



Во второй книге 11 тома «Полного собрания сочинений» И. П. Павлова публикуются труды И. П. Павлова по физиологии и патологии пищеварения, вышедшие в период с 1897 по 1911 г., в том числе и «Лекции о работе главных пищеварительных желез».
Приложенные в конце "Лекций" списки работ автора и его сотрудников, а также библиографические данные к другим статьям уточнены и в большей своей части дополнены.


 Теперь следовало бы заняться вопросом: может ли деятельность нашего желудочка быть верным отражением отделительной работы большого желудка при нормальном пищеварении, когда в большом желудке пища соприкасается со стенками, а наш остается пустым? Однако подробный ответ на него я откладываю до одной из позднейших лекций, когда для решения вопроса мы будем располагать большим фактическим материалом. Здесь же коротко скажу, что, помимо точных выводов из ряда бесспорных фактов, многочисленные случаи прямого сравнения желудочка и желудка в отношении условий работы и качества продукта не оставляют сомнения в том, что желудочек является в своих полных и законных правах, когда мы нем сосредоточиваем изучение нормальной желудочной деятельности. Уже в ближайшие лекции наш желудочек выставит себя с серьезной и поучительной стороны.
Как уже упомянуто выше, в самое недавнее время (после опубликования нашего метода) доктору Фремону удалось изолировать по способу Тири целый желудок собаки, т. е. соединить нижний конец пищевода с duodenum, и в желудок, закрытый с обоих концов, вставить обыкновенную фистульную трубку. Этот методический прием может служить, однако, лишь для некоторых отдельных опытов над желудочным отделением, о чем впоследствии. Как общий метод он представляет два существенных недостатка: 1) при обыкновенном пищеварении у таких собак нельзя рассчитывать на полную нормальность условий отделительного процесса в желудке, так как не имеется ни малейшего рефлекторного раздражения слизистой оболочки желудка пищей; 2) в случае введения веществ прямо в желудок, образуется смесь отделения с этими введенными веществами. Что касается до получения сока из такого желудка для каких-нибудь практических целей, то в этом отношении наш способ комбинации обыкновенной желудочной фистулы с эзофаготомиею, как нам кажется, имеет большие преимущества перед фремоновским уже по одному тому, что наш неизмеримо проще с операционной стороны и при хорошей обстановке не требует напрасных жертв, к тому же наши животные живут многие годы вполне здоровыми и цветущими. То ли с собаками доктора Фримена?
Обыкновенная обстановка собирания сока из нашего желудочка такова. В слепой мешок вставляется стеклянная или, лучше, каучуковая трубка с дырчатыми стенками того конца, который вводится в желудочек. Трубочка или только вкладывается в желудочек, или подвязывается эластическим шнуром около туловища, и собирают сок то в лежачем, то в стоячем положении животного.
Насколько мне кажется сейчас, метод уединенного желудочка должен считаться единственно возможным и вполне правильным в принципе; остаются мелкие подробности вроде разъедания краев раны, потери сока, которые или легко устраняются, или не имеют большого значения и которые, наконец, впоследствии можно было бы исключить совершенно.
В интересах изучения всей отделительной работы пищеварительного канала остается желать всяческого упрощения описанных методов с технической стороны, устранения в них второстепенных недостатков, чтобы иметь возможность сделать на одном и том же животном, без опасности для жизни и вреда здоровью, несколько одновременных фистул.
Из выше сделанного общего обзора пищеварения ясно, до какой степени приобретает важность изучение согласования работ отдельных желез, а оно может быть произведено с безусловной точностью в отношении временинтенсивности ит. д. только в том случае, если мы следим на одном животном сразу за деятельностью всех или многих желез.
Заканчивая методическую часть своих лекций, я нахожу нелишним несколько остановиться на значении хирургических приемов в физиологии. Мне кажется, что в ряду современных методов физиологии должна усилить свою позицию хирургическая методика (противопоставляю ее чисто вивисекционной методике), т. е. производство (искусство, как и творчество) более или менее сложных операций, имеющих целью или удалить органы, или открыть доступ к физиологическим явлениям, происходящим скрыто в глубине тела, уничтожить ту или другую существующую связь между органами или, наоборот, установить новую и т. д., и затем уменье залечить и вернуть, сколько это допускается сущностью операции, общее состояние животного к норме.
Мне представляется такое выдвигание оперативного приема необходимым потому главнейше, что обыкновенное, простое резание животного в остром опыте, как это выясняется теперь с каждым днем все более и более, заключает в себе большой источник ошибок, так как акт грубого нарушения организма сопровождается массою задерживающих влияния на функцию разных органов. Весь организм как осуществление тончайшей и целесообразной связи огромного количества отдельных частей не может остаться индифферентным по своей сущности к разрушающим его агентам и должен в своих интересах одно усилить, другое затормозить, т. е. как бы временно, оставив другие задачи, сосредоточиться на спасении того, что можно. Если это обстоятельство служило и служит большой помехой в аналитической физиологии, то оно кажется непреодолимым препятствием для развития синтетической физиологии, когда понадобится точно определить действительное течение тех или других физиологических явлений в целом и нормальном организме. Вместе с тем оперативная изобретательность как прием физиологического мышления не только не изжита в физиологии, а, наоборот, как указывает действительность, только входит в пору расцвета. Припомним экстирпацию pancreas Минковским, переведение крови из воротной вены в нижнюю полую доктором Экком и, наконец, поразительные операции Гольца с удалением различных отделов центральной нервной системы. Разве не решаются этим многие физиологические вопросы и не возникает в счет их огромная масса новых? Мне скажут, что это уже и есть! Да, но, во-первых, в целом редко, изолированно. Если сравнить, например, количество физических инструментов, вводимых и предлагаемых ежегодно для изучения физиологических явлений, а также число физиолого-химических методов и их вариаций, с числом новых физиологических операций, после которых животное должно оставаться жить, то бедность последних неоспорима при сравнении с богатством первых. Во-вторых, обращает на себя внимание, что большое число таких операции было сделано в свое время хирургами, а не физиологами; физиологи как бы не считают это своим существенным делом или не располагают для достижения цели нужными средствами. Наконец самое сильное свидетельство за несоответствующее интересу дела положение хирургической методики в физиологии представляет собою отсутствие в плане современной физиологической лаборатории серьезного, т. е. отвечающего вполне цели, хирургического отделения, как есть химическое, физическое, микроскопическое и вивисекционное.
Если нужны частые и сложные операции над животными, после которых они должны жить, то нельзя, конечно, экономно употребляя время и труд, делать эти операции в общих комнатах, не применяя всех тех приемов и обстановок, которых требует современная хирургия. Нет сомнения, что некоторые операции в обыкновенных лабораториях, хотя бы и при применении правил антисептики и асептики, не удадутся, не пойдут, потому что при животных наблюсти полную чистоту как во время операции, так и в первый период после нее почти невозможно без обширного и специально устроенного для этого отделения. Укажу для примера на хорошо мне известную историю экковской операции соединительного свища нижней полой и воротной вен. В старых лабораториях, несмотря на всю энергию и остроумие ее изобретателя, не удалось получить надолго в живых животных, подвергшихся этой операции. Та же неудача преследовала и профессора Стольникова, который при содействии доктора Экка, не жалея собак и труда, повторял ту же операцию. И лишь в операционном отделении физиологической лаборатории Института экспериментальной медицины, тогда (в 1891 г.) только что учрежденного, следовательно чистого в хирургическом отношении здания, впервые получился значительный процент успеха. Но эта счастливая пора операции продолжалась только год. Так как первоначальное физиологическое отделение Института было тесно, то, несмотря на принимаемые меры, загрязнение лаборатории пошло так быстро, что та же экковская операция в тех же руках, может быть даже теперь еще более привычных, стала бесплодным занятием. И это, несмотря на упорство операторов (около года), продолжалось до тех пор, пока в Институте не построена была новая физиологическая лаборатория, где операционному отделу отведено обширное помещение.
Позволяю себе привлечь ваше внимание к этому первому, сколько я знаю, примеру специального операционного отделения физиологической лаборатории. Может быть, этот пример даст некоторые полезные указания и товарищам физиологам при устройстве новых физиологических институтов. Отделение занимает половину верхнего этажа, четверть всего здания лаборатории. Оно состоит, с одной стороны, из ряда комнат для операций собственно: в первой из них животное получает ванну и обсушивается на особенных платформах; в следующей комнате (подготовительная операционная) животное наркотизуется, бреется в определенных местах и моется антисептическими жидкостями; третья комната служит для стерилизации инструментов, белья, мытья рук операторов и переодевания их, и четвертая - операционная с усиленным освещением. В эту комнату наркотизованное и приготовленное животное переносится, без стола, участвующими в операции лицами. Служители обыкновенно дальше второй комнаты не пускаются в операционном отделении.
Капитальной стеной от этих комнат отделается ряд комнаток для содержания оперированвотных в первые 10 дней после операции. Каждая из комнаток имеет большое окно с форточкой, площадь ее около квадратной сажени, высота с лишком пять аршин, нагревание производится трубами с гретым воздухом и освещение - электричеством. Перед собачьими комнатами идет во всю длину коридор, причем каждая комнатка отделяется от коридора массивной, плотно пригнанной дверью. Полы во всем отделении из цемента, со стоком в каждой комнате. Комнатки для собак внизу кругом всех стен имеют свинцовую трубку с отверстиями, из которых во всякое время из коридора, не заходя в комнатки, может быть обмываем весь пол. Все отделение сверху донизу окрашено белой масляной краской. Длинный ряд операционных комнат представляется лучшею гарантиею против проникания грязи в последнюю, главную операционную комнату. Как ни одолжена много физиология интеллекту собак вообще, надеяться на помощь умных животных при достижении хирургических целей было бы напрасно. И лишь устраивая такой длинный ряд преград для грязи, в простом и хирургическом смысле слова, можно было рассчитывать сохранить операционное отделение на долгое время на высоте его задачи. Два года пользования этим отделением не повели к его загрязнению, как следует из показаний реактива хирургической чистоты -- экковской операции. Проводя в моей памяти результаты операций за 20 лет в различных помещениях, постоянно работая над однообразным, т. е. всегда здоровым, материалом и часто повторяя одни и те же операции, я резче, может быть, чем хирурги, поражался торжеством чистоты, сохранявшей огромное число жизней животных и чрезвычайно экономизировавшей время и труд экспериментатора.
Надеюсь, вы простите мне столь длинную экскурсию в сторону хирургического метода в физиологии. Я убежден, что только развитие оперативного остроумия и искусства в области пищеварительного канала раскроет перед нами всю поразительную красоту химической работы этого органа, отдельные черты которой мы можем подсмотреть уже и с современными методическими средствами. Прошу припомнить эти мои фразы в конце лекций - и вы признаете, я уверен в том, заключенную в них правду.
Теперь следовало бы заняться вопросом: может ли деятельность нашего желудочка быть верным отражением отделительной работы большого желудка при нормальном пищеварении, когда в большом желудке пища соприкасается со стенками, а наш остается пустым? Однако подробный ответ на него я откладываю до одной из позднейших лекций, когда для решения вопроса мы будем располагать большим фактическим материалом. Здесь же коротко скажу, что, помимо точных выводов из ряда бесспорных фактов, многочисленные случаи прямого сравнения желудочка и желудка в отношении условий работы и качества продукта не оставляют сомнения в том, что желудочек является в своих полных и законных правах, когда мы нем сосредоточиваем изучение нормальной желудочной деятельности. Уже в ближайшие лекции наш желудочек выставит себя с серьезной и поучительной стороны.
Как уже упомянуто выше, в самое недавнее время (после опубликования нашего метода) доктору Фремону удалось изолировать по способу Тири целый желудок собаки, т. е. соединить нижний конец пищевода с duodenum, и в желудок, закрытый с обоих концов, вставить обыкновенную фистульную трубку. Этот методический прием может служить, однако, лишь для некоторых отдельных опытов над желудочным отделением, о чем впоследствии. Как общий метод он представляет два существенных недостатка: 1) при обыкновенном пищеварении у таких собак нельзя рассчитывать на полную нормальность условий отделительного процесса в желудке, так как не имеется ни малейшего рефлекторного раздражения слизистой оболочки желудка пищей; 2) в случае введения веществ прямо в желудок, образуется смесь отделения с этими введенными веществами. Что касается до получения сока из такого желудка для каких-нибудь практических целей, то в этом отношении наш способ комбинации обыкновенной желудочной фистулы с эзофаготомиею, как нам кажется, имеет большие преимущества перед фремоновским уже по одному тому, что наш неизмеримо проще с операционной стороны и при хорошей обстановке не требует напрасных жертв, к тому же наши животные живут многие годы вполне здоровыми и цветущими. То ли с собаками доктора Фримена?
Обыкновенная обстановка собирания сока из нашего желудочка такова. В слепой мешок вставляется стеклянная или, лучше, каучуковая трубка с дырчатыми стенками того конца, который вводится в желудочек. Трубочка или только вкладывается в желудочек, или подвязывается эластическим шнуром около туловища, и собирают сок то в лежачем, то в стоячем положении животного.
Насколько мне кажется сейчас, метод уединенного желудочка должен считаться единственно возможным и вполне правильным в принципе; остаются мелкие подробности вроде разъедания краев раны, потери сока, которые или легко устраняются, или не имеют большого значения и которые, наконец, впоследствии можно было бы исключить совершенно.
В интересах изучения всей отделительной работы пищеварительного канала остается желать всяческого упрощения описанных методов с технической стороны, устранения в них второстепенных недостатков, чтобы иметь возможность сделать на одном и том же животном, без опасности для жизни и вреда здоровью, несколько одновременных фистул.
Из выше сделанного общего обзора пищеварения ясно, до какой степени приобретает важность изучение согласования работ отдельных желез, а оно может быть произведено с безусловной точностью в отношении временинтенсивности ит. д. только в том случае, если мы следим на одном животном сразу за деятельностью всех или многих желез.
Заканчивая методическую часть своих лекций, я нахожу нелишним несколько остановиться на значении хирургических приемов в физиологии. Мне кажется, что в ряду современных методов физиологии должна усилить свою позицию хирургическая методика (противопоставляю ее чисто вивисекционной методике), т. е. производство (искусство, как и творчество) более или менее сложных операций, имеющих целью или удалить органы, или открыть доступ к физиологическим явлениям, происходящим скрыто в глубине тела, уничтожить ту или другую существующую связь между органами или, наоборот, установить новую и т. д., и затем уменье залечить и вернуть, сколько это допускается сущностью операции, общее состояние животного к норме.
Мне представляется такое выдвигание оперативного приема необходимым потому главнейше, что обыкновенное, простое резание животного в остром опыте, как это выясняется теперь с каждым днем все более и более, заключает в себе большой источник ошибок, так как акт грубого нарушения организма сопровождается массою задерживающих влияния на функцию разных органов. Весь организм как осуществление тончайшей и целесообразной связи огромного количества отдельных частей не может остаться индифферентным по своей сущности к разрушающим его агентам и должен в своих интересах одно усилить, другое затормозить, т. е. как бы временно, оставив другие задачи, сосредоточиться на спасении того, что можно. Если это обстоятельство служило и служит большой помехой в аналитической физиологии, то оно кажется непреодолимым препятствием для развития синтетической физиологии, когда понадобится точно определить действительное течение тех или других физиологических явлений в целом и нормальном организме. Вместе с тем оперативная изобретательность как прием физиологического мышления не только не изжита в физиологии, а, наоборот, как указывает действительность, только входит в пору расцвета. Припомним экстирпацию pancreas Минковским, переведение крови из воротной вены в нижнюю полую доктором Экком и, наконец, поразительные операции Гольца с удалением различных отделов центральной нервной системы. Разве не решаются этим многие физиологические вопросы и не возникает в счет их огромная масса новых? Мне скажут, что это уже и есть! Да, но, во-первых, в целом редко, изолированно. Если сравнить, например, количество физических инструментов, вводимых и предлагаемых ежегодно для изучения физиологических явлений, а также число физиолого-химических методов и их вариаций, с числом новых физиологических операций, после которых животное должно оставаться жить, то бедность последних неоспорима при сравнении с богатством первых. Во-вторых, обращает на себя внимание, что большое число таких операции было сделано в свое время хирургами, а не физиологами; физиологи как бы не считают это своим существенным делом или не располагают для достижения цели нужными средствами. Наконец самое сильное свидетельство за несоответствующее интересу дела положение хирургической методики в физиологии представляет собою отсутствие в плане современной физиологической лаборатории серьезного, т. е. отвечающего вполне цели, хирургического отделения, как есть химическое, физическое, микроскопическое и вивисекционное.
Если нужны частые и сложные операции над животными, после которых они должны жить, то нельзя, конечно, экономно употребляя время и труд, делать эти операции в общих комнатах, не применяя всех тех приемов и обстановок, которых требует современная хирургия. Нет сомнения, что некоторые операции в обыкновенных лабораториях, хотя бы и при применении правил антисептики и асептики, не удадутся, не пойдут, потому что при животных наблюсти полную чистоту как во время операции, так и в первый период после нее почти невозможно без обширного и специально устроенного для этого отделения. Укажу для примера на хорошо мне известную историю экковской операции соединительного свища нижней полой и воротной вен. В старых лабораториях, несмотря на всю энергию и остроумие ее изобретателя, не удалось получить надолго в живых животных, подвергшихся этой операции. Та же неудача преследовала и профессора Стольникова, который при содействии доктора Экка, не жалея собак и труда, повторял ту же операцию. И лишь в операционном отделении физиологической лаборатории Института экспериментальной медицины, тогда (в 1891 г.) только что учрежденного, следовательно чистого в хирургическом отношении здания, впервые получился значительный процент успеха. Но эта счастливая пора операции продолжалась только год. Так как первоначальное физиологическое отделение Института было тесно, то, несмотря на принимаемые меры, загрязнение лаборатории пошло так быстро, что та же экковская операция в тех же руках, может быть даже теперь еще более привычных, стала бесплодным занятием. И это, несмотря на упорство операторов (около года), продолжалось до тех пор, пока в Институте не построена была новая физиологическая лаборатория, где операционному отделу отведено обширное помещение.
Позволяю себе привлечь ваше внимание к этому первому, сколько я знаю, примеру специального операционного отделения физиологической лаборатории. Может быть, этот пример даст некоторые полезные указания и товарищам физиологам при устройстве новых физиологических институтов. Отделение занимает половину верхнего этажа, четверть всего здания лаборатории. Оно состоит, с одной стороны, из ряда комнат для операций собственно: в первой из них животное получает ванну и обсушивается на особенных платформах; в следующей комнате (подготовительная операционная) животное наркотизуется, бреется в определенных местах и моется антисептическими жидкостями; третья комната служит для стерилизации инструментов, белья, мытья рук операторов и переодевания их, и четвертая - операционная с усиленным освещением. В эту комнату наркотизованное и приготовленное животное переносится, без стола, участвующими в операции лицами. Служители обыкновенно дальше второй комнаты не пускаются в операционном отделении.
Капитальной стеной от этих комнат отделается ряд комнаток для содержания оперированвотных в первые 10 дней после операции. Каждая из комнаток имеет большое окно с форточкой, площадь ее около квадратной сажени, высота с лишком пять аршин, нагревание производится трубами с гретым воздухом и освещение - электричеством. Перед собачьими комнатами идет во всю длину коридор, причем каждая комнатка отделяется от коридора массивной, плотно пригнанной дверью. Полы во всем отделении из цемента, со стоком в каждой комнате. Комнатки для собак внизу кругом всех стен имеют свинцовую трубку с отверстиями, из которых во всякое время из коридора, не заходя в комнатки, может быть обмываем весь пол. Все отделение сверху донизу окрашено белой масляной краской. Длинный ряд операционных комнат представляется лучшею гарантиею против проникания грязи в последнюю, главную операционную комнату. Как ни одолжена много физиология интеллекту собак вообще, надеяться на помощь умных животных при достижении хирургических целей было бы напрасно. И лишь устраивая такой длинный ряд преград для грязи, в простом и хирургическом смысле слова, можно было рассчитывать сохранить операционное отделение на долгое время на высоте его задачи. Два года пользования этим отделением не повели к его загрязнению, как следует из показаний реактива хирургической чистоты -- экковской операции. Проводя в моей памяти результаты операций за 20 лет в различных помещениях, постоянно работая над однообразным, т. е. всегда здоровым, материалом и часто повторяя одни и те же операции, я резче, может быть, чем хирурги, поражался торжеством чистоты, сохранявшей огромное число жизней животных и чрезвычайно экономизировавшей время и труд экспериментатора.
Надеюсь, вы простите мне столь длинную экскурсию в сторону хирургического метода в физиологии. Я убежден, что только развитие оперативного остроумия и искусства в области пищеварительного канала раскроет перед нами всю поразительную красоту химической работы этого органа, отдельные черты которой мы можем подсмотреть уже и с современными методическими средствами. Прошу припомнить эти мои фразы в конце лекций - и вы признаете, я уверен в том, заключенную в них правду.
 Задаемся дальнейшим вопросом. Если железа в течение пищеварительного периода резко и определенно изменяет напряжение своей работы, что касается массы доставляемого ею сока, то не свойственно ли ей видоизменять также и самые качества выделяемого продукта? Рассуждая теоретически, можно ожидать, что в различных фазах обработки одной и той же пищи понадобится сок различного состава. Вся масса пищи может сразу или постепенно измениться под влиянием первых порций сока в известных химических и физических отношениях и, таким образом, при продолжении обработки нуждаться в других свойствах сока: в большем или в меньшем содержании воды, в большей или меньшей кислотности, щелочности и в том или другом содержании различных ферментов. Все эти отдельные факторы соков, конечно, не без значения, а на основании опытов с пищеварением в химическом стакане мы далеко не отдали себе отчета в смысле всех их. Правда, наука уже давно умеет положительный ответ на вопрос об изменении качеств сока во время отделительного периода. Но, мне кажется, наблюденный факт не был оценен во всем его значении; иначе он должен был бы сделаться неистощимой темой настойчивых исследований: для чего это и как это происходит? Я опять приведу из нашего запаса наблюдений примеры этих в высшей степени любопытных колебаний свойств соков во время одного и того же отделительного периода. Конечно, в этом случае наибольший интерес привязывается по привычке к колебаниям содержания ферментов, хотя, в сущности говоря, одинаково заслуживают точного констатирования и удовлетворяющего объяснения колебания и других свойств соков.
Существующий материал, в особенности относительно желудочного сока, нельзя считать достаточным. Опыты с обыкновенной желудочной фистулой допускали только очень условные заключения, так как относились не к чистому соку, а к смеси сока с пищей. Наблюдения же, поставленные Гейденгайном на уединенном дне желудка, не могли претендовать на приложение их к нормальному пищеварению, так как деятельность уединенного желудочка, вследствие перерезки его секреторных нервов, очевидно, сильно отклонена от нормальной. Только исследование Гейденгайном хода панкреатического отделения на собаках, оперированных по способу, указанному в первой лекции, должно быть признано вполне точным научным материалом. К сожалению, исследование это приводится лишь в очень сокращенном виде в известном энциклопедическом руководстве по физиологии, изданном Германом, не появившись виде журнальной статьи.
