЗИКО
ХИМИЧЕСКИЕ
ФАКТОРЫ
БИОЛОГИЧЕСКОЙ
ЭВОЛЮЦИИ
Утверждено к печати Институтом биологической физики Академия наук СССР
Редакторы издательства М. В. Медннкова, А. М. \ лалевич Художник Т. Е. Андр'_,*ва Художественный редактор Н. Н. Еласик Технический редактор Р. М. Денисова Корректор Р. А. Тютина
ИБ № 16229
Сдано в набор 14.11.78.
Подписано к печати 13,03.79 Т-02712. Формат 60X 90'/.е- Бумага типографская № I Гарнитура литепатурная Печать высыхая Уел. печ. л. '16,5. Уч.-изд. л. 18,1 Тираж 3950 экз. Тип. Зак. 4376 Цена I р. 80 к.
Издательство «Наука»
(7364 ГСП 7, Москва, В-485, Профсоюзная ул л 94а
2-я типография ручательства «Наука*
121099, Моско^ * О, Шубинский пер., 10
1 Чрезвычайно большое значение имеет открытие А. И. Опариным возможности «добиологического» возникновения основных химических компонентов живых систем. Этим объяснялась детерминированность, неизбежность начала биологической эволюции ^227, 228). (См. также [315]).
1 В относительно недавнее время представления о белках, как о «живых молекулах», наделенных способностью к самообновлению — «обмену веществ», ошибочно развивались [149]. Критика этих работ опублиховаиа [334].
1 Закономерности эволюции автокаталитических систем рассмотрены А. П. Руденко [249].
1 «Факторы биологической эволюции» в контексте этой книги — синоним «критерии естественного отбора». Факторы эволюции — то, что определяет ее направление в данный момент. Употребляемое здесь понимание термина «факторы эволюции» не совпадает с употреблением этого термина в ряде предшествующих работ [см. 289, 290, 316]. К. М. Завадский и Э. И. Колчинский [107] обращают внимание на различный смысл, вкладываемый разными авторами в словосочетание «факторы эволюции», и подчеркивают необходимость точного определения этого термина.
1 Как это ни странно, каталитические свойства нуклеотидов и полинуклеотидов совершенно не изучены. Мы вполне твердо знаем, что биологическими катализаторами служат именно белки, и поэтому даже не начали изучения ферментативных свойств нуклеиновых кислот. Как ясно из приведенных выше соображений, нуклеиновые кислоты должны проявлять каталитические свойства — иначе были бы невозможны первые стадии биологической эволюции.
Наличие специфических каталитических свойств полинуклеотидов следует, возможно, и из того, что большинство современных коферментов близко по химической природе к нуклеотидам или являются нуклеотидами (пиридин- нуклеотиды, флавиннуклеотиды, Ко — А, тиаминпирофосфат, пиридоксаль- фосфат, УТФ, АТФ и т. д ).
1 С современными представлениями о механизмах биосинтеза белка можно ознакомиться по книгам [11а, 176, 276, 297, 325],
1 Функциональные особенности универсальных клеточных органелл — митохои- дрий обеспечивают целесообразные изменения происходящих в них процессов. Состояния митохондрий соответствуют (определяют) физиологическое состояние тканей и органов (см. [146, 146а]. Генетические аспекты возникновения клеточных структур рассмотрены [320].
1 В самом деле, какова консистенция протоплазмы? После замечательных успехов электронно-микроскопических исследований в биологии возникло общее мнение: протоплазма — не жидкость, ие «суп», где свободно плавают орга- неллы, а жестко организованное гелеобразными структурами образование. Однако в большинстве случаев это представление основано иарассмотрении электронных микрофотографий фиксированных препаратов, тогда как ответ на вопрос о консистенции протоплазмы могут дать лишь прижизненные наблюдения. Именно такие наблюдения (например, над формой и движением микрокапли масла, введенной в живую клетку), показали, что протоплазма представляет собой изотропную жидкость. Эти данные, а также другие подтверждения жидкого состояния протоплазмы приведены в книге патриарха физиологии клетки Л. Гейльбруиа «Динамика живой протоплазмы» [64]. (См. также [396, 397].
1 Одчако в нашем случае этой небольшой выгодой можно пренебречь. Любопытно, что еще недавно в спонтанном переходе к более стабильным термодинамическим последовательностям аминокислот — в замене в уже синтезированных молекулах белка одних аминокислотных остатков на другие, даю щис более прочные пептидные связи, видели причину старения.
1 Фотохимия комплексов Fe2+ и Fe3+ (как, впрочем, и простых солей), изучена далеко ие достаточно (см. [287]). Комплексные соли типа красной и жел
той кровяных солей отличаются устойчивостью: они ие гидролизуются в обычных услсвиях, и скорость окислительно-восстановительных реакций с их участием почти на пять порядков выше, чем с участием некомплексных ионов-
железа.
1 Биологическому морфогенезу посвящено большое число высокоценных работ (среди них необходимо назвать статьи Н. К. Кольцова [144] и К- Уод- дингтоиа [295, 296], А. Г. Гурвича [76], Л. В. Белоусова [18], Б. П. Токи- иа [291], Дж. Иберта [113], Соннебориа [274].