Прежде чем обратиться к нашим фактам, я принужден, хоть на короткое время, занять ваше внимание теми особенными приемами анализа соков, которыми мы пользовались в наших работах. Переваривающая белки сила сока определялась по способу Метта, выработанному и постоянно употребляемому в нашей лаборатории. Способ состоит в том, что в стеклянную палочку диаметра в 1-2 мм втягивается жидкий яичный белок и свертывается там при определенной температуре (95°), 50 затем стеклянная палочка ломается на куски, которые опускаются в 1--2 куб. см испытуемой жидкости. Жидкость с палочками ставят в термостат при температуре в 3710 часов, без какого бы то ни было дальнейшего наблюдения. На концах стеклянной трубки происходит растворение белка. По истечении срока, измеряя под микроскопом с малым увеличением, при помощи миллиметровой линейки, длину стеклянного кусочка и длину оставшегося в нем непереваренным белкового цилиндрика, мы легко определяем величины переваривания в миллиметрах и его долях. Способ не оставляет ничего лучшего желать в отношении удобства, объективности и точности. Специальные опыты (д-ра Самойлова) показали, что переваривание белкового цилиндрика идет строго пропорционально времени, по крайней мере в течение десяти часов, и при таких силах переваривающих жидкостей, которые исследовали мы и которые должны быть признаны вообще наибольшими. Этим исключалось существенное подозрение, что растворение белка на различных глубинах нашего цилиндрика могло происходить с различной быстротой вследствие большего или меньшего застаивания продуктов переваривания в полости трубки. Таким образом количество миллиметров белка, переваренного за один и тот же период времени порциями разных соков, представляет точную сравнительную меру переваривающей силы этих порций. В исследованиях Борисова над этим методом, произведенных в лаборатории профессора Тарханова, отчетливо выступило правило соотношения между количеством миллиметров переваренного белка и количеством пепсина в сравниваемых растворах, именно: количества пепсина относились как квадраты скоростей переваривания, т. е. как квадрать чисел миллиметров белкового столбика, растворенного заин и тот же срок времени.
Поясним правило примером на числах. Если одна жидкость переварила 2 мм, а другая за то же время, то относительное количество пепсина в этих жидкостях выражается не числами 2 и 3, а их квадратами, т. е. 4 и 9. Разница очевидна: прямо по миллиметрам выходило, что во втором случае фермента в полтора раза больше, а на основании правила, т. е. по квадратам этих чисел, - в два раза с четвертью. Конечно, это правило было выведено на основании сравнения искусственно и точно составленных растворов пепсина. Результат, полученный Борисовым самостоятельно, был уже до него установлен Шюцем путем определения в растворе помощью поляризационного прибора образующихся при переваривании пептонов. Такое совпадение при различных методах составляет вескую гарантию точности правила. Не могу при этом не выразить сожаления, что ментовский метод, предложенный еще в 1889 г., до сих пор все еще не приобрел себе такого распространения, которого он по справедливости заслуживает. Он мог бы легко стать универсальным методом для измерения силы белковых ферментов, так что все исследования над этими ферментами были бы легко сравниваемы между собою. Нельзя отрицать, что это очень и очень желательно. Тогда все наблюдения над соками различных животных и людей, представляя одну и ту же скалу, повели бы к важным заключениям колебанияы ферментов по индивидуумам, родам и видам животных. Нужно прибавить еще, что в меттовском методе диаметр трубки в широких пределах не имеет значения, а куриные яйца в нашем отношении оказываются чрезвычайностоянного состава. Шюц-борисовское правило оказалось применимо в полной силе и к трипсину.
Определение других ферментов менее совершенно и в наших исследованиях постоянно подвергалось и подвергается видоизменениям. Крахмальный фермент панкреатического сока долгое время определялся в нашей лаборатории путем известного титрования фелинговой жидкостью сахара, получающегося из крахмала при известных и всегда одинаковых условиях ферментного действия. В таком случае число миллиграммов образованного сахара являлось мерою количества крахмального фермента. Метод этот, давая хорошие и точные цифры, требовал, однако, очень значительного времени и был чувствительно тяжел в таких опытах, где надобилось многопределений. Поэтому естественно было искать более скорого метода. В последнее время. лаборатория в лице докторов Глинского и Вальтера старалась достигнуть этой цели, настаивая вместе с тем на однообразии метода определения обоих ферментов. Тоненькие стеклянные трубочки наполнялись вареным крахмалом, обыкновенно чем-нибудь подкрашенным, и затем в термостате подвергались определенный срок времени (обыкновенно полчаса) действию испытуемой жидкости. Крахмал с концов растворялся, и граница растворения, благодаря окраске, была вполне резка. Как выше при белке, сосчитывались миллиметры растворенного крахмального столбика. Многократные пробы с искусственным раствором фермента (панкреатический сок, разбавленный 2-3 раза, и т. д.) установили точные соотношения числа миллиметров с содержанием фермента. Здесь также оказалось вполне приложимо правило Шюца и Борисова, т. е. количества фермента относились, как квадраты миллиметров. Поэтому в приводимых нами ниже опытах будут встречаться обе указанные меры крахмального фермента: миллиграммы образованного сахара и миллиметры растворенного крахмального цилиндрика.
К сожалению, все усилия свести и определение жирового фермента к методу цилиндрика в стеклянной трубочке до сих пор оставались бесплодными. В конце концов пришлось пользоваться титрованием баритом смеси жира панкреатическим соком, стоявшей определенное время при определенной температуре и периодически взбалтываемой. Следовательно, число куб. сантиметров баритового титра, истрачиваемого на образующуюся кислоту, является мерою жирового фермента. Конечно, неудачи не должны останавливать дальнейших попыток здесь добиться успеха, как с другими ферментами. настоящем своем виде метод нуждается в постоянном внимании со стороны экспериментатора и потому затруднителен при массе определений по часам или в еще более короткие промежутки времени. К тому же метод этот представляет некоторые колебания в результатах. Однако правило Шюца и Борисова оказалось приложимо и здесь. Конечно, в наших дальнейших опытах, строго говоря, имеется только ферментное действие, и речь о количествах и суммах ферментов должна пониматься условно.
Несколько слов об определении щелочности панкреатического сока. С этою целью сожигали на слабом огне твердый остаток от определенной порции сока и, растворив золу в дистиллированной воде, подвергали ее титрованию. Результат рассчитывался на соду и выражался в процентах взятой первоначально массы сока.
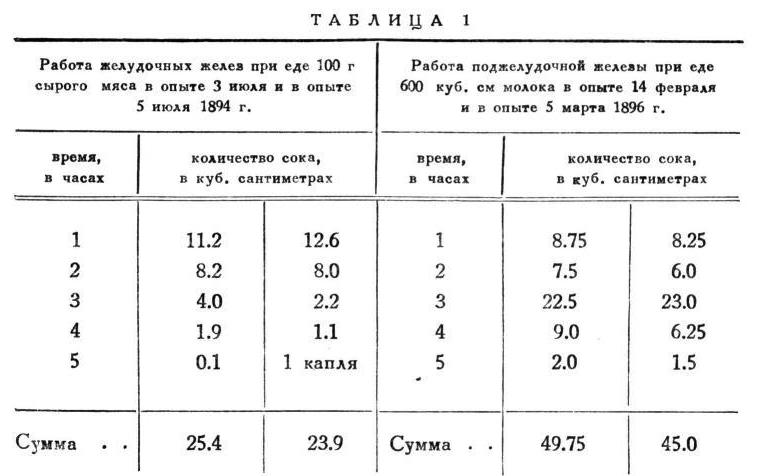
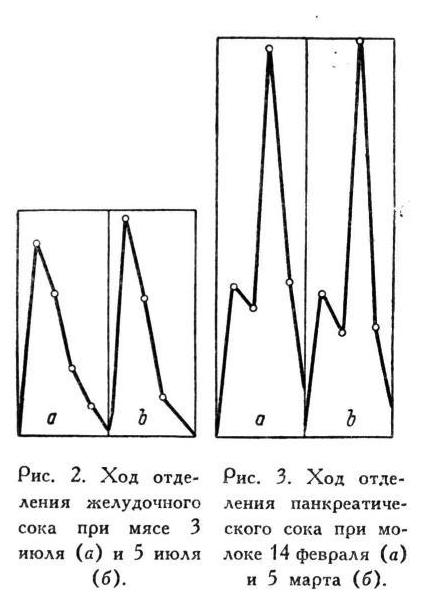
Опыты, перед которыми я прервал изложение ради необходимых предварительных методических разъяснений, приведу опять в виде двух пар: одну - для желудочных желез и другую - для поджелудочной, как доказательство и при колебаниях состава сока в течение отделительного периода той же закономерности, с которою мы уже познакомились выше при колебаниях количества сока по часам (табл. 2 и 3).
То же представляю в виде кривых (рис. 4).
Задаемся дальнейшим вопросом. Если железа в течение пищеварительного периода резко и определенно изменяет напряжение своей работы, что касается массы доставляемого ею сока, то не свойственно ли ей видоизменять также и самые качества выделяемого продукта? Рассуждая теоретически, можно ожидать, что в различных фазах обработки одной и той же пищи понадобится сок различного состава. Вся масса пищи может сразу или постепенно измениться под влиянием первых порций сока в известных химических и физических отношениях и, таким образом, при продолжении обработки нуждаться в других свойствах сока: в большем или в меньшем содержании воды, в большей или меньшей кислотности, щелочности и в том или другом содержании различных ферментов. Все эти отдельные факторы соков, конечно, не без значения, а на основании опытов с пищеварением в химическом стакане мы далеко не отдали себе отчета в смысле всех их. Правда, наука уже давно умеет положительный ответ на вопрос об изменении качеств сока во время отделительного периода. Но, мне кажется, наблюденный факт не был оценен во всем его значении; иначе он должен был бы сделаться неистощимой темой настойчивых исследований: для чего это и как это происходит? Я опять приведу из нашего запаса наблюдений примеры этих в высшей степени любопытных колебаний свойств соков во время одного и того же отделительного периода. Конечно, в этом случае наибольший интерес привязывается по привычке к колебаниям содержания ферментов, хотя, в сущности говоря, одинаково заслуживают точного констатирования и удовлетворяющего объяснения колебания и других свойств соков.
Существующий материал, в особенности относительно желудочного сока, нельзя считать достаточным. Опыты с обыкновенной желудочной фистулой допускали только очень условные заключения, так как относились не к чистому соку, а к смеси сока с пищей. Наблюдения же, поставленные Гейденгайном на уединенном дне желудка, не могли претендовать на приложение их к нормальному пищеварению, так как деятельность уединенного желудочка, вследствие перерезки его секреторных нервов, очевидно, сильно отклонена от нормальной. Только исследование Гейденгайном хода панкреатического отделения на собаках, оперированных по способу, указанному в первой лекции, должно быть признано вполне точным научным материалом. К сожалению, исследование это приводится лишь в очень сокращенном виде в известном энциклопедическом руководстве по физиологии, изданном Германом, не появившись виде журнальной статьи.
Прежде чем обратиться к нашим фактам, я принужден, хоть на короткое время, занять ваше внимание теми особенными приемами анализа соков, которыми мы пользовались в наших работах. Переваривающая белки сила сока определялась по способу Метта, выработанному и постоянно употребляемому в нашей лаборатории. Способ состоит в том, что в стеклянную палочку диаметра в 1-2 мм втягивается жидкий яичный белок и свертывается там при определенной температуре (95°), 50 затем стеклянная палочка ломается на куски, которые опускаются в 1--2 куб. см испытуемой жидкости. Жидкость с палочками ставят в термостат при температуре в 3710 часов, без какого бы то ни было дальнейшего наблюдения. На концах стеклянной трубки происходит растворение белка. По истечении срока, измеряя под микроскопом с малым увеличением, при помощи миллиметровой линейки, длину стеклянного кусочка и длину оставшегося в нем непереваренным белкового цилиндрика, мы легко определяем величины переваривания в миллиметрах и его долях. Способ не оставляет ничего лучшего желать в отношении удобства, объективности и точности. Специальные опыты (д-ра Самойлова) показали, что переваривание белкового цилиндрика идет строго пропорционально времени, по крайней мере в течение десяти часов, и при таких силах переваривающих жидкостей, которые исследовали мы и которые должны быть признаны вообще наибольшими. Этим исключалось существенное подозрение, что растворение белка на различных глубинах нашего цилиндрика могло происходить с различной быстротой вследствие большего или меньшего застаивания продуктов переваривания в полости трубки. Таким образом количество миллиметров белка, переваренного за один и тот же период времени порциями разных соков, представляет точную сравнительную меру переваривающей силы этих порций. В исследованиях Борисова над этим методом, произведенных в лаборатории профессора Тарханова, отчетливо выступило правило соотношения между количеством миллиметров переваренного белка и количеством пепсина в сравниваемых растворах, именно: количества пепсина относились как квадраты скоростей переваривания, т. е. как квадрать чисел миллиметров белкового столбика, растворенного заин и тот же срок времени.
Поясним правило примером на числах. Если одна жидкость переварила 2 мм, а другая за то же время, то относительное количество пепсина в этих жидкостях выражается не числами 2 и 3, а их квадратами, т. е. 4 и 9. Разница очевидна: прямо по миллиметрам выходило, что во втором случае фермента в полтора раза больше, а на основании правила, т. е. по квадратам этих чисел, - в два раза с четвертью. Конечно, это правило было выведено на основании сравнения искусственно и точно составленных растворов пепсина. Результат, полученный Борисовым самостоятельно, был уже до него установлен Шюцем путем определения в растворе помощью поляризационного прибора образующихся при переваривании пептонов. Такое совпадение при различных методах составляет вескую гарантию точности правила. Не могу при этом не выразить сожаления, что ментовский метод, предложенный еще в 1889 г., до сих пор все еще не приобрел себе такого распространения, которого он по справедливости заслуживает. Он мог бы легко стать универсальным методом для измерения силы белковых ферментов, так что все исследования над этими ферментами были бы легко сравниваемы между собою. Нельзя отрицать, что это очень и очень желательно. Тогда все наблюдения над соками различных животных и людей, представляя одну и ту же скалу, повели бы к важным заключениям колебанияы ферментов по индивидуумам, родам и видам животных. Нужно прибавить еще, что в меттовском методе диаметр трубки в широких пределах не имеет значения, а куриные яйца в нашем отношении оказываются чрезвычайностоянного состава. Шюц-борисовское правило оказалось применимо в полной силе и к трипсину.
Определение других ферментов менее совершенно и в наших исследованиях постоянно подвергалось и подвергается видоизменениям. Крахмальный фермент панкреатического сока долгое время определялся в нашей лаборатории путем известного титрования фелинговой жидкостью сахара, получающегося из крахмала при известных и всегда одинаковых условиях ферментного действия. В таком случае число миллиграммов образованного сахара являлось мерою количества крахмального фермента. Метод этот, давая хорошие и точные цифры, требовал, однако, очень значительного времени и был чувствительно тяжел в таких опытах, где надобилось многопределений. Поэтому естественно было искать более скорого метода. В последнее время. лаборатория в лице докторов Глинского и Вальтера старалась достигнуть этой цели, настаивая вместе с тем на однообразии метода определения обоих ферментов. Тоненькие стеклянные трубочки наполнялись вареным крахмалом, обыкновенно чем-нибудь подкрашенным, и затем в термостате подвергались определенный срок времени (обыкновенно полчаса) действию испытуемой жидкости. Крахмал с концов растворялся, и граница растворения, благодаря окраске, была вполне резка. Как выше при белке, сосчитывались миллиметры растворенного крахмального столбика. Многократные пробы с искусственным раствором фермента (панкреатический сок, разбавленный 2-3 раза, и т. д.) установили точные соотношения числа миллиметров с содержанием фермента. Здесь также оказалось вполне приложимо правило Шюца и Борисова, т. е. количества фермента относились, как квадраты миллиметров. Поэтому в приводимых нами ниже опытах будут встречаться обе указанные меры крахмального фермента: миллиграммы образованного сахара и миллиметры растворенного крахмального цилиндрика.
К сожалению, все усилия свести и определение жирового фермента к методу цилиндрика в стеклянной трубочке до сих пор оставались бесплодными. В конце концов пришлось пользоваться титрованием баритом смеси жира панкреатическим соком, стоявшей определенное время при определенной температуре и периодически взбалтываемой. Следовательно, число куб. сантиметров баритового титра, истрачиваемого на образующуюся кислоту, является мерою жирового фермента. Конечно, неудачи не должны останавливать дальнейших попыток здесь добиться успеха, как с другими ферментами. настоящем своем виде метод нуждается в постоянном внимании со стороны экспериментатора и потому затруднителен при массе определений по часам или в еще более короткие промежутки времени. К тому же метод этот представляет некоторые колебания в результатах. Однако правило Шюца и Борисова оказалось приложимо и здесь. Конечно, в наших дальнейших опытах, строго говоря, имеется только ферментное действие, и речь о количествах и суммах ферментов должна пониматься условно.
Несколько слов об определении щелочности панкреатического сока. С этою целью сожигали на слабом огне твердый остаток от определенной порции сока и, растворив золу в дистиллированной воде, подвергали ее титрованию. Результат рассчитывался на соду и выражался в процентах взятой первоначально массы сока.
Опыты, перед которыми я прервал изложение ради необходимых предварительных методических разъяснений, приведу опять в виде двух пар: одну - для желудочных желез и другую - для поджелудочной, как доказательство и при колебаниях состава сока в течение отделительного периода той же закономерности, с которою мы уже познакомились выше при колебаниях количества сока по часам (табл. 2 и 3).
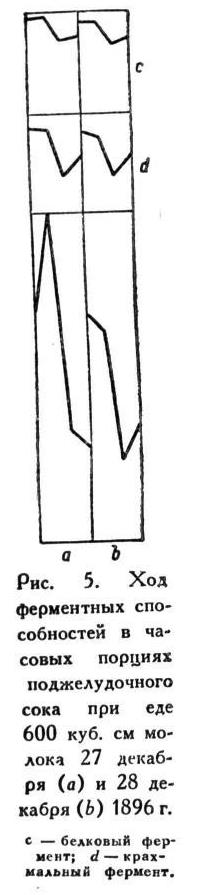
То же представляю в виде кривых (рис. 4).

 То же представляю в виде кривых (рис. 5).
Как видите, опять поражающая точность в работе: то, что раз требуется от железы, воспроизводится постоянно, что называется, в обрез. Итак, перед нами в полной отчетливости важный для работы желез факт, что железа может готовить сок различного состава, с большим или меньшим содержанием ферментов, с различным содержанием различных ферментов, если их, как в панкреатическом соке, - несколько, измеряя также и другие свойства сока, помимо ферментных. Анализ всех имеющихся у нас в этом отношении цифр, сопоставление их с цифрами хода часовых количеств сока, исключает допущение, что сок только меняет свою концентрацию в зависимости от скорости отделения. Имеются самые разнообразные отношения между водой сока и его ферментами: высокая переваривающая сила встречается как при обильном, так и при скудном выделении сока, а факт неоднообразного колебания различных ферментов в одном и том же соке решительно доказывает способность желез с сложную химическую деятельностью, как панкреатическая, вырабатывать в известные периоды отделительного периода то те, то другие фабрикаты. Что сказано относетельно ферментов, то может быть отнесено и к солям сока.
То же представляю в виде кривых (рис. 5).
Как видите, опять поражающая точность в работе: то, что раз требуется от железы, воспроизводится постоянно, что называется, в обрез. Итак, перед нами в полной отчетливости важный для работы желез факт, что железа может готовить сок различного состава, с большим или меньшим содержанием ферментов, с различным содержанием различных ферментов, если их, как в панкреатическом соке, - несколько, измеряя также и другие свойства сока, помимо ферментных. Анализ всех имеющихся у нас в этом отношении цифр, сопоставление их с цифрами хода часовых количеств сока, исключает допущение, что сок только меняет свою концентрацию в зависимости от скорости отделения. Имеются самые разнообразные отношения между водой сока и его ферментами: высокая переваривающая сила встречается как при обильном, так и при скудном выделении сока, а факт неоднообразного колебания различных ферментов в одном и том же соке решительно доказывает способность желез с сложную химическую деятельностью, как панкреатическая, вырабатывать в известные периоды отделительного периода то те, то другие фабрикаты. Что сказано относетельно ферментов, то может быть отнесено и к солям сока.
 Тем интереснее рядом с этим является факт постоянной, как с правом можно думать, кислотности желудочного сока. Хотя клинические наблюдения отделительной желудочной деятельности человека чуть не ежедневно твердят колебании кислотности, хотя и в наших наблюдениях, где дело идет об абсолютно чистом соке, эти колебания вполне заметны, тем не менее внимательный пересмотр фактов ведет к почти несомненному заключению, что желудочный сок приготовляется железами всегда с одной и той же кислотностью. Ведь и при нашей методике сок не получается прямо из желез, а, выйдя из них, течет по стенкам желудка, которые покрыты щелочной слизью, и, следовательно, неизбежно подвергается нейтрализованию - уменьшению своей кислотности. Что этому обстоятельству следует приписать видимые колебания кислотности желудочного сока, явствует из многих наблюдений. Почти постоянное явление, что кислотность сока и скорость его отделения крайне тесно связаны между собой: чем сильнее отделение, тем выше кислотность, и наоборот. Эта связь легко понимается с точки зрения нашего объяснения. Чем в большем количестве отделяется сок, чем быстрее он проходит по делудочным стенкам, тем, естественно, он меньше нейтрализуется по всей своей массе, тем его кислотность более приближается к подлинной. Доктор Кетчер с целью проверить это объяснение применил несколько форм опытов. Так как при пустом желудке стенки его обыкновенно покрыты значительным слоем слизи, то понятно, что первые порции начинающего вытекать сока, например под влиянием мнимого кормления, обыкновенно бывают самой низкой кислотности. Чем больше и обильнее вытекает сок, тем кислотность его все повышается. При загихании отделения, очевидно благодаря сильному нейтрализованию слизи, уже не получается той высокой кислотности, которая в начале опыта наблюдалась при той же скорости. Повторяя несколько раз такие обильные, так сказать, обмывания стенок желудка соком, можно иногда совсем разорвать связь между быстротой и кислотностью, т. е. сок остается одинаково высококислым при значительной, как и малой, быстроте отделения. С другой стороны, доктор Кетчер, прм же мнимом кормлении, каждые 5 минут попеременно, то собирал сок при открытой все время фистуле, то выливал его из желудка в конце каждых 5 минут. Оказалось, чточти все разом вылитые порции, т. e. остававшиеся в желудке 5 минут, имели отчетливо более низкую кислотность. Понятно, если в чистом соке указанным образом производятся колебания кислотности, то тем скорее они окажутся в желудке при вступлении в него слюны вместе с пищей. Если все это действительно так, то, следовательно, та или другая надобности в кислоте при пищеварении должна покрываться главным образом за счет массы желудочного сока, а не степени его кислотности. Однако возможно, что нейтрализование слизью сока также представляет нарочитый и приспособленный к известной цели процесс. Даже при нормальном желудке совершенно чистый сок может терять до 25% своей кислотности благодаря нейтрализованию одной слизью. Почем знать, может быть, природа нашла наиболее выгодным в интересах всего организма или процесса обработки пищи изменять кислотность именно таким образом. Ведь факт измененной кислотности остается фактом, как бы он ни произошел.
Вернемся к главной нити нашего изложения. Вы видели на резких примерах, что желудочная и поджелудочная железы в течение периода обработки одной и той же пищи дают сок не однообразного состава, а меняющийся в различных отношениях. В высшей степени любопытная и важная задача - совершенно уяснить: в какой связи находятся эти колебания с ходом пищеварения, с пользой его? Решение этой задачи в целом - дело будущего. Но целесообразность, по крайней мере некоторых пунктов, бросается в глаза и сейчас.
Возьмем первую порцию желудочного сока; она отличается весьма высокой переваривающей силой. Понятие, что это как нельзя более кстати в начале обработки, при наличности всей массы пищи и наибольшей грубости ее внешнего вида. Значит, наисильнейший реактив выливается тогда, когда в нем наибольшая надобность. Объяснение целесообразности изменений в составе панкреатического сока, очевидно, представит большие трудности, так как теперь дело касается позднейшей инстанции завода, где обрабатывается уже измененный и сортированный желудком пищевой материал. К тому же в кишках возникает необходимость создавать благоприятные химические условия для действия поджелудочного сока, изменяя в пищевой массе обстановку желудочного пищеварения, наверное неблагоприятную для поджелудочного. Мы знаем, что пепсин разрушает трипсин, а высокая кислотность вредит проявлению действия трипсина. К затронутым вопросам мы подойдем ближе впоследствии, анализируя подробно механизм возбуждения желез.
Познакомившись со столькими фактами, свидетельствующими о способности желез сообразоваться с каждым моментом пищевой обработки, мы имеем сильное основание ждать, что при разных сортах еды указанная способность должна выступить в полной своей яркости и красоте. Раз пища состоит из различных составных частей, а в пищеварительный канал изливаются различные реактивы, то естественно предполагать, что на известные сорта пищи будут изливаться по преимуществу то те, то другие сока и, ввиду их изменяемости, то с теми, то с другими свойствами. Как же стоит дело на самом деле? Конечно, о такой задаче не могло быть и речи при старых способах исследования, и, обратно, разработка этих вопросов является лучшей рекомендацией, лучшей заслугой новых методов. Теперь действительно можно было убедиться в том, что представлялось вероятным a priori: каждому роду пищи отвечает своя деятельность желез, свои свойства реактивов.
Начнем с желудка. Исследования на собаках с изолированным желудком показали (д-р Хижин), что смешанной пище, молоку, хлебу, мясу и т. д. отвечает всякий раз совершенно особенная работа желудочных желез в отношении качества всего сока, его количества, отделения и продолжительности всего отделительного процесса. Переберем эти элементы по порядку.
Самой высокой переваривающей силой обладает сок, вытекающий на хлеб; будем называть его для краткости хлебным, как и остальные соответственно. Для него в среднем у доктора Хижина значится 6.64 мм. Мясной пище отвечает сок 3.99 мм. Наконец молочная еда дает сок переваривающей силы 3.26 мм. Воспользуемся правилом Шюца и Борисова для точного сравнения состава всех этих соков. Квадрат хлебного сока 44, квадрат мясного - 16 и квадрат молочного -- 11; следовательно, хлебный сок содержит в себе в 4 раза более фермента, чем молочный, он вчетверо концентрированнее молочного.
Хорошей иллюстрацией к сказанному могут служить следующие опыты (из работы д-ра Хижина).
Тем интереснее рядом с этим является факт постоянной, как с правом можно думать, кислотности желудочного сока. Хотя клинические наблюдения отделительной желудочной деятельности человека чуть не ежедневно твердят колебании кислотности, хотя и в наших наблюдениях, где дело идет об абсолютно чистом соке, эти колебания вполне заметны, тем не менее внимательный пересмотр фактов ведет к почти несомненному заключению, что желудочный сок приготовляется железами всегда с одной и той же кислотностью. Ведь и при нашей методике сок не получается прямо из желез, а, выйдя из них, течет по стенкам желудка, которые покрыты щелочной слизью, и, следовательно, неизбежно подвергается нейтрализованию - уменьшению своей кислотности. Что этому обстоятельству следует приписать видимые колебания кислотности желудочного сока, явствует из многих наблюдений. Почти постоянное явление, что кислотность сока и скорость его отделения крайне тесно связаны между собой: чем сильнее отделение, тем выше кислотность, и наоборот. Эта связь легко понимается с точки зрения нашего объяснения. Чем в большем количестве отделяется сок, чем быстрее он проходит по делудочным стенкам, тем, естественно, он меньше нейтрализуется по всей своей массе, тем его кислотность более приближается к подлинной. Доктор Кетчер с целью проверить это объяснение применил несколько форм опытов. Так как при пустом желудке стенки его обыкновенно покрыты значительным слоем слизи, то понятно, что первые порции начинающего вытекать сока, например под влиянием мнимого кормления, обыкновенно бывают самой низкой кислотности. Чем больше и обильнее вытекает сок, тем кислотность его все повышается. При загихании отделения, очевидно благодаря сильному нейтрализованию слизи, уже не получается той высокой кислотности, которая в начале опыта наблюдалась при той же скорости. Повторяя несколько раз такие обильные, так сказать, обмывания стенок желудка соком, можно иногда совсем разорвать связь между быстротой и кислотностью, т. е. сок остается одинаково высококислым при значительной, как и малой, быстроте отделения. С другой стороны, доктор Кетчер, прм же мнимом кормлении, каждые 5 минут попеременно, то собирал сок при открытой все время фистуле, то выливал его из желудка в конце каждых 5 минут. Оказалось, чточти все разом вылитые порции, т. e. остававшиеся в желудке 5 минут, имели отчетливо более низкую кислотность. Понятно, если в чистом соке указанным образом производятся колебания кислотности, то тем скорее они окажутся в желудке при вступлении в него слюны вместе с пищей. Если все это действительно так, то, следовательно, та или другая надобности в кислоте при пищеварении должна покрываться главным образом за счет массы желудочного сока, а не степени его кислотности. Однако возможно, что нейтрализование слизью сока также представляет нарочитый и приспособленный к известной цели процесс. Даже при нормальном желудке совершенно чистый сок может терять до 25% своей кислотности благодаря нейтрализованию одной слизью. Почем знать, может быть, природа нашла наиболее выгодным в интересах всего организма или процесса обработки пищи изменять кислотность именно таким образом. Ведь факт измененной кислотности остается фактом, как бы он ни произошел.
Вернемся к главной нити нашего изложения. Вы видели на резких примерах, что желудочная и поджелудочная железы в течение периода обработки одной и той же пищи дают сок не однообразного состава, а меняющийся в различных отношениях. В высшей степени любопытная и важная задача - совершенно уяснить: в какой связи находятся эти колебания с ходом пищеварения, с пользой его? Решение этой задачи в целом - дело будущего. Но целесообразность, по крайней мере некоторых пунктов, бросается в глаза и сейчас.
Возьмем первую порцию желудочного сока; она отличается весьма высокой переваривающей силой. Понятие, что это как нельзя более кстати в начале обработки, при наличности всей массы пищи и наибольшей грубости ее внешнего вида. Значит, наисильнейший реактив выливается тогда, когда в нем наибольшая надобность. Объяснение целесообразности изменений в составе панкреатического сока, очевидно, представит большие трудности, так как теперь дело касается позднейшей инстанции завода, где обрабатывается уже измененный и сортированный желудком пищевой материал. К тому же в кишках возникает необходимость создавать благоприятные химические условия для действия поджелудочного сока, изменяя в пищевой массе обстановку желудочного пищеварения, наверное неблагоприятную для поджелудочного. Мы знаем, что пепсин разрушает трипсин, а высокая кислотность вредит проявлению действия трипсина. К затронутым вопросам мы подойдем ближе впоследствии, анализируя подробно механизм возбуждения желез.
Познакомившись со столькими фактами, свидетельствующими о способности желез сообразоваться с каждым моментом пищевой обработки, мы имеем сильное основание ждать, что при разных сортах еды указанная способность должна выступить в полной своей яркости и красоте. Раз пища состоит из различных составных частей, а в пищеварительный канал изливаются различные реактивы, то естественно предполагать, что на известные сорта пищи будут изливаться по преимуществу то те, то другие сока и, ввиду их изменяемости, то с теми, то с другими свойствами. Как же стоит дело на самом деле? Конечно, о такой задаче не могло быть и речи при старых способах исследования, и, обратно, разработка этих вопросов является лучшей рекомендацией, лучшей заслугой новых методов. Теперь действительно можно было убедиться в том, что представлялось вероятным a priori: каждому роду пищи отвечает своя деятельность желез, свои свойства реактивов.
Начнем с желудка. Исследования на собаках с изолированным желудком показали (д-р Хижин), что смешанной пище, молоку, хлебу, мясу и т. д. отвечает всякий раз совершенно особенная работа желудочных желез в отношении качества всего сока, его количества, отделения и продолжительности всего отделительного процесса. Переберем эти элементы по порядку.
Самой высокой переваривающей силой обладает сок, вытекающий на хлеб; будем называть его для краткости хлебным, как и остальные соответственно. Для него в среднем у доктора Хижина значится 6.64 мм. Мясной пище отвечает сок 3.99 мм. Наконец молочная еда дает сок переваривающей силы 3.26 мм. Воспользуемся правилом Шюца и Борисова для точного сравнения состава всех этих соков. Квадрат хлебного сока 44, квадрат мясного - 16 и квадрат молочного -- 11; следовательно, хлебный сок содержит в себе в 4 раза более фермента, чем молочный, он вчетверо концентрированнее молочного.
Хорошей иллюстрацией к сказанному могут служить следующие опыты (из работы д-ра Хижина).
 Влияние разного рода пищи на переваривающую силу сока совершенно очевидно. Однако, чтобы исключить мысль, что на результат мог иметь влияние порядок введения пищи, приводим другой опыт.
Влияние разного рода пищи на переваривающую силу сока совершенно очевидно. Однако, чтобы исключить мысль, что на результат мог иметь влияние порядок введения пищи, приводим другой опыт.
 Как переваривающая сила, колеблется кислотность [11] по родам пищи, являясь самой высокой при мясе (0.56%) и самой низкой при хлебе (0.46%). Так же различаются количества сока и сроки продолжительности отделения случае того или другого сорта еды, возьмем ли мы равные весовые количества этих сорТОВ -- равные ли по твердому остатку или, что еще поучительнее, равные по азоту, так как мы имели перед собой действие желудочного сока только на белки. Соответственно этому оказываются различными средние часовые количества сока, получаемые делением всей массы сока на число часов отделения, т. е. среднее часовое напряжение желез. На равные весовые количества всего больше выливается сока на мясо и всего меньше на молоко, на эквивалентные по азоту - всего больше на хлеб и всего меньше на мясо; часовое напряжение желез почти одинаково у мяса и молока и резко меньше у хлеба, т. e. по продолжительности обработки выделяется из других сортов еды хлеб, сильно затягивая отделение.
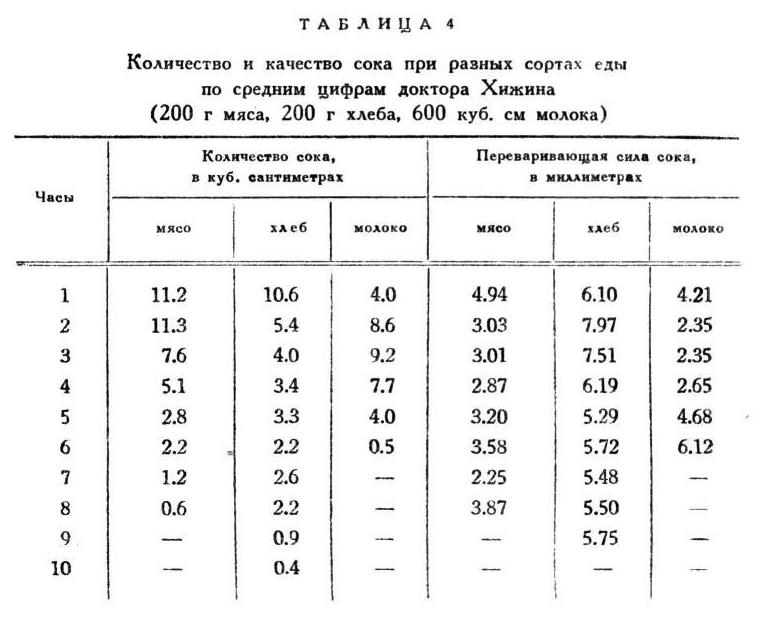
Характерность работы желез при каждом особом сорте еды не исчерпывается приведенными разницами, а дает себя знать выразительными особенностями в ходе отделения, как и в часовых колебаниях качеств сока. На этот раз я представляю только по одному примеру для каждого сорта с просьбою верить, что и здесь точность повторения не меньше, чем в ранее приведенных случаях (табл. 4).
То же представляю в виде кривых (рис. 6 и 7).
Перед нами в высшей степени резкие и любопытные факты: при каждой еде как количество, так и качество сока от часа к часу изменяются совершенно своеобразно. При мясе maximum отделения приходится то на первый, то на второй час, причем эти часы вообще очень мало разнятся друг от друга в отношении количества; у хлеба - всегда и резкий maximum падает на первый час, при молоке - на второй и даже на третий.
У мясного сока высшая концентрация принадлежит первой часовой порции, у хлебного - порции второго или третьего часа, а у молочного - самой последней часовой порции. Так же характерно положение minimum ов, как и вообще весь ход.
Как переваривающая сила, колеблется кислотность [11] по родам пищи, являясь самой высокой при мясе (0.56%) и самой низкой при хлебе (0.46%). Так же различаются количества сока и сроки продолжительности отделения случае того или другого сорта еды, возьмем ли мы равные весовые количества этих сорТОВ -- равные ли по твердому остатку или, что еще поучительнее, равные по азоту, так как мы имели перед собой действие желудочного сока только на белки. Соответственно этому оказываются различными средние часовые количества сока, получаемые делением всей массы сока на число часов отделения, т. е. среднее часовое напряжение желез. На равные весовые количества всего больше выливается сока на мясо и всего меньше на молоко, на эквивалентные по азоту - всего больше на хлеб и всего меньше на мясо; часовое напряжение желез почти одинаково у мяса и молока и резко меньше у хлеба, т. e. по продолжительности обработки выделяется из других сортов еды хлеб, сильно затягивая отделение.
Характерность работы желез при каждом особом сорте еды не исчерпывается приведенными разницами, а дает себя знать выразительными особенностями в ходе отделения, как и в часовых колебаниях качеств сока. На этот раз я представляю только по одному примеру для каждого сорта с просьбою верить, что и здесь точность повторения не меньше, чем в ранее приведенных случаях (табл. 4).
То же представляю в виде кривых (рис. 6 и 7).
Перед нами в высшей степени резкие и любопытные факты: при каждой еде как количество, так и качество сока от часа к часу изменяются совершенно своеобразно. При мясе maximum отделения приходится то на первый, то на второй час, причем эти часы вообще очень мало разнятся друг от друга в отношении количества; у хлеба - всегда и резкий maximum падает на первый час, при молоке - на второй и даже на третий.
У мясного сока высшая концентрация принадлежит первой часовой порции, у хлебного - порции второго или третьего часа, а у молочного - самой последней часовой порции. Так же характерно положение minimum ов, как и вообще весь ход.
 Приведенные факты, как мне кажется, чрезвычайно подкрепляют наше раннее заключение о серьезном значении колебаний отделительной работы в течение отделительного периода: если каждому роду пищи отвечает свой собственный ход отделения. то, значит, он имеет определенное значение и известную важность.
Мы познакомились теперь с очень многими колебаниями железистой работы в различных случаях. Их законность свидетельствует об их важности. Является интересным и необходимым понять смысл каждого колебания. Такое понимание способствовало бы значительному объединению многочисленных фактов, теперь стоящих более или менее особняком и, может быть, неприятно загромождающих ваше внимание и память. Я, однако, при передаче их отнюдь не имел в виду запоминание всех этих сложных отношений, для чего, конечно, требуются многократное повторение и изучение предмета. Мне нужно было только обосновать в вас убеждение, что работа желез крайне эластична, вместе с тем характерна, точна и вполне целесообразна. К сожалению, что касается последнего пункта - это почти непочатый угол для исследования; убеждение в целесообразности колебаний работы желез в настоящее время может основываться главным образом на общих соображениях и лишь частью - на отдельных более или менее ясных и бесспорных случаях ее. Определим количество фермента, истрачиваемого желудком на эквивалентные по N количества разных сортов еды. На хлеб идет 1600 единиц фермента, на мясо - 430 и на молоко - 340. Я получаю эти цифры следующим образом. Для мяса и молока у доктора Хижина есть готовые числа: 100 г мяса эквивалентны по N 600 куб. см молока и 250 г хлеба. Опыта с таким количеством хлеба у доктора Хижина нет, но на основании правила о пропорциональности между количеством пищи и соком легко рассчитать нужное число. Таким образом получается следующие числа:
Приведенные факты, как мне кажется, чрезвычайно подкрепляют наше раннее заключение о серьезном значении колебаний отделительной работы в течение отделительного периода: если каждому роду пищи отвечает свой собственный ход отделения. то, значит, он имеет определенное значение и известную важность.
Мы познакомились теперь с очень многими колебаниями железистой работы в различных случаях. Их законность свидетельствует об их важности. Является интересным и необходимым понять смысл каждого колебания. Такое понимание способствовало бы значительному объединению многочисленных фактов, теперь стоящих более или менее особняком и, может быть, неприятно загромождающих ваше внимание и память. Я, однако, при передаче их отнюдь не имел в виду запоминание всех этих сложных отношений, для чего, конечно, требуются многократное повторение и изучение предмета. Мне нужно было только обосновать в вас убеждение, что работа желез крайне эластична, вместе с тем характерна, точна и вполне целесообразна. К сожалению, что касается последнего пункта - это почти непочатый угол для исследования; убеждение в целесообразности колебаний работы желез в настоящее время может основываться главным образом на общих соображениях и лишь частью - на отдельных более или менее ясных и бесспорных случаях ее. Определим количество фермента, истрачиваемого желудком на эквивалентные по N количества разных сортов еды. На хлеб идет 1600 единиц фермента, на мясо - 430 и на молоко - 340. Я получаю эти цифры следующим образом. Для мяса и молока у доктора Хижина есть готовые числа: 100 г мяса эквивалентны по N 600 куб. см молока и 250 г хлеба. Опыта с таким количеством хлеба у доктора Хижина нет, но на основании правила о пропорциональности между количеством пищи и соком легко рассчитать нужное число. Таким образом получается следующие числа:


 Беря квадраты миллиметров, я имею соответственно 38 для хлеба, 16 для мяса и 10 для молока.
Эти числа дают возможность сравнивать содержание фермента в одинаковом объеме сока. Для того чтобы рассчитать все количество фермента при всех сортах еды, нужно принять во внимание разные количества сока; ради этого приведенные квадраты относятся к одному куб. сантиметру как к единице и множатся соответственно на число куб. сантиметров каждого сока; тогда оказываются вышеприведенные цифры: 1600, 430 и 340, а это значит, что на то же количество хлебного белка издерживается желудком пепсина почти в пять раз больше, чем на белок молока, а на мясной - на 25% больше, чем на молочный. Этот ряд. ферментных затрат на разные белки вполне совпадает с результатами физиолого-химического исследования перевариваемости всех этих белков. При сравнении работы желудочных желез при разных сортах еды нельзя не заметить целесообразности и в другом отношении. Надобность в большом количестве фермента для растительного белка покрывается не столько количеством сока, сколько чрезвычайно увеличенной концентрацией хлебного сока сравнительно с другими соками. Следовательно, можно думать, что надобилось только большое количество фермента и являлось излишним, даже вредным большое количество кислоты. Что действительно в желудке избегался избыток кислоты, на это указывает и другая особенность хлебноготделения. В целом только незначительно большая масса хлебного сока, сравнительно с молочным, распределяется, однако, на гораздо большее время, так что средняя часовая величина хлебного отделеная, как сказано выше, в полтора раза меньше, чем при молоке и мясе. Таким образом при переваривании хлеба в желудке во весь отделительный период содержится относительно небольшое количество соляной кислоты. Эти факты опять хорошо совпадают с физислого-химическими наблюдениями, что избыток кислоты мешает перевариванию крахмала, который в хлебе в таком большом количестве сопровождает белок. И из клинических наблюдений мы знаем, что при hyperaciditas, когда мясо переваривается очень хорошо, масса хлебного крахмала проходит пищеварительный канал неусвоенной.
Может быть, в интересах той же крахмальной обработки существует явление, о котором уже упоминалось ранее несколько раз, но которое оставалось до сих пор без всякого толкования, это длинный период, по крайней мере в 5 минут, всегда наблюдающийся между кормлением животного и началом истечения сока, делается ли наблюдение на целом желудке, как при мнимом кормлении, или на нашем маленьком уединением желудочке. Этот, так сказать, латентный период, не бывая меньше 41/2-5 минут, в другую сторону представляется довольно колеблющимся, чаще всего до 10 минут. Что значит он? Отнести его в какие-нибудь внешние условия, например вроде наполнения желез до края, увлажнения всей поверхности желудка до образования потоков по направлению к фистульному отверстию, нет достаточных оснований, потому что он строго сохраняется при несомненно наполненных железах и при смоченной соком стенке желудка. Представить себе почему-либо неспособность желудочных желез скорее реагировать на раздражение, чем срок времени в 5 минут, было бы странностью. Остается одно- признать в этом какую-то особенную цель. Может быть, эти 5-10 минут рассчитаны на беспрепятственное развитие действия слюнного крахмального фермента. Конечно, такое объяснение не может претендовать на большую убедительность, раз дело идет о факте, еще не подпавшем систематическому научному анализу.
Тем охотнее перехожу я к работе поджелудочной железы, что здесь факт целесообразности ее, по самой сущности наблюдаемых явлений, стоит вне всяких споров и сомнений. Вот таблица (5) опытов с данными количества и содержания ферментов, при тех же сортах еды (из работы д-ра Вальтера).
Беря квадраты миллиметров, я имею соответственно 38 для хлеба, 16 для мяса и 10 для молока.
Эти числа дают возможность сравнивать содержание фермента в одинаковом объеме сока. Для того чтобы рассчитать все количество фермента при всех сортах еды, нужно принять во внимание разные количества сока; ради этого приведенные квадраты относятся к одному куб. сантиметру как к единице и множатся соответственно на число куб. сантиметров каждого сока; тогда оказываются вышеприведенные цифры: 1600, 430 и 340, а это значит, что на то же количество хлебного белка издерживается желудком пепсина почти в пять раз больше, чем на белок молока, а на мясной - на 25% больше, чем на молочный. Этот ряд. ферментных затрат на разные белки вполне совпадает с результатами физиолого-химического исследования перевариваемости всех этих белков. При сравнении работы желудочных желез при разных сортах еды нельзя не заметить целесообразности и в другом отношении. Надобность в большом количестве фермента для растительного белка покрывается не столько количеством сока, сколько чрезвычайно увеличенной концентрацией хлебного сока сравнительно с другими соками. Следовательно, можно думать, что надобилось только большое количество фермента и являлось излишним, даже вредным большое количество кислоты. Что действительно в желудке избегался избыток кислоты, на это указывает и другая особенность хлебноготделения. В целом только незначительно большая масса хлебного сока, сравнительно с молочным, распределяется, однако, на гораздо большее время, так что средняя часовая величина хлебного отделеная, как сказано выше, в полтора раза меньше, чем при молоке и мясе. Таким образом при переваривании хлеба в желудке во весь отделительный период содержится относительно небольшое количество соляной кислоты. Эти факты опять хорошо совпадают с физислого-химическими наблюдениями, что избыток кислоты мешает перевариванию крахмала, который в хлебе в таком большом количестве сопровождает белок. И из клинических наблюдений мы знаем, что при hyperaciditas, когда мясо переваривается очень хорошо, масса хлебного крахмала проходит пищеварительный канал неусвоенной.
Может быть, в интересах той же крахмальной обработки существует явление, о котором уже упоминалось ранее несколько раз, но которое оставалось до сих пор без всякого толкования, это длинный период, по крайней мере в 5 минут, всегда наблюдающийся между кормлением животного и началом истечения сока, делается ли наблюдение на целом желудке, как при мнимом кормлении, или на нашем маленьком уединением желудочке. Этот, так сказать, латентный период, не бывая меньше 41/2-5 минут, в другую сторону представляется довольно колеблющимся, чаще всего до 10 минут. Что значит он? Отнести его в какие-нибудь внешние условия, например вроде наполнения желез до края, увлажнения всей поверхности желудка до образования потоков по направлению к фистульному отверстию, нет достаточных оснований, потому что он строго сохраняется при несомненно наполненных железах и при смоченной соком стенке желудка. Представить себе почему-либо неспособность желудочных желез скорее реагировать на раздражение, чем срок времени в 5 минут, было бы странностью. Остается одно- признать в этом какую-то особенную цель. Может быть, эти 5-10 минут рассчитаны на беспрепятственное развитие действия слюнного крахмального фермента. Конечно, такое объяснение не может претендовать на большую убедительность, раз дело идет о факте, еще не подпавшем систематическому научному анализу.