1 Надмолекулярные комплексы, рассмотренные выше, образуются в результате взаимодействия циклически изменяющих свою конформацию макромолекул белков. Обычно рассматривают надмолекулярные структуры без учета «конформационных колебаний» [45].
1 С. В. Багоцкий полагает, что инфузории произошли от примитивных многоклеточных организмов — реснитчатых червей. Тогда миофибриллы — мионе- мы инфузорий — в некотором смысле рудиментарные образования, обеспечивающие быстрые изменения формы тела.
1 А. Н. Мосолов обнаружил весьма важное явление: все хромосомы прикреплены к общему цеитромерному кольцу и движутся вслед за движением этого кольца. Их расположение друг относительно друга определено таким образом [216].
1 Такое подтверждение получено в работе (473].
1 См., однако, [260].
1 Недавнее открытие и изучение противоградиентного движения протонов через мембрану Halobacterium halobium [374] за счет энергии света, поглощаемого бактериородопсином, может резко изменить отношение к каротинои- дам. Теперь вполне ясно, что каротиноиды ие просто светозащитные очки, а высокоэффективные преобразователи энергии в биологических системах (см. гл. 7, с. 141).
1 Я благодарен А. П. Сарвазяиу за консультацию по этим вопросам.
1 В современных исследованиях, насколько мне известно, микротрубочки не рассматриваются с такой точки зрения.
1 В ходе эволюции в качестве медиатора, вероятно, выступали различные вещества— от аминокислот, АТФ, метаболитов, до специфических веществ — ацетилхолина, адреналина, серотонина.
1 На это обратил мое внимание Л. П. Каюшии.
1 Беседа с О. С. Виноградовой была очень полезна мне при анализе этого вопроса.
1 Проблеме детерминированности биологической эволюции посвящено большое число исследований философов. Библиографию по этому вопросу следует искать в специальных изданиях [309, 273а].
[1] Цитата приведена с сокращениями.
[2] Термодинамические особенности эволюционирующих систем анализируются в работах [209, 210].
[3] Недавно М. Эйген опубликовал результаты дальнейшей разработки этих проблем [384].
[4] В этой связи существен вопрос: зачем рибосоме РНК? Неужели только для правильного расположения рибосомальиых белков (каждый из которых должен узнавать соответствующее место на РНК [275])? Очень вероятно, что РНК в рибосоме иесет и каталитические функции, ускоряя синтез полипси- тидиой цепи [278, 406, 4561.
[5] Проблемы физики макромолекул, их своеобразие, обусловленное положением иа грани компетенций макрофизики (механики) и микрофизики (квантовой механики), разрабатываются И. М. Лифшицем [179а, б]. Применительно к биологическим проблемам это своеобразие макромолекул анализирует Л. А. Блюменфельд [29].
[6] На медленность ферментативных реакций, независимо от меня, обратил внимание в своем докладе на семинаре Кафедры биофизики физического факультета МГУ Камен Недев.
[7] Д. М. Спитковский [277] обращает особое внимание на роль теплового расширения воды, происходящего при выделении энергии в ферментативной реакции. Этим вызываются конвекционные потоки, способствующие перемешиванию и транспорту вещестл в протоплазме.
[8] Современные данные по этим вопросам изложены, например, в книге О. Хита.. Фотосинтез. Физиологические аспекты. М.: Мир, 1972.
[9] На самом деле, некоторые последовательности нуклеотидов или аминокислот в соответствующих полимерах термодинамически выгоднее, чем другие.
[10] Обсуждение проблемы происхождения нефти (см. [230, 242]).
133
[11] Я благодарен А. И. Опарину, обратившему мое внимание на это обстоятельство.
[12] Я благодарен Г. Б. Кедрову за ценную консультацию по этим вопросам.
[13] Вопросы синхронизации распределенных в среде осцилляторов рассмотрены в книге 10. М. Романовского и др. [245].
[14] Сокращение белкового чехла фагов при иифецироваиии клеток отличается от иных форм биологической подвижности однократностью действия. Механизмы этого сокращения (см. [236]).
[15] Я не рассматриваю здесь возможные пути стаиовлеиия многоклеточности. Этому посвящены труды А. А. Захваткииа [ПО], А. В. Иванова [114, 115], С. С. Шульмана [348], В. В. Хлебовича [312]. Моей задачей является лишь попытка анализа эволюционных причин, т. е. главным образом физико-химических факторов эволюции, критериев естественного отбора, приводящих к появлению многоклеточности.
[16] Ранее и независимо этот метод был разработай и использован для регистрации движения беспозвоночных животных Б. Я- Виленкиным и А. Н. Заики- ным [50а].
[17] Эту возможность допускает и Э. Пондер (см. [239]).
[18] См. статьи М. В. Волькеиштейна в журналах «Наука и жизнь» и «Химия и
жизнь».
[19] См. также статью А. 3. Серикова Г2Я2].
[20] Этот вывод был сделан мною до появления экспериментальных исследований, подтверждающих его верность.
[21] Написано совместно с А. С. Кондрашовым.