Тем охотнее перехожу я к работе поджелудочной железы, что здесь факт целесообразности ее, по самой сущности наблюдаемых явлений, стоит вне всяких споров и сомнений. Вот таблица (5) опытов с данными количества и содержания ферментов, при тех же сортах еды (из работы д-ра Вальтера).
 Под концентрациею сока разумеется квадрат числа миллиметров растворенных цилиндриков или куб. сантиметров титрованной щелочи, под абсолютным числом ферментных единиц произведение квадрата на число куб. сантиметров выделенного сока. Сравнению подлежат опять эквивалентные количества по азоту. Мы видим, что у каждого сорта еды свое количество сока, резко разнящееся от других. Но поразительно отношение ферментов. Для каждой еды свой сок по ферментам: по белковому ферменту самый сильный - молочный сок, затем идут хлебный и мясной; по крахмальному - самый сильной хлебный и затем молочный и мясной, и по жировому ферменту - очень слабый хлебный и очень сильный молочный, мясной занимает среднее положение. В последних двух случаях приспособление очевидно без дальнейших расследований; для еды с крахмалом усилен крахмаль ный фермент, для еды с жиром - жировой фермент. Это видно уже в колебаниях концентрации, но в особенности - в абсолютном количестве ферментов. Некоторое недоумение может вызвать сначала только первый случай, тe. изменение белкового фермента по родам еды. При желудочной работе мы видели совершенно обратное: на молоко выливался самыйслабый раствор фермента, когда здесь - самый сильный. Однако, принимая во внимание массу сока, мы находим и здесь, что на одно и то же количество белка: хлебного - выливается 1978 единиц белкового фермента, мясного -- 1502 и молочного - только 1085 единиц, т. е. и в случае панкреатического сока растительный белок требует на себя фермента всего больше, а молочный - всего меньше. Разница с желудочными железами оказывается, следовательно, только в том, что при них большее количество фермента доставляется на хлеб в концентрированном растворе, а при поджелудочной железе - в более разжиженном. Как кажется, факт этот придает лишний вес нашему раннему предположению, что в желудке при переваривании хлеба нарочито избегалось накопление большого количества кислоты. Во всяком случае, только что приведенное отношение чрезвычайно усиливает интерес к сложности изучаемого нами механизма; очевидно, почва кишит нерешенными и важными вопросами.
Как и при желудочных железах, работа pancreas. кроме количества и качества сока, характеризуется при всяком сорте еды и ходом ее. Прилагаю числа и соответствующие кривые из работы доктора Вальтера (рис. 8).
Под концентрациею сока разумеется квадрат числа миллиметров растворенных цилиндриков или куб. сантиметров титрованной щелочи, под абсолютным числом ферментных единиц произведение квадрата на число куб. сантиметров выделенного сока. Сравнению подлежат опять эквивалентные количества по азоту. Мы видим, что у каждого сорта еды свое количество сока, резко разнящееся от других. Но поразительно отношение ферментов. Для каждой еды свой сок по ферментам: по белковому ферменту самый сильный - молочный сок, затем идут хлебный и мясной; по крахмальному - самый сильной хлебный и затем молочный и мясной, и по жировому ферменту - очень слабый хлебный и очень сильный молочный, мясной занимает среднее положение. В последних двух случаях приспособление очевидно без дальнейших расследований; для еды с крахмалом усилен крахмаль ный фермент, для еды с жиром - жировой фермент. Это видно уже в колебаниях концентрации, но в особенности - в абсолютном количестве ферментов. Некоторое недоумение может вызвать сначала только первый случай, тe. изменение белкового фермента по родам еды. При желудочной работе мы видели совершенно обратное: на молоко выливался самыйслабый раствор фермента, когда здесь - самый сильный. Однако, принимая во внимание массу сока, мы находим и здесь, что на одно и то же количество белка: хлебного - выливается 1978 единиц белкового фермента, мясного -- 1502 и молочного - только 1085 единиц, т. е. и в случае панкреатического сока растительный белок требует на себя фермента всего больше, а молочный - всего меньше. Разница с желудочными железами оказывается, следовательно, только в том, что при них большее количество фермента доставляется на хлеб в концентрированном растворе, а при поджелудочной железе - в более разжиженном. Как кажется, факт этот придает лишний вес нашему раннему предположению, что в желудке при переваривании хлеба нарочито избегалось накопление большого количества кислоты. Во всяком случае, только что приведенное отношение чрезвычайно усиливает интерес к сложности изучаемого нами механизма; очевидно, почва кишит нерешенными и важными вопросами.
Как и при желудочных железах, работа pancreas. кроме количества и качества сока, характеризуется при всяком сорте еды и ходом ее. Прилагаю числа и соответствующие кривые из работы доктора Вальтера (рис. 8).
 Ввиду всех приведенных фактов и зная два других тканях организма способность более или менее стойко изменяться под долгим влиянием усиленной работы или бездействия, можно было то же самое полагать и при наших железах. В самом деле, направленное на этот пункт исследование поджелудочной железы увенчалось полным успехом. При продолжительном изменении характера еды ферментный состав сока меняется постеленно - день ото дня - все больше и больше. Если, например, исходить из свойств панкреатического сока собаки, несколько недель питавшейся только хлебом и молоком, и затем перевести ее на мясную еду, т. е. на еду с большим содержанием белковых веществ при почти полном отсутствии крахмалистых, то наблюдается постепенное усиление способности переваривать белки. Эта способность все растет и растет с продолжением кормления мясом; способность же переваривать крахмал относится совершенно обратно, т. е. постепенно слабеет. Вот опыт из работы доктора Васильева.
Ввиду всех приведенных фактов и зная два других тканях организма способность более или менее стойко изменяться под долгим влиянием усиленной работы или бездействия, можно было то же самое полагать и при наших железах. В самом деле, направленное на этот пункт исследование поджелудочной железы увенчалось полным успехом. При продолжительном изменении характера еды ферментный состав сока меняется постеленно - день ото дня - все больше и больше. Если, например, исходить из свойств панкреатического сока собаки, несколько недель питавшейся только хлебом и молоком, и затем перевести ее на мясную еду, т. е. на еду с большим содержанием белковых веществ при почти полном отсутствии крахмалистых, то наблюдается постепенное усиление способности переваривать белки. Эта способность все растет и растет с продолжением кормления мясом; способность же переваривать крахмал относится совершенно обратно, т. е. постепенно слабеет. Вот опыт из работы доктора Васильева.
 Собака ежедневно получает две бутылки молока и фунт белого хлеба в продолжение полутора месяцев. Часовые порции сока за первые 6 часов после еды дают следующие цифры: для белкового фермента (в миллиметрах) - 0.0, 0.0, 0.25, 0.25, 0.25, 0.25; для крахмального фермента (в миллиграммах сахара) - 8, 13, 10, 16, 18, 15. Затем эта собака переводится на мясо - полфунта в день. Уже через 3 дня замечается повышение силы белкового фермента и понижение крахмального. На 23-й день мясной диеты эти величины, постепенно изменяясь в указанном смысле, представляются в следующем виде: для белкового фермента (опять же в порциях первых шести часов после еды) - 1.5, 1.0, 1.5, 3.5, 3.5, 3.0; для крахмального фермента соответственно -- 4, 3, 3, 7, 4, 6. При этом нужно заметить, что действие сока на крахмал в последнем опыте продолжалось в 2 раза дольше, чем в первом.
Хотя результат опытов совершеннотчетлив, тем не менее против него было возможно возражение, что та или другая выработка ферментов могла переместиться с одных часов отделительного периода на другие. Поэтому решено было, ради абсолютной точности результатов, сравнить ферментные способности суточных масс сока. Вот трудный опыт, исполненный доктором Яблонским.
Собака, долго питавшаяся мясом и достигшая большой силы в переваривании белков панкреатическим соком, затем переводится на молочно-хлебную диету, причем белковый фермент начинает постепенно убывать, судя по порциям первых шести часов. На 30-й день последней диеты ставится опыт собирания сока за целые сутки. Переваривающая сила этого сока по отношению к белкам выражается (по Мэтту) 4 мм; 10 дней спустя опыт повторяется, и теперь переваривающая сила суточной массы представляется только 2.25 мм. Третий суточный опыт ставится еще на 12 дней позже и дает переваривание в 1.25 мм. Наконец при четвертом опыте, поставленном еще спустя 24 дня, переваривающая сила стала нулем (по Месту). Крахмальный фермент, сперва правильно увеличиваясь, затем представлял неопределенные колебания, однако с наклонностью к понижению. Последний пункт требует, однако, новой проверки. Результат опыта, что касается изменения белкового фермента, не оставляет ничего большего желать. Конечно, важно так же точно проследить изменения и остальных двух ферментов. Более или менее стойкое и с продолжением данного пищевого режима все усиливающееся известное состояние железы можно было изменить на одной и той же собаке и по нескольку раз, в ту или другую сторону, меняя пищевые режимы. Это обстоятельство совершенно исключало подозрение, что в наших опытах имелось дело с какимнибудь самопроизвольным и бесповоротным изменением железы вследствие ли факта операции или какой-либо другой патологической причины.
Собака ежедневно получает две бутылки молока и фунт белого хлеба в продолжение полутора месяцев. Часовые порции сока за первые 6 часов после еды дают следующие цифры: для белкового фермента (в миллиметрах) - 0.0, 0.0, 0.25, 0.25, 0.25, 0.25; для крахмального фермента (в миллиграммах сахара) - 8, 13, 10, 16, 18, 15. Затем эта собака переводится на мясо - полфунта в день. Уже через 3 дня замечается повышение силы белкового фермента и понижение крахмального. На 23-й день мясной диеты эти величины, постепенно изменяясь в указанном смысле, представляются в следующем виде: для белкового фермента (опять же в порциях первых шести часов после еды) - 1.5, 1.0, 1.5, 3.5, 3.5, 3.0; для крахмального фермента соответственно -- 4, 3, 3, 7, 4, 6. При этом нужно заметить, что действие сока на крахмал в последнем опыте продолжалось в 2 раза дольше, чем в первом.
Хотя результат опытов совершеннотчетлив, тем не менее против него было возможно возражение, что та или другая выработка ферментов могла переместиться с одных часов отделительного периода на другие. Поэтому решено было, ради абсолютной точности результатов, сравнить ферментные способности суточных масс сока. Вот трудный опыт, исполненный доктором Яблонским.
Собака, долго питавшаяся мясом и достигшая большой силы в переваривании белков панкреатическим соком, затем переводится на молочно-хлебную диету, причем белковый фермент начинает постепенно убывать, судя по порциям первых шести часов. На 30-й день последней диеты ставится опыт собирания сока за целые сутки. Переваривающая сила этого сока по отношению к белкам выражается (по Мэтту) 4 мм; 10 дней спустя опыт повторяется, и теперь переваривающая сила суточной массы представляется только 2.25 мм. Третий суточный опыт ставится еще на 12 дней позже и дает переваривание в 1.25 мм. Наконец при четвертом опыте, поставленном еще спустя 24 дня, переваривающая сила стала нулем (по Месту). Крахмальный фермент, сперва правильно увеличиваясь, затем представлял неопределенные колебания, однако с наклонностью к понижению. Последний пункт требует, однако, новой проверки. Результат опыта, что касается изменения белкового фермента, не оставляет ничего большего желать. Конечно, важно так же точно проследить изменения и остальных двух ферментов. Более или менее стойкое и с продолжением данного пищевого режима все усиливающееся известное состояние железы можно было изменить на одной и той же собаке и по нескольку раз, в ту или другую сторону, меняя пищевые режимы. Это обстоятельство совершенно исключало подозрение, что в наших опытах имелось дело с какимнибудь самопроизвольным и бесповоротным изменением железы вследствие ли факта операции или какой-либо другой патологической причины.

 Если еда так резко и сильно действует на химический характер железы, то могло быть, что при постоянных природных обстоятельствах или под влиянием продолжительных (всю жизнь длящихся) домашних правил жизни (как это часто, например, практикуется на различных породах собак) должны были выработаться прочные определенные типы панкреатической железы. Наш экспериментальный материал, как нам кажется, действительно дает нам в этом отношении некоторые указания. При совершенно тождественных условиях питания у нас в лаборатории панкреатический сок разных собак часто очень разнится в отношении содержания ферментовветственно этому и переход от одного режима к другому у одних собак дает себя знать очень быстро в свойствах сока, между тем как у других изменение свойств сока наступает и развивается очень медленно. Кроме того, случалось замечать, что в последних случаях резкие переходы от одной еды к другой вели иногда к серьезному заболеванию животных.
Что касается до желудочных желез, то здесь вопрос о хронических изменениях их ферментной способности остается пока открытым. В нашей лаборатории получение чистого желудочного сока путем мнимого кормления производилось на массе собак (их надо считать десятками), и, однако, никогда резко не бросалась в глаза очень большая и постоянная разница в переваривающей силе сока разных собак. Доктор Самойлов (ненапечатанные опыты), ради разъяснения этого вопроса, наблюдал трех гастро- и эзофаготомированных собак, которые после многократного испытания путем мнимого кормления были посажены на различные пищевые режимы. После долгого времени никакого резкого указания на изменение состава сока, добываемого тем же путем мнимого кормления, не оказалось. Как относиться к этому результату? Неблагоприятен ли был способ суждения о ферментной способности желудочных желез или в самом деле на этом пункте желудочные железы существенно отличаются от поджелудочной железы? Конечно, возможно, что панкреатическая железа в значительной степени играет роль дополнительной резервной железы, которая, смотря по бремени, лежащему на пищеварительном канале, в силу своей специальной натуры, в особенности способна то усиливать, то ослаблять свою работу, между тем как желудочные железы, будучи первой сильной пищеварительной инстанцией, обязаны поэтому постоянно работать в максимальном размере их сил. Только в последнее время в нашей лаборатории (д-ром Лобановым) получается факт, правда, не простой для толкования, но как будто говорящий за стойкое изменение и желудочных желез при том или другом пищевом продолжительном режиме. Мы имеем собаку, у которой уединен кусок дна желудка по способу Гейденгайна, т. е. с перерезкой блуждающих нервов. Нужно сказать, что у таких собак, раз они выживают более или менее продолжительное время, отделение желудочного сока мало-помалу делается очень незначительным (наблюдение нашей лаборатории). На такой собаке было замечено следующее отношение. Когда собака надолго была посажена на обильную мясную пищу, у нее всякий раз затем, при испытании одним и тем же приемом, т. е. одной и той же едой в одном и том же количестве, достигалось гораздо более обильное отделение, чем когда животное питалось иначе, например хлебом с молоком или овсянкой. Ввиду, однако, очевидного нарушения нормальных условий работы желез у оперированных так животных нельзя уверенно опираться на описанный факт.
Приведенная сумма фактов, надеюсь, достаточно оправдывает уже сделанное выше и теперь повторяемое еще раз заключение, что работа исследованных желез весьма сложна, эластична, вместе с тем удивительно точна и, конечно, целесообразна, хотя мы эту целесообразность в настоящее время усматриваем бесслорно только в отдельных случаях.
Если еда так резко и сильно действует на химический характер железы, то могло быть, что при постоянных природных обстоятельствах или под влиянием продолжительных (всю жизнь длящихся) домашних правил жизни (как это часто, например, практикуется на различных породах собак) должны были выработаться прочные определенные типы панкреатической железы. Наш экспериментальный материал, как нам кажется, действительно дает нам в этом отношении некоторые указания. При совершенно тождественных условиях питания у нас в лаборатории панкреатический сок разных собак часто очень разнится в отношении содержания ферментовветственно этому и переход от одного режима к другому у одних собак дает себя знать очень быстро в свойствах сока, между тем как у других изменение свойств сока наступает и развивается очень медленно. Кроме того, случалось замечать, что в последних случаях резкие переходы от одной еды к другой вели иногда к серьезному заболеванию животных.
Что касается до желудочных желез, то здесь вопрос о хронических изменениях их ферментной способности остается пока открытым. В нашей лаборатории получение чистого желудочного сока путем мнимого кормления производилось на массе собак (их надо считать десятками), и, однако, никогда резко не бросалась в глаза очень большая и постоянная разница в переваривающей силе сока разных собак. Доктор Самойлов (ненапечатанные опыты), ради разъяснения этого вопроса, наблюдал трех гастро- и эзофаготомированных собак, которые после многократного испытания путем мнимого кормления были посажены на различные пищевые режимы. После долгого времени никакого резкого указания на изменение состава сока, добываемого тем же путем мнимого кормления, не оказалось. Как относиться к этому результату? Неблагоприятен ли был способ суждения о ферментной способности желудочных желез или в самом деле на этом пункте желудочные железы существенно отличаются от поджелудочной железы? Конечно, возможно, что панкреатическая железа в значительной степени играет роль дополнительной резервной железы, которая, смотря по бремени, лежащему на пищеварительном канале, в силу своей специальной натуры, в особенности способна то усиливать, то ослаблять свою работу, между тем как желудочные железы, будучи первой сильной пищеварительной инстанцией, обязаны поэтому постоянно работать в максимальном размере их сил. Только в последнее время в нашей лаборатории (д-ром Лобановым) получается факт, правда, не простой для толкования, но как будто говорящий за стойкое изменение и желудочных желез при том или другом пищевом продолжительном режиме. Мы имеем собаку, у которой уединен кусок дна желудка по способу Гейденгайна, т. е. с перерезкой блуждающих нервов. Нужно сказать, что у таких собак, раз они выживают более или менее продолжительное время, отделение желудочного сока мало-помалу делается очень незначительным (наблюдение нашей лаборатории). На такой собаке было замечено следующее отношение. Когда собака надолго была посажена на обильную мясную пищу, у нее всякий раз затем, при испытании одним и тем же приемом, т. е. одной и той же едой в одном и том же количестве, достигалось гораздо более обильное отделение, чем когда животное питалось иначе, например хлебом с молоком или овсянкой. Ввиду, однако, очевидного нарушения нормальных условий работы желез у оперированных так животных нельзя уверенно опираться на описанный факт.
Приведенная сумма фактов, надеюсь, достаточно оправдывает уже сделанное выше и теперь повторяемое еще раз заключение, что работа исследованных желез весьма сложна, эластична, вместе с тем удивительно точна и, конечно, целесообразна, хотя мы эту целесообразность в настоящее время усматриваем бесслорно только в отдельных случаях.
 Во всех этих случаях количество сока при редкой даче кусков гораздо меньше, чем при сплошной. Из этих данных следует, вопервых, что в блуждающем нерве находятся специальные нервные волокна желудочных желез, а не сосудистые, и, во-вторых, что эти специальные волокна также нужно подразделять на секреторные и трофические, как это установлено Гейденгайном для слюнной иннервации, потому что выделение воды и выработка специальных веществ, очевидно, происходят независимо друг от друга. Массу доказательств тому же вы видели уже на второй лекции, где зачастую одни и те же часовые количества сока при различных условиях деятельности желез выливались с чрезвычайно различным содержанием фермента.
Как ни убедительно само по себе доказательство существования отдельных нервов желудка путем их перерезки, по многим основаниям является желательным применение и способа раздражения. Лишь искусственное раздражение нерва дает возможность подробно и точно изучить как действие нерва, так и заведуемый им процесс. В данном случае при опыте восстают большие трудности, которыми и объясняется горькая неудача огромного большинства авторов, занимавшихся предметом ранее. Мы исполнили этот опыт, обставив его совершенно особенным образом. Мы вышли из сомнения: едва ли обыкновенная форма острого, т. е. сейчас, без особенного приготовления, на свежем животном исполненного, физиологического опыта могла с правом претендовать на сохранение нормальных отношений в организме; при ней, наверное, многие физиологические явления искажаются и маскируются. В нашем случае сомнения были тем уместнее, что в науке уже имелись бесспорные факты резко задерживающего влияния болевых или вообще рефлекторных раздражений на деятельность главных пищеварительных желез. Бернштейн [19] в лаборатории Людвига и затем мы с профессором Афанасьевым 2 [20] показали, что чувствительные раздражения отчетливо, и часто надолго, тормозят отделительную работу поджелудочной железы. Доктор Нечаев [21] видел, что 2-3-минутное раздражение седалищного нерва совершенно останавливает пищеварение в желудке на многие часы. Отсюда естественно вытекало требование раздражать нерв, идущий в желудок, таким образом, чтобы этому раздражению не предшествовали и, тем более, его не сопровождали какие-нибудь значительные чувствительные или вообще рефлекторные раздражения.
Мы достигли этого вместе с г-жой Шумовой-Симановской, когда у собак, совершенно подобных только что вам показанной, т. е. заранее гастро- и эзофаготомированных, с перерезанным, под n. laryngeus inferior и сердечными ветвями, правым блуждающим нервом, перерезали, как сегодня, левый блуждающий нерв на шее и, отпрепаровав более или менее длинный периферический конец этого нерва, оставляли его на нитке прямо под кожей раны на некоторое время. Спустя 3-4 дня, при осторожном снимании швов кожи, рана легко раскрывалась и мы имели перед собой нерв. Таким образом раздражению нерва не предшествовало причинение сколько-нибудь значительной боли животному. Ценою таких приготовлений было достигнуто, что теперь всякий раз при раздражении нерва редкими (через 1-2 секунды) индукционными ударами (так называемое ритмическое раздражение) мы гнали из совершенно пустого желудка чистый желудочный сок.
Овладев предметом, можно было попытаться добиться того же успеха и на остром опыте, т. е. на животном, сейчас же приготовленном к опыту, конечно приняв некоторые особенные меры. Доктор Ушаков в своих первых опытах после спешной, но осторожной трахеотомии производил возможно быстро (в несколько секунд) перерезку спинного мозга прямо под продолговатым, чтобы в дальнейшем оперировании быть свободным от страха рефлекторных влияний на желудочные железы. Затем отпрепаровывались и перерезывались блуждающие нервы, в желудок вставлялась обыкновенная желудочная фистульная трубка, а на пищевод на шее и на pylorus накладывались крепкие лигатуры. После всего этого животное подвешивалось в станке в стоячем положении. В позднейших опытах доктором Ушаковым применялось кратковременное хлороформирование, причем в короткий период b продолжающегося наркоза (10-15 минут) спешили исполнить всю только что списанную оперативную часть опыта.
Нарочные опыты на собаке с гастро- и эзофаготомией показали, что хлороформирование такой продолжительности не влечет за собою сколько-нибудь значительного паралича желез и их нервов, так как 15-20 минут спустя после наркоза оправившееся животное с жадностью ест предлагаемую ему пищу, и из пустого желудка, через обычную паузу в 5 минут, начинает вытекать в нормальном количестве вполне деятельный сок. Теперь в остром опыте приступали к раздражению нервов и, согласно расчету, получили возможность видеть бесспорное и резкое секреторное действие раздражения, но, однако, только в половине всех опытов. При этом обращало на себя внимание, что положительный результат в последних опытах, с применением наркоза, встречался чаще, чем в первых. Во всех удачных случаях действие раздражения, однако, никогда не обнаруживалось сейчас же, всегда проходил известный период, от 15 минут до часу и больше, в который раздражение оставалось бесплодным. Когда, наконец, нерв начинал действовать, секреторное действие с прекращением раздражения постепенно исчезало; с повторением раздражения, теперь уже скоро, через несколько минут, возвращалось. При отравлении животного секреторнозадерживающим ядом - атропином нервы теряли свое действие. Факт предварительного длинного, бесплодного периода раздражения, объясняясь отчасти пониженной, вследствие оперирования, возбудимостью желез, для полного своего понимания требует, однако, другого толкования.
Как уже сказано, после наркоза мнимое кормление очень скоро дает совершенно нормальный результат относительно отделения сока. Между тем в острых опытах, поставленных с наркозом, скрытый период при раздражении нерва остается также долгим Считать значительным задерживающий рефлекс на железу со стороны оперирования во время наркоза и после перерезки спинного мозга едва ли было бы основательно. В таком случае приходится допустить, что при искусственном раздражении блуждающих нервов к железе посылаются как возбуждающие, так и задерживающие влияния. Всего проще это допущение выливалось бы в форму гипотезы о секреторнозадерживающих нервах, антагонистах секреторных нервов, как существуют антагонисты в области сосудистой, сердечной и других иннерваций. Ближе этой гипотезой мы займемся при поджелудочной железе, где для принятия ее существует уже достаточный фактический материал и даже, в последнее время, - прямые доказательства.
Итак, обе наши формы опыта, хроническая и острая, дают нам полное право признать в блуждающем нерве секреторный нерв желудочных желез. Однако, повторяем еще раз, не надо думать, что целость блуждающих нервов есть единственное условие отделительной работы желез. Как многие авторы до нас, так и мы имели случай с несомненностью убеждаться, что желудок способен к выработке своего специального секрета и без блуждающих нервов; при этом, однако, эта работа обладает известными особенностями как относительно условий ее наступления, так и характера ее продукта. Что это за отделение в отсутствии блуждающих нервов: нервное ли оно (через симпатическую систему) или какого другого происхождения, сказать сейчас вполне определенно еще нельзя. Впрочем, профессор Саноцкий на уединенном желудочке по способу Гейденгайна (значит, с перерезанными блуждающими нервами) вполне резко показал задерживающее действие атропина атропин есть специальный парализатор секреторной иннервации. Можно надеяться, что дальнейшие исследования, направленные на симпатическую систему, теперь, при знании отношения блуждающих нервов к желудочным железам, скоро разъяснят дело окончательно.
Нельзя не дать здесь места сожалению, что успевший сделаться привычным физиологический взгляд о независимости желудочных желез от нервов продолжает игнорировать вышеприведенные результаты, несмотря на то, что главнейшие из них опубликованы уже целых 7 лет тому назад не только у нас, но и в заграничной научной прессе. Часть авторов опирается на продолжение секреторной деятельности желез после перерезки блуждающих нервов, но при этом не желает знать особенностей этой деятельности, которыми в данном случае все и определяется. Ведь перерезка нервов и многих других органов не прекращает окончательно специальной деятельности этих органов, а это не дает права утверждать, что нет никакой иннервации этих органов. Другие авторы продолжают упорствовать на традиционной обстановке острого опыта, т. е. не принимают никаких мер предосторожности против рефлекторной задержки. Лишь некоторые авторы (Аксенфельд, Контежан, Шнейер) на собаких и других животных (птицах и лягушках) получили более или менее положительные результаты. Мы смеем верить, что всякая поверка наших опытов, при показанных нами условиях, во всяких руках даст то же самое и не оставит места ни малейшему сомнению относительно существования секреторной иннервации желудочных желез.
Во всех этих случаях количество сока при редкой даче кусков гораздо меньше, чем при сплошной. Из этих данных следует, вопервых, что в блуждающем нерве находятся специальные нервные волокна желудочных желез, а не сосудистые, и, во-вторых, что эти специальные волокна также нужно подразделять на секреторные и трофические, как это установлено Гейденгайном для слюнной иннервации, потому что выделение воды и выработка специальных веществ, очевидно, происходят независимо друг от друга. Массу доказательств тому же вы видели уже на второй лекции, где зачастую одни и те же часовые количества сока при различных условиях деятельности желез выливались с чрезвычайно различным содержанием фермента.
Как ни убедительно само по себе доказательство существования отдельных нервов желудка путем их перерезки, по многим основаниям является желательным применение и способа раздражения. Лишь искусственное раздражение нерва дает возможность подробно и точно изучить как действие нерва, так и заведуемый им процесс. В данном случае при опыте восстают большие трудности, которыми и объясняется горькая неудача огромного большинства авторов, занимавшихся предметом ранее. Мы исполнили этот опыт, обставив его совершенно особенным образом. Мы вышли из сомнения: едва ли обыкновенная форма острого, т. е. сейчас, без особенного приготовления, на свежем животном исполненного, физиологического опыта могла с правом претендовать на сохранение нормальных отношений в организме; при ней, наверное, многие физиологические явления искажаются и маскируются. В нашем случае сомнения были тем уместнее, что в науке уже имелись бесспорные факты резко задерживающего влияния болевых или вообще рефлекторных раздражений на деятельность главных пищеварительных желез. Бернштейн [19] в лаборатории Людвига и затем мы с профессором Афанасьевым 2 [20] показали, что чувствительные раздражения отчетливо, и часто надолго, тормозят отделительную работу поджелудочной железы. Доктор Нечаев [21] видел, что 2-3-минутное раздражение седалищного нерва совершенно останавливает пищеварение в желудке на многие часы. Отсюда естественно вытекало требование раздражать нерв, идущий в желудок, таким образом, чтобы этому раздражению не предшествовали и, тем более, его не сопровождали какие-нибудь значительные чувствительные или вообще рефлекторные раздражения.
Мы достигли этого вместе с г-жой Шумовой-Симановской, когда у собак, совершенно подобных только что вам показанной, т. е. заранее гастро- и эзофаготомированных, с перерезанным, под n. laryngeus inferior и сердечными ветвями, правым блуждающим нервом, перерезали, как сегодня, левый блуждающий нерв на шее и, отпрепаровав более или менее длинный периферический конец этого нерва, оставляли его на нитке прямо под кожей раны на некоторое время. Спустя 3-4 дня, при осторожном снимании швов кожи, рана легко раскрывалась и мы имели перед собой нерв. Таким образом раздражению нерва не предшествовало причинение сколько-нибудь значительной боли животному. Ценою таких приготовлений было достигнуто, что теперь всякий раз при раздражении нерва редкими (через 1-2 секунды) индукционными ударами (так называемое ритмическое раздражение) мы гнали из совершенно пустого желудка чистый желудочный сок.
Овладев предметом, можно было попытаться добиться того же успеха и на остром опыте, т. е. на животном, сейчас же приготовленном к опыту, конечно приняв некоторые особенные меры. Доктор Ушаков в своих первых опытах после спешной, но осторожной трахеотомии производил возможно быстро (в несколько секунд) перерезку спинного мозга прямо под продолговатым, чтобы в дальнейшем оперировании быть свободным от страха рефлекторных влияний на желудочные железы. Затем отпрепаровывались и перерезывались блуждающие нервы, в желудок вставлялась обыкновенная желудочная фистульная трубка, а на пищевод на шее и на pylorus накладывались крепкие лигатуры. После всего этого животное подвешивалось в станке в стоячем положении. В позднейших опытах доктором Ушаковым применялось кратковременное хлороформирование, причем в короткий период b продолжающегося наркоза (10-15 минут) спешили исполнить всю только что списанную оперативную часть опыта.
Нарочные опыты на собаке с гастро- и эзофаготомией показали, что хлороформирование такой продолжительности не влечет за собою сколько-нибудь значительного паралича желез и их нервов, так как 15-20 минут спустя после наркоза оправившееся животное с жадностью ест предлагаемую ему пищу, и из пустого желудка, через обычную паузу в 5 минут, начинает вытекать в нормальном количестве вполне деятельный сок. Теперь в остром опыте приступали к раздражению нервов и, согласно расчету, получили возможность видеть бесспорное и резкое секреторное действие раздражения, но, однако, только в половине всех опытов. При этом обращало на себя внимание, что положительный результат в последних опытах, с применением наркоза, встречался чаще, чем в первых. Во всех удачных случаях действие раздражения, однако, никогда не обнаруживалось сейчас же, всегда проходил известный период, от 15 минут до часу и больше, в который раздражение оставалось бесплодным. Когда, наконец, нерв начинал действовать, секреторное действие с прекращением раздражения постепенно исчезало; с повторением раздражения, теперь уже скоро, через несколько минут, возвращалось. При отравлении животного секреторнозадерживающим ядом - атропином нервы теряли свое действие. Факт предварительного длинного, бесплодного периода раздражения, объясняясь отчасти пониженной, вследствие оперирования, возбудимостью желез, для полного своего понимания требует, однако, другого толкования.
Как уже сказано, после наркоза мнимое кормление очень скоро дает совершенно нормальный результат относительно отделения сока. Между тем в острых опытах, поставленных с наркозом, скрытый период при раздражении нерва остается также долгим Считать значительным задерживающий рефлекс на железу со стороны оперирования во время наркоза и после перерезки спинного мозга едва ли было бы основательно. В таком случае приходится допустить, что при искусственном раздражении блуждающих нервов к железе посылаются как возбуждающие, так и задерживающие влияния. Всего проще это допущение выливалось бы в форму гипотезы о секреторнозадерживающих нервах, антагонистах секреторных нервов, как существуют антагонисты в области сосудистой, сердечной и других иннерваций. Ближе этой гипотезой мы займемся при поджелудочной железе, где для принятия ее существует уже достаточный фактический материал и даже, в последнее время, - прямые доказательства.
Итак, обе наши формы опыта, хроническая и острая, дают нам полное право признать в блуждающем нерве секреторный нерв желудочных желез. Однако, повторяем еще раз, не надо думать, что целость блуждающих нервов есть единственное условие отделительной работы желез. Как многие авторы до нас, так и мы имели случай с несомненностью убеждаться, что желудок способен к выработке своего специального секрета и без блуждающих нервов; при этом, однако, эта работа обладает известными особенностями как относительно условий ее наступления, так и характера ее продукта. Что это за отделение в отсутствии блуждающих нервов: нервное ли оно (через симпатическую систему) или какого другого происхождения, сказать сейчас вполне определенно еще нельзя. Впрочем, профессор Саноцкий на уединенном желудочке по способу Гейденгайна (значит, с перерезанными блуждающими нервами) вполне резко показал задерживающее действие атропина атропин есть специальный парализатор секреторной иннервации. Можно надеяться, что дальнейшие исследования, направленные на симпатическую систему, теперь, при знании отношения блуждающих нервов к желудочным железам, скоро разъяснят дело окончательно.
Нельзя не дать здесь места сожалению, что успевший сделаться привычным физиологический взгляд о независимости желудочных желез от нервов продолжает игнорировать вышеприведенные результаты, несмотря на то, что главнейшие из них опубликованы уже целых 7 лет тому назад не только у нас, но и в заграничной научной прессе. Часть авторов опирается на продолжение секреторной деятельности желез после перерезки блуждающих нервов, но при этом не желает знать особенностей этой деятельности, которыми в данном случае все и определяется. Ведь перерезка нервов и многих других органов не прекращает окончательно специальной деятельности этих органов, а это не дает права утверждать, что нет никакой иннервации этих органов. Другие авторы продолжают упорствовать на традиционной обстановке острого опыта, т. е. не принимают никаких мер предосторожности против рефлекторной задержки. Лишь некоторые авторы (Аксенфельд, Контежан, Шнейер) на собаких и других животных (птицах и лягушках) получили более или менее положительные результаты. Мы смеем верить, что всякая поверка наших опытов, при показанных нами условиях, во всяких руках даст то же самое и не оставит места ни малейшему сомнению относительно существования секреторной иннервации желудочных желез.
 Те же затруднения, с которыми пришлось бороться при исследовании иннервации желудочных желез, долго давали себя знать и при поджелудочной железе. Для характеристики этих затруднений позволяю себе привести из классической статьи Гейденгайна о поджелудочной железе следующее, очень выразительное место: «Наверное всякий наблюдатель, который занимался функциею поджелудочной железы более долгое время, оставит эту область с недовольным чувством, потому что он принужден был выбрасывать огромное число неудачных опытов. Ни крайняя осторожность, ни большая опытность не могут победить невероятную чувствительность органа, который чрезвычайно часто по совершении предварительной операции прекращает свою деятельность и не возвращается к ней, несмотря на применение действительнейших средств. Таким образом наблюдению здесь всегда присуща известная неверность, которую не в состоянии устранить ни крайнее разнообразие, ни увеличение массы опытов. Я должен откровенно признаться, что еще ни разу не предпринимал таких опытов, столь богатых собачьими жертвами и бедных соответственными результатами». 1 [22] Зато в настоящее время изучение нервных отношений этой железы во многих отношениях подвинулось сильно вперед. Как уже упомянуто, всего раньше (в лаборатории Людвига Бернштейном, затем мною вместе с профессором М. Афанасьевым) было показано задерживающее действие чувствительного раздражения на деятельность железы, потом Гейденгайну и его ученику Ландау [23] удалось при раздражении продолговатого мозга, в некоторых опытах среди многих неудачных, обнаружить бесспорное возбуждающее действие на железу. Однако весь вопрос об иннервации все еще оставался очень темным. Почему действие у Гейденгайна наблюдалось только в исключительных случаях? Каким именно нервам проводится возбуждающее действие из центральной нервной системы к железе? На чем основано задерживающее действие чувствительных раздражений? Все это оставалось без малейшего ответа. Начиная с 1887 г., мне, вместе с моими сотрудниками, посчастливилось более или менее разъяснить все выставленные вопросы.
Секреторным нервом поджелудочной железы оказался блуждающий нерв. Этот результат дался нам благодаря применению некоторых особенных обстановок опыта. Обстановку, в которой впервые пред нами объявилось действие этого нерва, я имею честь продемонстрировать сейчас. Вот собака, у которой была наложена постоянная панкреатическая фистула по способу, описанному в первой лекции. Собака совершенно оправилась от операции, все давно срослось. Четыре дня тому назад ей на шее перерезан блуждающий нерв, отпрепарован периферический конец его, взят на нитку и оставлен с ней прямо под кожей. Сейчас, сняв осторожно кожные швы, я с легкостью, без причинения какой-нибудь заметной неприятности животному, достаю нитку с нервом. Прошу обратить внимание, что из металлической воронки, прижатой широким краем к брюшной стенке с отверстием панкреатического протока, не вытекает ни капли сока. Я начинаю теперь раздражать прерывистым электрическим током мой нерв. Как видите, животное стоит совершенно спокойно в станке, не обнаруживая и сейчас ни малейшим движением никакой боли. Проходит целых 2 минуты без всякого действия на железу (что и прошу вас особенно удержать в памяти), и только в третью появляется первая капля сока, за которою следуют дальнейшие, все чаще и чаще. После трех минут я заканчиваю раздражение, сок продолжает течь сам по себе и постепенно затихает только через 4-5 минут по окончании раздражения. Повторяя раздражение еще раз, как вы видите, мы получаем совершенно то же самое. И так всегда и на всех собаках. Нужно прибавить, что блуждающий нерв, конечно, раздражали и раньше с той же целью, что и мы, и, однако, не имели того, что теперь так легко показывается даже публично. Причина нашей удачи - некоторые особенности в постановке опыта. Этих особенностей две: животное во время опыта не подвергается никаким болезненным раздражениям, не говоря уже об отравлениях, большей частью сопровождающих обычную форму опытов; с другой стороны, при раздражении блуждающего нерва на шее, благодаря перерезке его за 4 дня до опыта, исключены резкие расстройства кровообращения. К четвертому дню перерезки замедляющие волокна сердца настолько потеряли в своей возбудимости вследствие развивающегося перерождения нерва, что самое сильное раздражение нерва едва дает себя знать весьма незначительным и мимолетным замедлением ударов сердца. Для понимания этой стороны опыта нужно не забывать, что возбудимость различных нервов после перерезки исчезает через различные сроки - и, следовательно, сейчас уас замедляющие волокна сердца потеряли свою возбудимость ранее, чем секреторные. Итак, в нашем опыте железы нисколько не пострадали ни от операции, ни от обстоятельств, сопутствующих раздражению.
Однако постоянного результата с раздражением блуждающего нерва можно достигнуть и в остром опыте, если только он ведется по некоторому определенному плану. Дело делается так. На нормальном животном быстро и возможно деликатно делается трахеотомия, затем в несколько секунд отделяется продолговатый мозг от спинного и вводится искусственное дыхание. Теперь можно спокойнее приступить к дальнейшему оперированию: вскрытию грудной полости для отпрепарования блуждающих нервов ниже сердца и вскрытию брюшной полости для вставления стеклянной трубочки в проток панкреатической железы. При этих условиях во всех опытах также наблюдается положительное секреторное действие блуждающего нерва на поджелудочную железу, хотя в начале опыта приходится частенько раздражать нерв несколько раз без видимого эффекта. Смысл указанных условий очевиден. Благодаря перерезке спинного мозга устраняется вредное рефлекторно-задерживающее действие дальнейшего длительного оперирования, a раздражение блуждающего нерва в грудной полости совершенно исключает изменения в ритмесердца.
Дальнейший анализ на этой второй форме опыта выдвинул два обстоятельства, от которых приощи нервов может произойти задерживание секреторной энергии поджелудочной железы. В наших опытах, как и в ранних опытах некоторых других авторов, панкреатическая железа оказалась резко чувствительной в отношении кровообращения. Достаточно кратковременного (2-3 минуты) раздражения сосудосуживающих ее нервов или зажатия аорты такой же продолжительности, чтобы железа перестала реагировать в продолжение некоторого периода времени на раздражения блуждающих нервов, ранее действительные. Эти опыты уже в значительной степени делают понятным, почему после обыкновенной операции, сопровождающейся сильнейшим чувствительным раздражением и, следовательно, рефлекторным сужением сосудов, железа животного, находящегося в разгаре пищеварения, сплошь и рядом не дает ни капли сока. Еще большее значение надо приписать другому фактору, который постоянно привлекал к себе внимание в наших опытах. Как в том опыте, который был проделат перед вами, так и в острых опытах раздражение блуждающего нерва не вызывает отделения сока моментально, а всегда проходит более или менее значительное время (от 15 сек. до нескольких минут) между моментом приложения раздражителя к нерву и наступлением отделительного эффекта. Сплошь и рядом бывает так, что сок начинает вытекать лишь в тот момент, когда прекращается раздражение. Наконец часто приходится наблюдать следующее особенное явление (д-р Метт). Вы, положим, долгое время раздражали правый блуждающий нерв и уже имеете от него равномерное отделение сока. Стоит в это время, не прерывая прежнего раздражения, присоединить раздражение другого блуждающего нерва для того, чтобы ток сока сейчас же остановился на известный, часто довольно значительный период времени.
Все эти явления вели к заключению, что в блуждающих нервах, вместе с возбуждающими элементами для pancreas, имеется и нечто ее тормозящее. Об этих тормозящих элементах можно сделать несколько предположений: они могут быть сосудосуживающими нервами поджелудочной железы, двигательными нервами мускулов ее каналов и, наконец, истинными секреторнозадерживающими нервами как антагонистами секреторных. Если при многих органах несомненно доказано существование антагонистической пары управляющих органом нервов, то почему же не быть этому и при железах? Может быть даже, этот антагонизм есть общий принцип всех иннерваций. Факты, намекающие на существование секреторнозадерживающих нервов желез, встречаются изредка в физиологической литературе последних лет. Но, мне кажется, вопрос об их существовании получит окончательное разрешение именно при исследовании иннервации желудка и поджелудочной железы, так как здесь задерживающие явления выражены наиболее резко. Ранее подробного анализа этого вопроса, я приведу опыты, относящиеся до секреторного действия на поджелудочную железу симпатического нерва, так как они доставят с своей стороны некоторый материал для обсуждения интересующего нас вопроса. Вот результаты проф. Кудрявицкого.
Если в описанной выше острой форме опыта с нашей железой раздражать обыкновенным прерывистым индукционным током симпатический нерв, то только в самый первый момент раздражения замечается маленькое толчкообразное движение сока, а затем во все время раздражения и после него - ни малейшего отделения. Применяя же механическое раздражение (ряд толчков посредством тетаномотора Гейденгайна) вместо электрического, часто видят другой результат: спустя некоторое время после начала раздражения происходит довольно сильное отделение сока. Можно того же достигнуть и электрическим раздражением, если раздражать нерв не свежий, а перерезанный за 4-5 дней до этого и, следовательно, подвергающийся перерождению. Смысл этих явлений легко понять, если припомнить некоторые пункты из физиологии сосудодвигательных нервов. Известно, что эти нервы мало чувствительны к механическому раздражению, а после их перерезки ранее многих других волокон теряют свою возбудимость. Следовательно, мы имеем право принимать: 1) что в симпатическом нерве одновременно находятся как суживающие, так и отделительные нервы поджелудочной железы, 2) что при обыкновенном электрическом раздражении свежего нерва сосудосуживающие волокна вполне маскируют отделительные и 3) что лишь при особенных условиях (механическое раздражение и электрическое раздражение заранее перерезанного нерва), устраняющих вмешательства сосудосуживающих волокон, отделительные волокна получают возможность заявить о своем существовании.
Таким образом симпатический нерв представил нам удобный случай установить взаимное отношение сосудодвигательных и секреторных нервов поджелудочной железы. Однако применение указанных приемов при раздражении блуждающего нерва нисколько не изменило картины его действия на нашу железу: задерживающее действие блуждающего нерва осталось при этом в полной целости. И это дает хорошее основание думать, что тормозящее действие блуждающего нерва не обусловливается сужением кровеносных сосудов, самое последнее время д-р Попельский сильно подвинулся в решении занимающего нас вопроса. Им прежде всего была выработана форма опыта, при которой задерживающее по отношению к поджелудочной железе действие блуждающего нерва выступает постоянно и притом в очень резком виде. На острой форме опыта (как описано раньше) вливается в двенадцатиперстную кишку раствор соляной кислоты. Этим вызывается продолжительное и обильное отделение панкреатического сока. Сильное раздражение в это время блуждающих нервов всякий раз без исключения сейчас же обусловливает замедление, а чаще совершенное прекращение отделения. Раздражение же симпатического нерва только замедляет отделение и притом лишь спустя некоторое время после начала раздражения. Точно так же зажатие аорты останавливает отделение только через минуту, две, три. При этом нельзя не упомянуть еще, что, по последним опытам Франсуа-Франка, блуждающий нерв скорее расширяет сосуды поджелудочной железы, чем сужает их. Возможность вмешательства двигательных нервов протоков железы исключалась тем, что животное отравлялось физостигмином, сильнейшим раздражителем гладкой мускулатуры, причем, однако, не только не оказалось никакого задерживания сока, а скорее ядом было усилено отделение. Наконец при подробном препаровании нервов железы удалось найти такие ветви, которые при раздражении вызывали отделение без длинного латентного периода, почти так же быстро, как барабанная струна гонит слюну. Из последнего опыта нужно заключить, что в данном пункте секреторные волокна поджелудочной железы анатомически отделились от задерживающих элементов и что, следовательно, чистым секреторным нервам не принадлежит особенность обусловливать деятельность органа при искусственном раздражении лишь после продолжительного периода скрытого действия. Д-р Попельский нашел, наконец, в отдельности и такие ветви блуждающего нерва, которые только задерживали, но никогда не возбуждали отделения поджелудочного сока. Конечно, раз существуют такие специально задерживающие нервы, то вполне возможны и рефлекторные раздражения их как при нормальных условиях, так и при оперировании. Не исключается, наконец, возможность рефлекторного задерживания центров самих секреторных нервов pancreas.
Из приведенных наблюдений точно и фактически объясняются все неудачи и трудности прежних исследований над иннервациею поджелудочной железы. Например, почему Гейденгайн при раздражении продолговатого мозга получил положительный результат только в немногих опытах? Не говоря о задерживающем влиянии операции, раздражая мозг, он вызывал и сильное сужение сосудов и нарушение деятельности сердца, а к тому же оставалось в силе и раздражение антагонистических волокон.
Вы уже, конечно, заметили, до чего сходно идут нервные явления на желудочных и панкреатических железах: во всех стношениях иннервация одних есть копия других. Вот почему недостающее в одной иннервации против другой можно с правом восполнить по аналогии. На этом основании мы не можем сомневаться, например, в том, что секреторные волокна желез, помимо блуждающих нервов, находятся и в симпатическом нерве.
В заключение несколько слов о вышеупомянутом опыте двух французских авторов над желудком обезглавленного преступника. После знакомства с фактами крайней щепетильности наших желез нелегко поверить выводу авторов, что они имели перед собою истинный отделительный эффект блуждающего нерва 40 минут спустя после обескровления органа.
Мне кажется, я могу верить, что после всего сообщенного и показанного вам представляются так же бесспорными и действительными отделительные нервы желудочных и поджелудочной желез, как классическая, всем известная chorda tympani при слюнных железах. Само собой разумеется, что, кроме этих специальных нервов, в наши железы входят и сосудистые нервы, сосудосуживающие и сосудорасширяющие.
Те же затруднения, с которыми пришлось бороться при исследовании иннервации желудочных желез, долго давали себя знать и при поджелудочной железе. Для характеристики этих затруднений позволяю себе привести из классической статьи Гейденгайна о поджелудочной железе следующее, очень выразительное место: «Наверное всякий наблюдатель, который занимался функциею поджелудочной железы более долгое время, оставит эту область с недовольным чувством, потому что он принужден был выбрасывать огромное число неудачных опытов. Ни крайняя осторожность, ни большая опытность не могут победить невероятную чувствительность органа, который чрезвычайно часто по совершении предварительной операции прекращает свою деятельность и не возвращается к ней, несмотря на применение действительнейших средств. Таким образом наблюдению здесь всегда присуща известная неверность, которую не в состоянии устранить ни крайнее разнообразие, ни увеличение массы опытов. Я должен откровенно признаться, что еще ни разу не предпринимал таких опытов, столь богатых собачьими жертвами и бедных соответственными результатами». 1 [22] Зато в настоящее время изучение нервных отношений этой железы во многих отношениях подвинулось сильно вперед. Как уже упомянуто, всего раньше (в лаборатории Людвига Бернштейном, затем мною вместе с профессором М. Афанасьевым) было показано задерживающее действие чувствительного раздражения на деятельность железы, потом Гейденгайну и его ученику Ландау [23] удалось при раздражении продолговатого мозга, в некоторых опытах среди многих неудачных, обнаружить бесспорное возбуждающее действие на железу. Однако весь вопрос об иннервации все еще оставался очень темным. Почему действие у Гейденгайна наблюдалось только в исключительных случаях? Каким именно нервам проводится возбуждающее действие из центральной нервной системы к железе? На чем основано задерживающее действие чувствительных раздражений? Все это оставалось без малейшего ответа. Начиная с 1887 г., мне, вместе с моими сотрудниками, посчастливилось более или менее разъяснить все выставленные вопросы.
Секреторным нервом поджелудочной железы оказался блуждающий нерв. Этот результат дался нам благодаря применению некоторых особенных обстановок опыта. Обстановку, в которой впервые пред нами объявилось действие этого нерва, я имею честь продемонстрировать сейчас. Вот собака, у которой была наложена постоянная панкреатическая фистула по способу, описанному в первой лекции. Собака совершенно оправилась от операции, все давно срослось. Четыре дня тому назад ей на шее перерезан блуждающий нерв, отпрепарован периферический конец его, взят на нитку и оставлен с ней прямо под кожей. Сейчас, сняв осторожно кожные швы, я с легкостью, без причинения какой-нибудь заметной неприятности животному, достаю нитку с нервом. Прошу обратить внимание, что из металлической воронки, прижатой широким краем к брюшной стенке с отверстием панкреатического протока, не вытекает ни капли сока. Я начинаю теперь раздражать прерывистым электрическим током мой нерв. Как видите, животное стоит совершенно спокойно в станке, не обнаруживая и сейчас ни малейшим движением никакой боли. Проходит целых 2 минуты без всякого действия на железу (что и прошу вас особенно удержать в памяти), и только в третью появляется первая капля сока, за которою следуют дальнейшие, все чаще и чаще. После трех минут я заканчиваю раздражение, сок продолжает течь сам по себе и постепенно затихает только через 4-5 минут по окончании раздражения. Повторяя раздражение еще раз, как вы видите, мы получаем совершенно то же самое. И так всегда и на всех собаках. Нужно прибавить, что блуждающий нерв, конечно, раздражали и раньше с той же целью, что и мы, и, однако, не имели того, что теперь так легко показывается даже публично. Причина нашей удачи - некоторые особенности в постановке опыта. Этих особенностей две: животное во время опыта не подвергается никаким болезненным раздражениям, не говоря уже об отравлениях, большей частью сопровождающих обычную форму опытов; с другой стороны, при раздражении блуждающего нерва на шее, благодаря перерезке его за 4 дня до опыта, исключены резкие расстройства кровообращения. К четвертому дню перерезки замедляющие волокна сердца настолько потеряли в своей возбудимости вследствие развивающегося перерождения нерва, что самое сильное раздражение нерва едва дает себя знать весьма незначительным и мимолетным замедлением ударов сердца. Для понимания этой стороны опыта нужно не забывать, что возбудимость различных нервов после перерезки исчезает через различные сроки - и, следовательно, сейчас уас замедляющие волокна сердца потеряли свою возбудимость ранее, чем секреторные. Итак, в нашем опыте железы нисколько не пострадали ни от операции, ни от обстоятельств, сопутствующих раздражению.
Однако постоянного результата с раздражением блуждающего нерва можно достигнуть и в остром опыте, если только он ведется по некоторому определенному плану. Дело делается так. На нормальном животном быстро и возможно деликатно делается трахеотомия, затем в несколько секунд отделяется продолговатый мозг от спинного и вводится искусственное дыхание. Теперь можно спокойнее приступить к дальнейшему оперированию: вскрытию грудной полости для отпрепарования блуждающих нервов ниже сердца и вскрытию брюшной полости для вставления стеклянной трубочки в проток панкреатической железы. При этих условиях во всех опытах также наблюдается положительное секреторное действие блуждающего нерва на поджелудочную железу, хотя в начале опыта приходится частенько раздражать нерв несколько раз без видимого эффекта. Смысл указанных условий очевиден. Благодаря перерезке спинного мозга устраняется вредное рефлекторно-задерживающее действие дальнейшего длительного оперирования, a раздражение блуждающего нерва в грудной полости совершенно исключает изменения в ритмесердца.
Дальнейший анализ на этой второй форме опыта выдвинул два обстоятельства, от которых приощи нервов может произойти задерживание секреторной энергии поджелудочной железы. В наших опытах, как и в ранних опытах некоторых других авторов, панкреатическая железа оказалась резко чувствительной в отношении кровообращения. Достаточно кратковременного (2-3 минуты) раздражения сосудосуживающих ее нервов или зажатия аорты такой же продолжительности, чтобы железа перестала реагировать в продолжение некоторого периода времени на раздражения блуждающих нервов, ранее действительные. Эти опыты уже в значительной степени делают понятным, почему после обыкновенной операции, сопровождающейся сильнейшим чувствительным раздражением и, следовательно, рефлекторным сужением сосудов, железа животного, находящегося в разгаре пищеварения, сплошь и рядом не дает ни капли сока. Еще большее значение надо приписать другому фактору, который постоянно привлекал к себе внимание в наших опытах. Как в том опыте, который был проделат перед вами, так и в острых опытах раздражение блуждающего нерва не вызывает отделения сока моментально, а всегда проходит более или менее значительное время (от 15 сек. до нескольких минут) между моментом приложения раздражителя к нерву и наступлением отделительного эффекта. Сплошь и рядом бывает так, что сок начинает вытекать лишь в тот момент, когда прекращается раздражение. Наконец часто приходится наблюдать следующее особенное явление (д-р Метт). Вы, положим, долгое время раздражали правый блуждающий нерв и уже имеете от него равномерное отделение сока. Стоит в это время, не прерывая прежнего раздражения, присоединить раздражение другого блуждающего нерва для того, чтобы ток сока сейчас же остановился на известный, часто довольно значительный период времени.
Все эти явления вели к заключению, что в блуждающих нервах, вместе с возбуждающими элементами для pancreas, имеется и нечто ее тормозящее. Об этих тормозящих элементах можно сделать несколько предположений: они могут быть сосудосуживающими нервами поджелудочной железы, двигательными нервами мускулов ее каналов и, наконец, истинными секреторнозадерживающими нервами как антагонистами секреторных. Если при многих органах несомненно доказано существование антагонистической пары управляющих органом нервов, то почему же не быть этому и при железах? Может быть даже, этот антагонизм есть общий принцип всех иннерваций. Факты, намекающие на существование секреторнозадерживающих нервов желез, встречаются изредка в физиологической литературе последних лет. Но, мне кажется, вопрос об их существовании получит окончательное разрешение именно при исследовании иннервации желудка и поджелудочной железы, так как здесь задерживающие явления выражены наиболее резко. Ранее подробного анализа этого вопроса, я приведу опыты, относящиеся до секреторного действия на поджелудочную железу симпатического нерва, так как они доставят с своей стороны некоторый материал для обсуждения интересующего нас вопроса. Вот результаты проф. Кудрявицкого.
Если в описанной выше острой форме опыта с нашей железой раздражать обыкновенным прерывистым индукционным током симпатический нерв, то только в самый первый момент раздражения замечается маленькое толчкообразное движение сока, а затем во все время раздражения и после него - ни малейшего отделения. Применяя же механическое раздражение (ряд толчков посредством тетаномотора Гейденгайна) вместо электрического, часто видят другой результат: спустя некоторое время после начала раздражения происходит довольно сильное отделение сока. Можно того же достигнуть и электрическим раздражением, если раздражать нерв не свежий, а перерезанный за 4-5 дней до этого и, следовательно, подвергающийся перерождению. Смысл этих явлений легко понять, если припомнить некоторые пункты из физиологии сосудодвигательных нервов. Известно, что эти нервы мало чувствительны к механическому раздражению, а после их перерезки ранее многих других волокон теряют свою возбудимость. Следовательно, мы имеем право принимать: 1) что в симпатическом нерве одновременно находятся как суживающие, так и отделительные нервы поджелудочной железы, 2) что при обыкновенном электрическом раздражении свежего нерва сосудосуживающие волокна вполне маскируют отделительные и 3) что лишь при особенных условиях (механическое раздражение и электрическое раздражение заранее перерезанного нерва), устраняющих вмешательства сосудосуживающих волокон, отделительные волокна получают возможность заявить о своем существовании.
Таким образом симпатический нерв представил нам удобный случай установить взаимное отношение сосудодвигательных и секреторных нервов поджелудочной железы. Однако применение указанных приемов при раздражении блуждающего нерва нисколько не изменило картины его действия на нашу железу: задерживающее действие блуждающего нерва осталось при этом в полной целости. И это дает хорошее основание думать, что тормозящее действие блуждающего нерва не обусловливается сужением кровеносных сосудов, самое последнее время д-р Попельский сильно подвинулся в решении занимающего нас вопроса. Им прежде всего была выработана форма опыта, при которой задерживающее по отношению к поджелудочной железе действие блуждающего нерва выступает постоянно и притом в очень резком виде. На острой форме опыта (как описано раньше) вливается в двенадцатиперстную кишку раствор соляной кислоты. Этим вызывается продолжительное и обильное отделение панкреатического сока. Сильное раздражение в это время блуждающих нервов всякий раз без исключения сейчас же обусловливает замедление, а чаще совершенное прекращение отделения. Раздражение же симпатического нерва только замедляет отделение и притом лишь спустя некоторое время после начала раздражения. Точно так же зажатие аорты останавливает отделение только через минуту, две, три. При этом нельзя не упомянуть еще, что, по последним опытам Франсуа-Франка, блуждающий нерв скорее расширяет сосуды поджелудочной железы, чем сужает их. Возможность вмешательства двигательных нервов протоков железы исключалась тем, что животное отравлялось физостигмином, сильнейшим раздражителем гладкой мускулатуры, причем, однако, не только не оказалось никакого задерживания сока, а скорее ядом было усилено отделение. Наконец при подробном препаровании нервов железы удалось найти такие ветви, которые при раздражении вызывали отделение без длинного латентного периода, почти так же быстро, как барабанная струна гонит слюну. Из последнего опыта нужно заключить, что в данном пункте секреторные волокна поджелудочной железы анатомически отделились от задерживающих элементов и что, следовательно, чистым секреторным нервам не принадлежит особенность обусловливать деятельность органа при искусственном раздражении лишь после продолжительного периода скрытого действия. Д-р Попельский нашел, наконец, в отдельности и такие ветви блуждающего нерва, которые только задерживали, но никогда не возбуждали отделения поджелудочного сока. Конечно, раз существуют такие специально задерживающие нервы, то вполне возможны и рефлекторные раздражения их как при нормальных условиях, так и при оперировании. Не исключается, наконец, возможность рефлекторного задерживания центров самих секреторных нервов pancreas.
Из приведенных наблюдений точно и фактически объясняются все неудачи и трудности прежних исследований над иннервациею поджелудочной железы. Например, почему Гейденгайн при раздражении продолговатого мозга получил положительный результат только в немногих опытах? Не говоря о задерживающем влиянии операции, раздражая мозг, он вызывал и сильное сужение сосудов и нарушение деятельности сердца, а к тому же оставалось в силе и раздражение антагонистических волокон.
Вы уже, конечно, заметили, до чего сходно идут нервные явления на желудочных и панкреатических железах: во всех стношениях иннервация одних есть копия других. Вот почему недостающее в одной иннервации против другой можно с правом восполнить по аналогии. На этом основании мы не можем сомневаться, например, в том, что секреторные волокна желез, помимо блуждающих нервов, находятся и в симпатическом нерве.
В заключение несколько слов о вышеупомянутом опыте двух французских авторов над желудком обезглавленного преступника. После знакомства с фактами крайней щепетильности наших желез нелегко поверить выводу авторов, что они имели перед собою истинный отделительный эффект блуждающего нерва 40 минут спустя после обескровления органа.
Мне кажется, я могу верить, что после всего сообщенного и показанного вам представляются так же бесспорными и действительными отделительные нервы желудочных и поджелудочной желез, как классическая, всем известная chorda tympani при слюнных железах. Само собой разумеется, что, кроме этих специальных нервов, в наши железы входят и сосудистые нервы, сосудосуживающие и сосудорасширяющие.

 Ясно, что поддразнивание не только не уступает мнимому кормлению, а скорее в данном случае превосходит его по сокогонному эффекту.
Итак, факт Биддера и Шмидта вполне правилен, но нельзя сказать, чтобы он был общепризнан и вполне оценен в физиологии. Известно несколько авторов, которые не могли убедиться в нем, и многие учебники физиологии не считают надобным упоминать о нем. В настоящее время мы можем отдать себе полный отчет о судьбе факта в руках различных исследователей. Этот факт может обнаруживаться только при определенных условиях. Вопервых, для удачи опыта, конечно, требуется нормальное состояние животного как в отношении самочувствия, так и полной неприкосновенности слизистой оболочки желудка, что у многих авторов, получивших отрицательный результат, судя по их описаниям, не всегда бывало. Во-вторых, успех опыта определяет, как сказано выше, степень желания есть, она зависит от того, как много и как давно перед этим собака ела и чем ее дразнят: действительно ли ей интересным блюдом или таким, к которому она относится равнодушно. Известно, что собаки имеют столь же различные вкусы, как и люди. В-третьих, и между собаками встречаются более положительные и хладнокровные типы, которые не имеют привычки дразниться мечтой, тем, что далеко от рта, а терпеливо и спокойно ждут, когда пища окажется у них во рту. Следовательно, для опыта нужны более жадные и более мечтательные животные. Наконец, в-четвертых, чрезвычайно важный момент, с которым надо считаться в этих опытах, это - догадливость и обидчивость животных. Довольно часто попадаются собаки, которые скоро замечают, что их дразнят едой, и сердятся на это, упорно отворачиваясь от всего того, что вы проделываете перед ними. Поэтому всегда лучше ставить опыт с поддразниванием так, как будто вы и не думаете дразнить животное, а просто собираетесь к его корму. При внимании к перечисленным условиям опыт с психическим отделением желудочного сока, как его обыкновенно называют, делается таким же постоянным, как и опыт с мнимым кормлением. При долгом занятии работою желудочных желез при различных условиях проникаешься убеждением, до чего опасным для всех опытов является факт психического отделения сока. Вы должны постоянно, так сказать, вести борьбу с этим фактором, постоянно считаться с ним, постоянно обеспечивать себя против него. Если собака долго не ела, то каждое ваше движение, каждый ваш выход из комнаты, каждое появление служителя, который ее кормит, и т. п., - все это может быть иногда толчком к работе желез. Самое неустанное и тщательное внимание требуется для того, чтобы избежать этого источника ошибок, и едва ли мы ошибемся, если скажем, что немалое в прежнем материале, относящемся до работы желудочных желез, было приписано тем или другим условиям опыта, когда на самом деле определялось просматриваемым психическим моментом. Поэтому мы во многих опытах, ради полной бесспорности заключения о значении того или другого условия отделения, старались псльзоваться спящим животным, так как на массе опытов убедились, что сон не имеет заметного задерживающего влияния на работу желудочных желез.
Итак, имея в виду бесплодность попыток каким-нибудь раздражением полости рта вызвать отделение желудочного сока, а с другой стороны, вполне убедившись в действительности, постоянстве и силе, при определенных условиях, психического момента как раздражителя секреторных нервов желудка, мы приходим к окончательному заключению, что при нашем опыте с мнимым кормлением весь отделительный эффект определяется только психическим моментом, т. е. страстным желанием еды и наслаждением ею. Ввиду важного значения акта еды, счевидного уже прямо (а при исследовании дальнейшего периода отделительной деятельности желудка оно окажется еще больше), мы не жалели ни времени, ни труда, чтобы вполне бесспорно установить механизм интересующего нас факта. Мы проделали ради этого массу видоизменений опыта с мнимым кормлением, которые все только усиливали наше заключение об его натурс. Если вы подготовите животное продолжительным голоданием (в продолжение двухтрех дней), то, что бы вы ему теперь ни дали для еды (мясо вареное, сырое, хлеб, вареный белок и т. д.), на все в ответ получится весьма обильное отделение желудочного сока; между тем как собака, не голодавшая (т. е. часов через 15- 20 после последней еды), будет резко различать между перечисленными сортами еды, может одни есть с большою жадностью, другие вяло, а то и совсем не есть, и соответственно с этим также резко будут колебаться и количество и качество отделяемого сока. Чем жаднее собака ест, тем сока выделяется больше и с гораздо большею переваривающею способностью. Большинство собак предпочитает мясо хлебу, и, в согласии с этим, обыкновенно при мнимом кормлении хлебом сока выделяется меньше и более слабого по пищеварительной силе. Но попадаются собаки лучше, с большим аппетитом накидывающиеся на хлеб, чем на мясо, и у таких собак при мнимом кормлении хлебом, вопреки правилу, получается и больше сока и более сильного. Приведем еще такой случай. Вы даете известной собаке вареное мяс известною, определенною частотою, кусками определенной величины. Собака ест, но уже по общему поведению собаки вы замечаете, что она особенной жадности к этой еде не обнаруживает, что и доказывается вполне тем, что через 15 20 минут она перестает брать от вас куски. Вместе с тем отделение сока или совсем не начинается при этом, или, начавшись с опозданием против 5 минут, останется до конца еды незначительным. Той же собаке, выждав, когда затихнет предшествующее отделение, или в ближайший день, вы даете такими же кусками и с тою же частотою, как и раньше, сырое мясо, которое ей, очевидно, сильно по вкусу, потому что она будет его есть часами, и теперь отделение сока начнется ровно через 5 минут и будет обильно. У другой же собаки, предпочитающей вареное мясо сырому, все окажется наоборот. Бульон, суп, молоко, к которым собаки по правилу всегда равнодушнее, чем к твердой пище, часто или совсем не возбуждают отделения, или слабое, хотя бульон воспроизводит все вкусовые свойства мяса.
Совершенно ясно, что психический момент, работая при мнимой еде, легко приобретает абсолютно постоянный характер. Все условия, которые перечислены выше как необходимые для успеха опыта с психическим возбуждением желудочного сока, при мнимом кормлении - налицо и соединяются друг с другом; животное с жадностью на наших глазах ест, следовательно, то, что ест, ему по вкусу; ест на самом деле, а не воображает только о еде, и, конечно, никаких поводов к обиде не имеется, так как ни одно животное, конечно, не догадывается тщете того дела, которым его занимают.
Итак, при акте еды, при нашем мнимом кормлении, раздражителем железистых нервов желудка является психический момент, приобревший физиологический характер, т. е. сделавшийся обязательным, непременно повторяющимся при определенном условии, как любое, вполне изученное физиологическое явление. Смотря на все явления только с чисто физиологической стороны, можно сказать, что это сложный рефлекс. Его сложность понятна, потому что физиологическая цель в данном случае может быть достигнута лишь целым рядом деятельностей организма. Объект пищеварения -- пища - находится вне тела, во внешнем мире, она должна быть доставлена в организм не только при помощи мышечной силы, но и высших отправлений организма - смысла, воли и желания животного. Соответственно этому одновременное раздражение пищей различных органов чувств: зрения, слуха, обоняния и Вкуса, в особенности последних, так как деятельность их связана с нахождением пищи поблизости или уже в сфере организма, является вернейшим и сильнейшим ударом по секреторным нервам желез. Страстным инстинктом еды настойчивая и неустанная природа тесно связала искание, добывание еды с началом ее обработки в организме. Нетрудно догадаться, что столь подробно анализированный нами факт находится в тесной связи с повседневным явлением людской жизни - аппетитом. Этот деятель, столь важный в жизни и вместе остававшийся таинственным для науки, облекается, наконец, в научную плоть и кровь, превращается из субъективного ощущения точный лабораторный факт.
Итак, мы считаем себя вправе сказать, что аппетит есть первый и сильнейший раздражитель секреторных нервов желудочных желез, есть то, что при мнимом кормлении наших собак обусловливает истечение из совершенно пустого желудка многих сотен куб. сантиметров энергичнеишего желудочного сока. Сильный аппетит при еде - значит обильное отделение с самого начала еды сильного сока; нет аппетита, нет и этого начального сока; возвратить аппетит человеку - значит дать ему большую порцию хорошего сока в начале еды.
Ясно, что поддразнивание не только не уступает мнимому кормлению, а скорее в данном случае превосходит его по сокогонному эффекту.
Итак, факт Биддера и Шмидта вполне правилен, но нельзя сказать, чтобы он был общепризнан и вполне оценен в физиологии. Известно несколько авторов, которые не могли убедиться в нем, и многие учебники физиологии не считают надобным упоминать о нем. В настоящее время мы можем отдать себе полный отчет о судьбе факта в руках различных исследователей. Этот факт может обнаруживаться только при определенных условиях. Вопервых, для удачи опыта, конечно, требуется нормальное состояние животного как в отношении самочувствия, так и полной неприкосновенности слизистой оболочки желудка, что у многих авторов, получивших отрицательный результат, судя по их описаниям, не всегда бывало. Во-вторых, успех опыта определяет, как сказано выше, степень желания есть, она зависит от того, как много и как давно перед этим собака ела и чем ее дразнят: действительно ли ей интересным блюдом или таким, к которому она относится равнодушно. Известно, что собаки имеют столь же различные вкусы, как и люди. В-третьих, и между собаками встречаются более положительные и хладнокровные типы, которые не имеют привычки дразниться мечтой, тем, что далеко от рта, а терпеливо и спокойно ждут, когда пища окажется у них во рту. Следовательно, для опыта нужны более жадные и более мечтательные животные. Наконец, в-четвертых, чрезвычайно важный момент, с которым надо считаться в этих опытах, это - догадливость и обидчивость животных. Довольно часто попадаются собаки, которые скоро замечают, что их дразнят едой, и сердятся на это, упорно отворачиваясь от всего того, что вы проделываете перед ними. Поэтому всегда лучше ставить опыт с поддразниванием так, как будто вы и не думаете дразнить животное, а просто собираетесь к его корму. При внимании к перечисленным условиям опыт с психическим отделением желудочного сока, как его обыкновенно называют, делается таким же постоянным, как и опыт с мнимым кормлением. При долгом занятии работою желудочных желез при различных условиях проникаешься убеждением, до чего опасным для всех опытов является факт психического отделения сока. Вы должны постоянно, так сказать, вести борьбу с этим фактором, постоянно считаться с ним, постоянно обеспечивать себя против него. Если собака долго не ела, то каждое ваше движение, каждый ваш выход из комнаты, каждое появление служителя, который ее кормит, и т. п., - все это может быть иногда толчком к работе желез. Самое неустанное и тщательное внимание требуется для того, чтобы избежать этого источника ошибок, и едва ли мы ошибемся, если скажем, что немалое в прежнем материале, относящемся до работы желудочных желез, было приписано тем или другим условиям опыта, когда на самом деле определялось просматриваемым психическим моментом. Поэтому мы во многих опытах, ради полной бесспорности заключения о значении того или другого условия отделения, старались псльзоваться спящим животным, так как на массе опытов убедились, что сон не имеет заметного задерживающего влияния на работу желудочных желез.
Итак, имея в виду бесплодность попыток каким-нибудь раздражением полости рта вызвать отделение желудочного сока, а с другой стороны, вполне убедившись в действительности, постоянстве и силе, при определенных условиях, психического момента как раздражителя секреторных нервов желудка, мы приходим к окончательному заключению, что при нашем опыте с мнимым кормлением весь отделительный эффект определяется только психическим моментом, т. е. страстным желанием еды и наслаждением ею. Ввиду важного значения акта еды, счевидного уже прямо (а при исследовании дальнейшего периода отделительной деятельности желудка оно окажется еще больше), мы не жалели ни времени, ни труда, чтобы вполне бесспорно установить механизм интересующего нас факта. Мы проделали ради этого массу видоизменений опыта с мнимым кормлением, которые все только усиливали наше заключение об его натурс. Если вы подготовите животное продолжительным голоданием (в продолжение двухтрех дней), то, что бы вы ему теперь ни дали для еды (мясо вареное, сырое, хлеб, вареный белок и т. д.), на все в ответ получится весьма обильное отделение желудочного сока; между тем как собака, не голодавшая (т. е. часов через 15- 20 после последней еды), будет резко различать между перечисленными сортами еды, может одни есть с большою жадностью, другие вяло, а то и совсем не есть, и соответственно с этим также резко будут колебаться и количество и качество отделяемого сока. Чем жаднее собака ест, тем сока выделяется больше и с гораздо большею переваривающею способностью. Большинство собак предпочитает мясо хлебу, и, в согласии с этим, обыкновенно при мнимом кормлении хлебом сока выделяется меньше и более слабого по пищеварительной силе. Но попадаются собаки лучше, с большим аппетитом накидывающиеся на хлеб, чем на мясо, и у таких собак при мнимом кормлении хлебом, вопреки правилу, получается и больше сока и более сильного. Приведем еще такой случай. Вы даете известной собаке вареное мяс известною, определенною частотою, кусками определенной величины. Собака ест, но уже по общему поведению собаки вы замечаете, что она особенной жадности к этой еде не обнаруживает, что и доказывается вполне тем, что через 15 20 минут она перестает брать от вас куски. Вместе с тем отделение сока или совсем не начинается при этом, или, начавшись с опозданием против 5 минут, останется до конца еды незначительным. Той же собаке, выждав, когда затихнет предшествующее отделение, или в ближайший день, вы даете такими же кусками и с тою же частотою, как и раньше, сырое мясо, которое ей, очевидно, сильно по вкусу, потому что она будет его есть часами, и теперь отделение сока начнется ровно через 5 минут и будет обильно. У другой же собаки, предпочитающей вареное мясо сырому, все окажется наоборот. Бульон, суп, молоко, к которым собаки по правилу всегда равнодушнее, чем к твердой пище, часто или совсем не возбуждают отделения, или слабое, хотя бульон воспроизводит все вкусовые свойства мяса.
Совершенно ясно, что психический момент, работая при мнимой еде, легко приобретает абсолютно постоянный характер. Все условия, которые перечислены выше как необходимые для успеха опыта с психическим возбуждением желудочного сока, при мнимом кормлении - налицо и соединяются друг с другом; животное с жадностью на наших глазах ест, следовательно, то, что ест, ему по вкусу; ест на самом деле, а не воображает только о еде, и, конечно, никаких поводов к обиде не имеется, так как ни одно животное, конечно, не догадывается тщете того дела, которым его занимают.
Итак, при акте еды, при нашем мнимом кормлении, раздражителем железистых нервов желудка является психический момент, приобревший физиологический характер, т. е. сделавшийся обязательным, непременно повторяющимся при определенном условии, как любое, вполне изученное физиологическое явление. Смотря на все явления только с чисто физиологической стороны, можно сказать, что это сложный рефлекс. Его сложность понятна, потому что физиологическая цель в данном случае может быть достигнута лишь целым рядом деятельностей организма. Объект пищеварения -- пища - находится вне тела, во внешнем мире, она должна быть доставлена в организм не только при помощи мышечной силы, но и высших отправлений организма - смысла, воли и желания животного. Соответственно этому одновременное раздражение пищей различных органов чувств: зрения, слуха, обоняния и Вкуса, в особенности последних, так как деятельность их связана с нахождением пищи поблизости или уже в сфере организма, является вернейшим и сильнейшим ударом по секреторным нервам желез. Страстным инстинктом еды настойчивая и неустанная природа тесно связала искание, добывание еды с началом ее обработки в организме. Нетрудно догадаться, что столь подробно анализированный нами факт находится в тесной связи с повседневным явлением людской жизни - аппетитом. Этот деятель, столь важный в жизни и вместе остававшийся таинственным для науки, облекается, наконец, в научную плоть и кровь, превращается из субъективного ощущения точный лабораторный факт.
Итак, мы считаем себя вправе сказать, что аппетит есть первый и сильнейший раздражитель секреторных нервов желудочных желез, есть то, что при мнимом кормлении наших собак обусловливает истечение из совершенно пустого желудка многих сотен куб. сантиметров энергичнеишего желудочного сока. Сильный аппетит при еде - значит обильное отделение с самого начала еды сильного сока; нет аппетита, нет и этого начального сока; возвратить аппетит человеку - значит дать ему большую порцию хорошего сока в начале еды.
 Вы видите в обоих случаях тождественное начало как в отнонении количества, так и в отношении переваривающей силы, и только затем, во втором часу, идет обособление отделительной работы по роду пищи. Что же это за начало? Не то ли, что мы видели при мнимом кормлении? Не струя ли начального психического сока в общем потоке отделения? Бесспорно, господа, это действительно так, и мы можем убедиться в том разнообразными способами. Прежде всего ясно само по себе: то, что имело место при нашем так называемом мнимом кормлении, не могло же почему-то исчезнуть при нормальной еде; ведь опыт мнимого кормления есть уединение, как бы отрезанное начало нормального акта пищеварения. Это законное рассуждение подтверждается уже одним сравнением отделения первого времени после еды мяса и хлеба с отделением при мнимом кормлении. При мясе, как и хлебе, бросается в глаза одинаковая и значительная переваривающая сила первого часа, и эта сила совпадает с наиболее частой силой сока при мнимом кормлении. Точно так и величина отделения первого часа в нашем уединенном желудочке, рассчитанная на весь желудок (для этого надо помножить ее на 10, так как уединенный желудочек составляет около десятой части всего желудка), падает в категорию тех количеств, которые обыкновенно получаются при мнимом кормлении. Наконец и самый ход обеих величин, переваривающей силы и количества, а именно, изменение их вскоре после акта еды (в мясе - переваривающей силы, в хлебе количества), отчетливо указывает на их связь с актом еды, с тем, что существует лишь временно и затем постепенно изглаживается, заменяясь другими условиями. Убедительноеть приведенного рассуждения усиливается рассмотрением дальнейшего случая еды. Дайте собаке съесть что-нибудь, не так ее интересующее, как мясо и хлеб, и вы не увидите этого взмаха в количестве и силе сока. Предложите, например, собаке молока, мнимое кормление коим довольно часто не сопровождается сколько-нибудь значительным отделением сока, - ни следа этого начального сильного отделения, этого взмаха. Вы уже видели эти цифры, но я считаю полезным показать их еще раз для сравнение с отделением при мясе и хлебе.
Дано съесть 600 куб. см молока (опыт доктора Хижина).
Вы видите в обоих случаях тождественное начало как в отнонении количества, так и в отношении переваривающей силы, и только затем, во втором часу, идет обособление отделительной работы по роду пищи. Что же это за начало? Не то ли, что мы видели при мнимом кормлении? Не струя ли начального психического сока в общем потоке отделения? Бесспорно, господа, это действительно так, и мы можем убедиться в том разнообразными способами. Прежде всего ясно само по себе: то, что имело место при нашем так называемом мнимом кормлении, не могло же почему-то исчезнуть при нормальной еде; ведь опыт мнимого кормления есть уединение, как бы отрезанное начало нормального акта пищеварения. Это законное рассуждение подтверждается уже одним сравнением отделения первого времени после еды мяса и хлеба с отделением при мнимом кормлении. При мясе, как и хлебе, бросается в глаза одинаковая и значительная переваривающая сила первого часа, и эта сила совпадает с наиболее частой силой сока при мнимом кормлении. Точно так и величина отделения первого часа в нашем уединенном желудочке, рассчитанная на весь желудок (для этого надо помножить ее на 10, так как уединенный желудочек составляет около десятой части всего желудка), падает в категорию тех количеств, которые обыкновенно получаются при мнимом кормлении. Наконец и самый ход обеих величин, переваривающей силы и количества, а именно, изменение их вскоре после акта еды (в мясе - переваривающей силы, в хлебе количества), отчетливо указывает на их связь с актом еды, с тем, что существует лишь временно и затем постепенно изглаживается, заменяясь другими условиями. Убедительноеть приведенного рассуждения усиливается рассмотрением дальнейшего случая еды. Дайте собаке съесть что-нибудь, не так ее интересующее, как мясо и хлеб, и вы не увидите этого взмаха в количестве и силе сока. Предложите, например, собаке молока, мнимое кормление коим довольно часто не сопровождается сколько-нибудь значительным отделением сока, - ни следа этого начального сильного отделения, этого взмаха. Вы уже видели эти цифры, но я считаю полезным показать их еще раз для сравнение с отделением при мясе и хлебе.
Дано съесть 600 куб. см молока (опыт доктора Хижина).
 Вот вам и начало анализа различных пунктов наших кривых отделения.
Ввиду важности предмета изучение не ограничилось выводами из ранних опытов; было поставлено несколько новых форм опытов.
Мы разделили обыкновенную мясную порцию нашей собаки - 400 г - на четыре части и давали их последовательно через каждые полтора часа (опыты приват-доцента Котляра и доктора Лобасова). Всякий раз, после дачи 100 г мяса мы получаем взмах количества и силы сока. Представляю таблицу (8) чисел.
В кривой (рис. 10) воспроизводится только колебание переваривающей силы сока как самого характерного свойства его.
Вот вам и начало анализа различных пунктов наших кривых отделения.
Ввиду важности предмета изучение не ограничилось выводами из ранних опытов; было поставлено несколько новых форм опытов.
Мы разделили обыкновенную мясную порцию нашей собаки - 400 г - на четыре части и давали их последовательно через каждые полтора часа (опыты приват-доцента Котляра и доктора Лобасова). Всякий раз, после дачи 100 г мяса мы получаем взмах количества и силы сока. Представляю таблицу (8) чисел.
В кривой (рис. 10) воспроизводится только колебание переваривающей силы сока как самого характерного свойства его.
 Ясно, что и высокая переваривающая сила и большое количество сока связаны именно с актом еды.
Ясно, что и высокая переваривающая сила и большое количество сока связаны именно с актом еды.
 Представлялось интересным у нашей собаки с уединенным желудочком определить непосредственно размер и качество того отделения, которое получается при акте еды. С этою целью мы вначале до известной степени только исходит у собак с перерезанным пищеводом. У собаки, кроме уединенного желудочка, имелась и обыкновенная фистула в главном желудке. Открыв эту фистулу и давая собаке есть обычным порядком нарезанное небольшими кусочками мясо, мы получаем их сейчас же назад из желудочной фистулы покрытые слюной. Совершенно, как при мнимом кормлении, не раньше 5 минут, начинает вытекать сок как из большого, так и из уединенного желудочков, причем истечение это происходит достаточно правильно в обеих полостях; точно так же совпадает и конец отделения там и сям по прекращении кормления. Представляю пример таких опытов (из работы доктора Лобасова).
В продолжение пяти минут собака съела 80 кусков мяса (172 г весом), которые скоро все вывалились из желудочной фистулы. Отделение из обоих желудков началось одновременно на 7-й минуте от начала кормлени продолжалось следующим образом (табл. 9).
Представлялось интересным у нашей собаки с уединенным желудочком определить непосредственно размер и качество того отделения, которое получается при акте еды. С этою целью мы вначале до известной степени только исходит у собак с перерезанным пищеводом. У собаки, кроме уединенного желудочка, имелась и обыкновенная фистула в главном желудке. Открыв эту фистулу и давая собаке есть обычным порядком нарезанное небольшими кусочками мясо, мы получаем их сейчас же назад из желудочной фистулы покрытые слюной. Совершенно, как при мнимом кормлении, не раньше 5 минут, начинает вытекать сок как из большого, так и из уединенного желудочков, причем истечение это происходит достаточно правильно в обеих полостях; точно так же совпадает и конец отделения там и сям по прекращении кормления. Представляю пример таких опытов (из работы доктора Лобасова).
В продолжение пяти минут собака съела 80 кусков мяса (172 г весом), которые скоро все вывалились из желудочной фистулы. Отделение из обоих желудков началось одновременно на 7-й минуте от начала кормлени продолжалось следующим образом (табл. 9).
 Отделение кончилось в обеих полостях в одно и то же время.
Этот опыт, во-первых, убеждает в параллельности работы большого и маленького желудков: моменты начала и конца и колебания в промежуточных стадиях совершенно совпадают. Вовторых, переваривающая сила вытекающего из обеих полостей сока также достаточно одинакова и вполне сходна с той, которая обыкновенно наблюдается при так называемом мнимом кормлении. Она осталась теперь такою же до последней капли, не изменившись в те меньшие величины, которые обыкновенно наблюдаются при отделении после нормального кормления мясом, начиная со второго часа.
То же самое оказалось и тогда, когда нашей собаке была сделана впоследствии операция эзофаготомии, и опыт с мнимым кормлением мог быть произведен на ней в своей типичной форме. Вот один из таких опытов (из работы д-ра Лобасова).
Первая капля показалась одновременно в обеих полостях на 6-й минуте от начала мнимого кормления, затем отделение продолжалось следующим образом при получасовом кормлении (табл. 10).
Отделение кончилось в обеих полостях в одно и то же время.
Этот опыт, во-первых, убеждает в параллельности работы большого и маленького желудков: моменты начала и конца и колебания в промежуточных стадиях совершенно совпадают. Вовторых, переваривающая сила вытекающего из обеих полостей сока также достаточно одинакова и вполне сходна с той, которая обыкновенно наблюдается при так называемом мнимом кормлении. Она осталась теперь такою же до последней капли, не изменившись в те меньшие величины, которые обыкновенно наблюдаются при отделении после нормального кормления мясом, начиная со второго часа.
То же самое оказалось и тогда, когда нашей собаке была сделана впоследствии операция эзофаготомии, и опыт с мнимым кормлением мог быть произведен на ней в своей типичной форме. Вот один из таких опытов (из работы д-ра Лобасова).
Первая капля показалась одновременно в обеих полостях на 6-й минуте от начала мнимого кормления, затем отделение продолжалось следующим образом при получасовом кормлении (табл. 10).
 Отделение кончилось одновременно в обеих полостях.
Представляю то же в виде кривых, причем масштаб для количеств сока, вытекающего из большого желудка, взят в 10 раз меньше [рис. 11, 12].
Как видите, ход отделения в обоих случаях тождественный.
Желудочная фистула в большом желудке нашей собаки дает нам возможность поставить опыт, совершенно обратный опыту с мнимым кормлением, истинный experimentum crucis, перекрестный опыт. Если в опыте с мнимым кормлением существует только одно начало пищеварительного акта, то в перекрестном опыте можно начать прямо, так сказать, с продолжения этого акта: стоит только незаметно для собаки вложить ту или другую пищу через фистульную трубку в большой желудок. Так как при этих опытах является существенным сделать это, не возбудив аппетита собаки, то всего проще поставить такой опыт на заснувшем животном. Однако сейчас же спешу заявить, что того же можно достигнуть и на бодром животном, если сделать это незаметно для животного, всячески отвлекая его мысли от еды.
Результаты опытов поразительны. Ничего подобного тому, что мы видели после обычной еды! Некоторые сорта еды, например хлеб и свернутый яичный белок, при введении прямо в желудок, в первый час и дальше не дают совершенно ни одной капли сока. Это касается как маленького, так и большого желудка; в последнем легко убедиться погружением стеклянной палочки в пищевую массу, находящуюся в большом желудке, палочка остается сухой. Мясо вызывает отделение и при вкладывании, но резко запаздывающее (начало отделения теперь 15 45 минут вместо 6 10 минут, как при еде), очень незначительное по размеру в первый час (3-5 куб. см вместо 11-12 куб. см, как при еде) и очень низкую переваривающею силою. Показываю таблицу опыта (д-ра Лобасова) (табл. 11).
Отделение сока началось 25 минут спустя после вкладывания.
Отделение кончилось одновременно в обеих полостях.
Представляю то же в виде кривых, причем масштаб для количеств сока, вытекающего из большого желудка, взят в 10 раз меньше [рис. 11, 12].
Как видите, ход отделения в обоих случаях тождественный.
Желудочная фистула в большом желудке нашей собаки дает нам возможность поставить опыт, совершенно обратный опыту с мнимым кормлением, истинный experimentum crucis, перекрестный опыт. Если в опыте с мнимым кормлением существует только одно начало пищеварительного акта, то в перекрестном опыте можно начать прямо, так сказать, с продолжения этого акта: стоит только незаметно для собаки вложить ту или другую пищу через фистульную трубку в большой желудок. Так как при этих опытах является существенным сделать это, не возбудив аппетита собаки, то всего проще поставить такой опыт на заснувшем животном. Однако сейчас же спешу заявить, что того же можно достигнуть и на бодром животном, если сделать это незаметно для животного, всячески отвлекая его мысли от еды.
Результаты опытов поразительны. Ничего подобного тому, что мы видели после обычной еды! Некоторые сорта еды, например хлеб и свернутый яичный белок, при введении прямо в желудок, в первый час и дальше не дают совершенно ни одной капли сока. Это касается как маленького, так и большого желудка; в последнем легко убедиться погружением стеклянной палочки в пищевую массу, находящуюся в большом желудке, палочка остается сухой. Мясо вызывает отделение и при вкладывании, но резко запаздывающее (начало отделения теперь 15 45 минут вместо 6 10 минут, как при еде), очень незначительное по размеру в первый час (3-5 куб. см вместо 11-12 куб. см, как при еде) и очень низкую переваривающею силою. Показываю таблицу опыта (д-ра Лобасова) (табл. 11).
Отделение сока началось 25 минут спустя после вкладывания.
 Теперь прошу сопоставить следующую таблицу (табл. 12).
Теперь прошу сопоставить следующую таблицу (табл. 12).
 To То же самое воспроизвожу на кривых. Рис. 13 -16 представляют ход отделения сока. Как видите, кривая при вкладывании мяса и гораздо медленнее поднимается и далеко не достигает высоты кривой при еде мяса, но если ее сложить по частям с кривой опыта мнимого кормления, то вы получите почти тождественную кривую с кривой при еде.
Точно такой же расчет с успехом может быть применен и к переваривающей силе соков вышеприведенных опытов. Пример синтеза кривой из ее элементов!
Наконец я в состоянии провести перед вами следующий поучительный опыт. В присутствии некоторых из слушателей, пришедших по уговору на лекцию часом раньше, я распорядился с двумя собаками, имеющими обыкновенные желудочные фистулы и перерезанный на шее пищевод, следующим образом. Одной из них, по возможности незаметно для нее, т. е. отвлекая ее ласками и принимая меры против раздражения обонятельных нервов собаки, введено через желудочную фистулу несколько десятков кусков сырого мяса, навязанных в виде четок на нитку, конец которой ущемлен в желудочной фистуле пробкой. Собака дальше предоставляется самой себе в отдельной комнате. С другой собакой, которой так же и столько же вложено мяса в желудок, одновременно с вкладыванием проделывается процедура мнимого кормления, затем животное также предоставляется себе. Обеим собакам вложено по 100 г мяса. Теперь прошло уже полтора часа после вкладывания; вынимая за нитки у обеих собак введенное мясо назад и взвешивая его, мы видим большую разницу в переваривании мяса у той и другой. то время как у собаки без мнимого кормления вес уменьшился лиць на 6 г, вес мяса у собаки с мнимым кормлением равняется всего 70 г, т. е. переварилось 30 г. Вот вам пищеварительная цена акта прохождения пищи через рот, цена страстного желания, сопровождающего еду, цена аппетита! Представляю, кроме того, ряд чисел, добытых при такой постановке опытов доктором Лобановым.
To То же самое воспроизвожу на кривых. Рис. 13 -16 представляют ход отделения сока. Как видите, кривая при вкладывании мяса и гораздо медленнее поднимается и далеко не достигает высоты кривой при еде мяса, но если ее сложить по частям с кривой опыта мнимого кормления, то вы получите почти тождественную кривую с кривой при еде.
Точно такой же расчет с успехом может быть применен и к переваривающей силе соков вышеприведенных опытов. Пример синтеза кривой из ее элементов!
Наконец я в состоянии провести перед вами следующий поучительный опыт. В присутствии некоторых из слушателей, пришедших по уговору на лекцию часом раньше, я распорядился с двумя собаками, имеющими обыкновенные желудочные фистулы и перерезанный на шее пищевод, следующим образом. Одной из них, по возможности незаметно для нее, т. е. отвлекая ее ласками и принимая меры против раздражения обонятельных нервов собаки, введено через желудочную фистулу несколько десятков кусков сырого мяса, навязанных в виде четок на нитку, конец которой ущемлен в желудочной фистуле пробкой. Собака дальше предоставляется самой себе в отдельной комнате. С другой собакой, которой так же и столько же вложено мяса в желудок, одновременно с вкладыванием проделывается процедура мнимого кормления, затем животное также предоставляется себе. Обеим собакам вложено по 100 г мяса. Теперь прошло уже полтора часа после вкладывания; вынимая за нитки у обеих собак введенное мясо назад и взвешивая его, мы видим большую разницу в переваривании мяса у той и другой. то время как у собаки без мнимого кормления вес уменьшился лиць на 6 г, вес мяса у собаки с мнимым кормлением равняется всего 70 г, т. е. переварилось 30 г. Вот вам пищеварительная цена акта прохождения пищи через рот, цена страстного желания, сопровождающего еду, цена аппетита! Представляю, кроме того, ряд чисел, добытых при такой постановке опытов доктором Лобановым.
 Вкладывается по 25 кусков, весом 100 г.
Мясо находилось в желудке 2 часа; переварилось без мнимого кормления 6.5%, с 8-минутнымнимым кормлением 31.6%.
Мясо находилось в желудке полтораса; переварилось без мнимого кормления 5.6%, с 5- 5-минутным мнимым кормлением -- 15 0/ %
Мясо находилось в желудке 5 часов; без мнимого кормления переварилось 58%, осталось 42%; с мнимым кормлением переварилось 85%, осталось 15%.
Я должен прибавить, однако, что опыт этот для демонстрации перед публикой довольно труден и нередко может не удаваться. С одной стороны, не легко скрыть от собаки вкладывание мяса, с другой - мнимое кормление в течение короткого времени, ввиду смущения животного при новой обстановке, не всегда может достичь желаемой энергии. Во избежание неудачи этот опыт лекционно можно ставить только на животных, привычных и с темпераментом которых экспериментатор хорошо знаком.
Из всего предшествующего, надеюсь, вы убедились, какое большое значение принадлежит акту прохождения пищи через ротовую и зевную полость или страстному желанию еды, что совершенно покрывается одно другим на основании раннего анализа. Без страстного желания, без аппетита некоторые пищевые вещества, хотя бы и попавшие в желудок, долго не получают на себя там никакого желудочного сока; другие, как мясо, хотя и обусловливают отделение, но слабого сока и не в таком большом количестве.
Ближе в смысл приведенных фактов мы можем войти только впоследствии, когда познакомимся с дальнейшими условиями от длительной работы желез. Почему хлеб, вложенный в желудок незаметно для собаки, часами не вызывает отделения сока, а мясо делает это довольно скоро, через десятки минут, - будет объяснено в следующей лекции, здесь же займемся пока некоторыми предварительными вопросами.
Как долго продолжается, отзвучивает первый удар по нервноотделительной системе желудка, как долго течет аппетитный сок при обыкновенной еде, которая, особенно у животных, продолжается недолго? Мы определяли много раз на разных собаках, помимо собаки с уединенным желудком, как велико так называемое в физиологии последействие приема мнимого кормления.
Вот опыт, сюда относящийся, из работы профессора Саноцкого. Гастро- и эзофаготомированная собака. После 5-минутного мнимого кормления началось отделение, продолжавшееся следующим образом (табл. 13).
Вкладывается по 25 кусков, весом 100 г.
Мясо находилось в желудке 2 часа; переварилось без мнимого кормления 6.5%, с 8-минутнымнимым кормлением 31.6%.
Мясо находилось в желудке полтораса; переварилось без мнимого кормления 5.6%, с 5- 5-минутным мнимым кормлением -- 15 0/ %
Мясо находилось в желудке 5 часов; без мнимого кормления переварилось 58%, осталось 42%; с мнимым кормлением переварилось 85%, осталось 15%.
Я должен прибавить, однако, что опыт этот для демонстрации перед публикой довольно труден и нередко может не удаваться. С одной стороны, не легко скрыть от собаки вкладывание мяса, с другой - мнимое кормление в течение короткого времени, ввиду смущения животного при новой обстановке, не всегда может достичь желаемой энергии. Во избежание неудачи этот опыт лекционно можно ставить только на животных, привычных и с темпераментом которых экспериментатор хорошо знаком.
Из всего предшествующего, надеюсь, вы убедились, какое большое значение принадлежит акту прохождения пищи через ротовую и зевную полость или страстному желанию еды, что совершенно покрывается одно другим на основании раннего анализа. Без страстного желания, без аппетита некоторые пищевые вещества, хотя бы и попавшие в желудок, долго не получают на себя там никакого желудочного сока; другие, как мясо, хотя и обусловливают отделение, но слабого сока и не в таком большом количестве.
Ближе в смысл приведенных фактов мы можем войти только впоследствии, когда познакомимся с дальнейшими условиями от длительной работы желез. Почему хлеб, вложенный в желудок незаметно для собаки, часами не вызывает отделения сока, а мясо делает это довольно скоро, через десятки минут, - будет объяснено в следующей лекции, здесь же займемся пока некоторыми предварительными вопросами.
Как долго продолжается, отзвучивает первый удар по нервноотделительной системе желудка, как долго течет аппетитный сок при обыкновенной еде, которая, особенно у животных, продолжается недолго? Мы определяли много раз на разных собаках, помимо собаки с уединенным желудком, как велико так называемое в физиологии последействие приема мнимого кормления.
Вот опыт, сюда относящийся, из работы профессора Саноцкого. Гастро- и эзофаготомированная собака. После 5-минутного мнимого кормления началось отделение, продолжавшееся следующим образом (табл. 13).
 Итак, эффект мнимого кормления, хотя бы и кратковременного, затягивается. То же, конечно, необходимо допустить и при настоящей еде. Но нельзя не иметь в виду, что в то время как при мнимом кормлении есть все данные для продления его сокогонного эффекта (свежесть и реальность впечатления при отсутствии удовлетворения голода, т. е. разжигание желания, которое и есть действующая сила), при настоящей еде, наоборот, удовлетворения желания, чувство насыщения, как известно, наступающее гораздо ранее заканчивания процесса пищеварения, только вследствие наполнения желудка, его растяжения, должно прерывать желание еды, а с ним и его сокогонное действие. Потому представляется невероятным, чтобы весь отделительный процесс в желудке, при иных количествах и сортах еды затягивающийся до 10-12 часов, мог бы быть отнесен весь на счет исследуемого нами до сих пор фактора, тем более что 5-минутное мнимое кормление, при самых благоприятных условиях, вызывает отделение самое большее на 3-4 часа. Мы принуждены, таким образом, искать дальнейших возбудителей иннервационного прибора желудочных желез. Чем же и как продолжается отделение желудочного сока, начатое психическим моментом? Пее Первое, что придет всем в голову при этом вопросе, - это, конечно, действие пищи, находящейся велудке, на самые стенки желудка. Да, это так и есть, но, наверное, не в том грубом и простом виде, как это представляют многие физиологи, а за ними, конечно, и врачи. Когда я заявил, что хлеб и яичный белок, вложенные прямо в желудок собаки, в продолжение часов не вызывают на себя ни малейшего отделения, вероятно, многие из моих слушателей-врачей пришли в полное недоумение: «Как же тогда понять насильственное кормление чахоточных, психически больных и кормление людей с желудочными фистулами вследствие того или другого закрытия пищевода?». Начну мой ответ с довольно неожиданного положения.
Утверждение, что механическое раздражение пищей стенок желудка есть верный и действительный возбудитель отделительной работы желудка, утверждение, так резко выражаемое в многих физиологических учебниках и так крепко засевшие в головах врачей, представляет собою ни более, ни менее как печальное заблуждение, приобревшее характер упорного предрассудка. Наши неоднократные заявления о фантастичности этого утверждения в статьях, на докторских диспутах, в заседаниях медицинских обществ большею частью встречали покачивания головой, а то и прямой отпор, что этого не может быть. Я искренно сожалею, что господа упорные отрицатели не пожаловали сюда, чтобы все наше дело с ними публично подвергнуть суду фактов, к которым мы теперь и переходим. Этому пункту я придаю весьма большое значение; на нем, по моему мнению, должно разыграться генеральное сражение господствующего взгляда о способности слизистой оболочки желудка раздражаться чем попало с теорией о специфической, разборчивой раздражительности этой оболочки. Раз у защитников старого взгляда будет отбита эта позиция (действительность механического раздражения), им ничего не останется, как обратиться к новой точке зрения и признать существенными стороны железистой работы, бывшие прежде совершенно в тени. Нужно думать, что потому-то главным образом так мало было обращено внимания на опыт Биддера и Шмидта о психическом отделении желудочного сока, что сильно верили в грубое и простое механическое раздражение, казавшееся таким верным и непременным. Мы воспроизведем перед вами опыт с механическим раздражением слизистой оболочки желудка прежде всего в его старой, нами заученной, классической форме.
Перед вами собака, имеющая обыкновенную желудочную Фистулу и перерезанный на шее пищевод. При открытии фистулы, как вы видите, из желудка ничего не вытекает; за час до этого желудок был чисто промыт водой. Мы берем пресловутые бородку пера и довольно толстую стеклянную палочку, а также несколько листов пропускной бумаги - одни окрашенные красным, кислым лакмусом, другие синим, щелочным. Я даю моему помощнику поручение в продолжение каждых пяти минут беспрерывно производить движения в полости желудка во всевозможных направлениях попеременно то бородкой пера, то стеклянной палочкой. Через каждые 5 минут одно орудие сменяется другим и, вынутое, тщательно вытирается как синим, так и красным лакмусовыми листами. Вы все видели, господа, что эта процедура настойчиво продолжалась в течение получаса. Ни одной капли сока не показалось из отверстия фистульной трубки, вместе с тем на лакмусовых листах, которые я вам передавал в течение этого получаса, все мокрые места, как вы в этом убедились сами, имеют синий оттенок на красных листах и происходили, очевидно, от щелочной слизи желудка, между тем как синие листы стали только мокроватыми, не изменив цвета. Следовательно, и в полости самого желудка, при таком настойчивом механическом раздражении, не оказалось ни одного пункта, который представлял бы хоть сколько-нибудь заметную кислую реакцию.
Где же струи чистого желудочного сока, о которых мы читали в учебниках? Что сказать против убедительности этого опыта? По-моему, только одно - что мы имели дело с больной собакой, почему-либо неспособной к нормальной деятельности желудочных желез. Это единственное возражение мы имеем, однако, возможность совершенно обессилить на ваших глазах. После неудач с механическим раздражением желудка мы сейчас же приступаем на этой собаке к опыту с мнимым кормлением. Собака ест предлагаемую пищу с большим аппетитом, и вы видите, что ровно через 5 минут после начала кормления показывается из желудка первая капля чистого сока, за которой следуют дальнейшие все чаще и чаще. Я принимаю несколько капель на синий лакмусовый лист. Вы видите яркокрасные пятна сильнокислого сока на листе. К концу лекции, т. е. за 30 минут продолжающегося мнимого кормления, мы получили 150 куб. см совершение чистого, как дистиллированная вода, даже без всякого фильтрования, желудочного сока. Нельзя сомневаться, что, когда был приложен действительный раздражитель, желудочные железа этой собаки отвечали на него вполне нормально и вполне нормальным соком, а отсюда неотразимо следует, что для первой, отрицательной половины опыта никакой другой причины быть не могло, кроме той, что слизистая оболочка желудка действительно абсолютно индифферентна к механическому раздражителю, что касается ее отделительной деятельности. И, однако, это механическое раздражение как возбудителя желудочных желез демонстрируют на лекциях физиологии. Смею думать, что те лекционные опыты должны будут отныне уступить место тому, который был проделан перед вами.
Повидимому, совершенно простой опыт с механическим раздражением желудка может быть правильно поставлен только при строгом соблюдении известных, хотя и очень простых правил, которых физиологи ранее как-то не принимали во внимание, очевидно, главным образом в силу предвзятого доверия к механическому раздражению. Этих правил два. Во-первых, необходимо, чтобы желудок был совершенно чист и чтобы в него ничего не поступало со стороны. Это условие раньше не соблюдалось. Из желудка выпускали его содержимое, открывая пробку фистульной трубки, но тщательно не промывали до полного исчезания кислой реакции, и в складках его всегда могли оставаться скопления ранней кислой жидкости. Вместе с тем всегда могла поступать из полости рта слона, которая в недостаточно вымытом желудке быстро подкислялась. Не мудрено, что при таком условии стеклянная трубка, вызывая движение желудка (отношение механического раздражения к двигательной способности желудка есть совершенно особая вещь от того, о чем до сих пор была речь), вела к выбрасыванию из фистульной трубки известного количества кислой жидкости. Что все это так, что наше объяснение отвечает вполне действительности, несомненно доказывается тем, что никогда и никто не получил таким способом настоящего, чистого желудочного сока с кислотностью 0.5-0.6%. Довольно напомнить, что Гейденгайн, когда сок из уединенного им желудка впервые был испытан на кислотность, пришел в немалое изумление от ее размера (0.5 0.6 %) и, не доверяя результату, просил своего тогдашнего ассистента Гшейдлена проверить все титры. Кислотность раннего, самого чистого сока еле достигала 0.3%. Кроме того, как дальнейшее доказательство, что прежние исследователи при механическом раздражении не имели настоящего, ответноео на него отделения, можно привести то обстоятельство, что никем из них не отмечен всегда строгий 5-минутный латентный период. Его нельзя было бы не заметить, если бы действительно имелось настоящее возбуждение желез.
Не менее важно второе условие правильного опыта с механическим раздражением. Требуется, конечно, чтобы желудочные железы были в недеятельном состоянии до опыта и чтобы во время опыта не существовали моменты, сами по себе, независимо от механического раздражения, способнызывать настоящую работу желез. В прошлом нет никаких указаний на то, чтобы ждали часами и убеждались в прекращении секреторной деятельности желудка. С другой стороне, не имеется ни малейших намеков, чтобы авторы сколько-нибудь обеспечивали себя против вмешательства в результат опыта психического отделения желудочного сока, а последнее, как уже сказано выше, дается чрезвычайно трудно. Есть такие возбудимые собаки, у которых почти невозможно достигнуть полного покоя желез или требуются для этого многие часы выжидания. Нужно чрезвычайное внимание экспериментатора, чтобы сделать наш опыт в безупречно чистой форме. Стоило стоять около собаки какойнибудь еде, даже быть только запаху от рук служителя, приготовлявшего еду, и массе еще более незначительных обстоятельств, чтобы палочка оказалась без вины виноватой в возбуждении желудочных желез. На собаке, которую вы видите перед собою, оба условия, очевидно, выполнены, результат опыта с ней стоит в неумолимом противоречии с прежними лекционными и лабораторными опытами по этому предмету.
Выясненная выше важность этого опыта дает мне право злоупотребить вашим вниманием и представить вам еще два видоизменения того же самого опыта. Кто-либо все же мог бы сказать, что для успеха механического раздражения требуется одновременное прикосновение механического деятеля к большому числу точек внутренней поверхности желудка. Ввиду такого предположения я проделаю перед вами две новых формы нашего опыта.
Опять такая же собака, т. е. гастро- изофаготомированная. Желудок чисто промыт и находится в полном отделительном покое. Я ввожу в него толстую стеклянную трубку с концом, усеянным дырочками, в 2 3мм в диаметре: другой конец трубки припаян к большому баллону, содержащему в себе довольно крупный песок; через другую трубку баллона посредством каучукового насоса я произвожу сильный вихрь песчаных частичек. Ритмически работая каучуковым баллоном, я с силою выбрасываю песок в желудок в продолжение 10-15 минут. Никакого намека на отделение желудочного сока! Высыпающийся между стеклянной и фистульной трубкой песок или совсем сухой, или слегка мокроватый, но отнюдь не красящий синий лакмусовый лист в красный цвет. Очевидно, в этой форме опыта мы имеем дело и с сильным и с широко распространенным раздражением. Прошу посмотреть на работу нашего приборчика вне желудка. Вы видите, как через дырочки трубки (не один десяток) с силою вырываются песчаные струи. Подставляя руку под струю, вы отчетливо чувствуете силу ударов многочисленных песчинок. И теперь, по окончании опыта с песком, мы легко и бесспорно, посредством опыта с мнимым кормлением, удостоверяемся в полной нормальности отделительных отношений нашей собаки.
Еще опыт. Опять такая же собака. Этой в пустой и покойный желудок мы вводим каучуковый шар и будем его постепенно надувать спринцовкой, например, до размеров головки младенца; оставим его минуту-другую раздутым и затем дадим ему спасться. Повторяем это в продолжение 10-15 минут. За все это время из желудка не вытекает ни капли сока. Поверхность вынутого в конце опыта шара всюду представляет только щелочную реакцию. И здесь последовательное мнимое кормление резко свидетельствует о полной годности собаки. Относительно этого опыта следует только заметить, что для него нужно брать не особенно голодных животных (10--12 часов после еды), иначе легко получить возбуждение желез.
Если бы смотреть на механическое раздражение беспристрастными глазами, то фиктивность его подтверждается в лаборатории на каждом шагу и, собственно говоря, лежит в основании всех наших методов, касающихся желудочного отделения. У собаки с обыкновенной желудочной фистулой вне пищеварительного периода и без особых причин из желудка не вытекает ни капли сока. Как бы это могло быть, если 6ы механическое раздражение было действительно? Ведь внутренний диск фистульной трубки постоянно находится в соприкосновении с слизистой оболочкой желудка. То же самое наблюдается и на собаке с уединенным желудочком; в этот желудочек на время опыта вставляется на порядочную глубину стеклянная или каучуковая трубка для собирания сока, и, однако, через нее не вытекает ни капли сока, и поверхность ее отнюдь не делается кислой, если нет налицо настоящих отделительных условий, а трубка часто вынимается и поправляется. У собак обыкновенной желудочной фистулой, если операция сделана давно (год и больше), над внутренним диском трубки собираются складки слизистой оболочки, так что отверстие трубки ими совершенно закрывается; в таком случае приходится вводить через фистульное отверстие длинную и толстую металлическую трубку с дырчатыми стенками на значительную глубину, и это, однако, само по себе не влечет за собой ни одной капли сока. Далее - весьма обычная вещь, что в желудке собак встречаются большие комья волос, и все же нахождение их нисколько не мешает полному перерыву отделительной деятельности желудка вне часов пищеварения. Особенно ярким подобное отношение выступает неоднократно у нашей собаки с двойным желудочком, когда ей устраивали подстилку из древесных опилок с целью предупредить разъедание раны изливающимся соком. Сплошь и рядом в большом желудке ее находили тогда огромные массы этих опилок, иногда до полуфунта разом. Очевидно, собака облизывала рану и при этом глотала приставшие ко рту опилки. Однако эти опилки, лежа в желудке, сами по себе не возбуждали ни малейшего отделения, хотя механический эффект этих опилок, как каждый может себе представить, весьма значителен. Мне кажется, что этой длинной вереницы представленных фактов вполне достаточно, чтобы окончательно похоронить всякую мысль о возможности привести непосредственно в действие нервноотделительный прибор желудка путем механического раздражения его слизистой оболочки.
И, однако, до последнего времени в некоторых учебниках и книжках, касающихся деятельности желудка, продолжают фигурировать в роли возбудителей желудочных желез бородка пера и стеклянная палочка. Правда, найдется не мало физиологов, считающих механическое раздражение, по отношению к отделутельной деятельности желудка, не особенно сильным, скорее второстепенным, рядом с другими раздражителями. Но я сейчас не знаю физиолога-автора, который бы отрицал вообще его действительность и не верил в возможность получить посредством его хоть немного сока.
В заключение этой лекции мы остановимся несколько на одном пункте, стоящем в известном отношении к занимавшему нас вопросу. Если прикосновение пищи к слизистой оболочке желудка не вызывает отделения сока непосредственно, то не стоит ли, однако, поступление пищи в желудок в какой-нибудь косвенной связи с отделительным процессом?
Едва ли можно сомневаться, что и при нормальных условиях желудок является местом известных ощущений, т. е. его внутренняя оболочка обладает известной степенью чувства осязания. Эти ощущения в общем очень слабы, и большинство людей привыкает совершенно отвлекаться от них при нормальном ходе питания, так что материал их входит лишь бессознательно в чувство благополучия вообще и наслаждения едой в частности. Что все это действительно так, следует из факто известного ощущения при голоде, относимого именно к области желудка. С другой стороны, конечно, каждому приходилось встречать лиц, описывающих подробно и любовно, как кусок какой-нибудь любимой еды или глоток любимого питья, особенно на пустой желудок, чувствуется ими на всем ходу по пищеводу и желудку. Конечно, любитель еды, концентрируя постоянно внимание на акте еды, в конце концов будет отчетливо чувствовать и сознавать то, что у другого нормально заслонено другими ощущениями и впечатлениями. Нужно думать поэтому, что в наслаждение едой входят не только разнообразные раздражения полости рта и зева, как у наших животных при мнимом кормлении, но и раздражение пищей и возникновение ощущений в дальнейших отделах пищеварительного канала, до желудка включительно. Иначе сказать, пища, проходящая только через рот и зев, может вызывать меньшее наслаждение едой и отсюда меньшее желание еды, чем пища, проходящая весь путь, вплоть до желудка. Аппетит как страстное желание еды есть, конечно, сложное ощущение, и для наличности его во многих случаях требуются не только существование в организме потребности в новом питательном материале, но и состояние полного здоровья, ощущение этого здоровья во всех аппаратах пищеварительного канала. Таким образом будет понятным, что люди, испытывающие болезненные ощущения в этих аппаратах и сознательно или бессознательно помнящие их, хотя бы они в данный момент даже не имелись налицо, могут в некоторых случаях не чувствовать аппетита, не иметь потребности в еде. Известны невропатологические случаи, где такой потерей аппетита страдали люди c анестезией желудка: они как бы не чувствовали у себя желудка, и это восстановляло их против акта еды; пища, по их словам, словно проваливалась в пустой, чужой мешок. Точно так же можно себе представить у некоторых людей потерю аппетита вследствие долговременного закрытия, по той или другой причине, пищевода; они могли бы как бы забывать свой желудок, и в таком случае вкладывание пищи прямо в желудок после операции могло бы сопровождаться взрывом аппетита. Позволяю себе для дальнейшей иллюстрации того же привести факт из личной жизни.
После какой-то мимолетной, но сильной лихорадочной формы я, совершенно оправившись в остальном, потерял всякий позыв к еде. Было даже что-то забавное в этом полном равнодушии к пище. Совершенно здоровый, я, однако, резко отличался от других тем, что, повидимому, с легкостью мог обходиться совершенно без всякой еды. Боясь сильного истощения, я через дватри дня такого состояния решил, для возвращения аппетита, выпить вина. При первом же глотке я живо почувствовал движение его по пищеводу и в желудке и буквально моментально испытал приступ сильного аппетита. Смысл приведенных наблюдений состоит в том, что осязание, так сказать, желудком входящей пищи может служить или толчком к возбуждению аппетита, или условием, его усиливающим. Известно, что недостаток в организме питательных веществ, или, лучше, потребность еды, не ведет всегда и сейчас же к аппетиту, к страстному желанию есть. Как часто случается, что давно прошли часы обычной еды, а вы, занятые, отвлеченные чем-нибудь, не испытываете никакого делания есть. Всем хорошо известно, это вошло даже в пословицу, что еда во рту вызывает аппетит. Если это так, то в иных случаях первый толчок к пробуждению аппетита может быть дан в желудке, а не во рту. Конечно, ранее, когда говорилось о желании еды как возбудителе секреторных нервов желудочных желез, разумелось именно страстное и сознаваемое желание еды, именно то, что называется аппетитом, а не недостаток питательных веществ в организме, скрытая потребность еды, еще не перешедшая в определенное страстное желание. Лучший пример полной отдельности этих моментов представляют наши собаки в опытах с мнимым кормлением. Потребность в еде имеется у них и раньше опыта, однако сок течет только тогда, когда потребность эта выливается в форму страстного желания. Таким образом возможен случай, что у иных собак, при известных степенях голодания, прикосновение каких-нибудь тел к слизистой оболочке желудка, механическое раздражение желудка или растяжение его вкладываемыми массами могут подать повод к возбуждению аппетита, а возбудится он - появится и сок. Вот и третье основание для старого опыта с мнимой действительностью механического раздражения. Такая точка зрения была бы до некоторой степени примирением между моим утверждением о недействительности механического раздражения и всеобщею верой в его силу. И я допускаю, что механические свойства пищи иногда могут обусловить работу желудочных желез, но не прямо, простым физиологическим рефлексом, а косвенно, предварительно возбудив, оживив представление о еде и, таким образом, вызвав страстное желание еды. Надеюсь, что вто нисколько не путает предмета и только лишний раз оттеняет старую, грубую точку зрения на наши явления от подробного и конкретного анализа их. Конечно, и этот пункт, носящий у нас поневоле более предположительный характер, может быть подвергнут экспериментальному исследованию: стоит только сравнить силу эффекта мнимого кормления собак с перерезанным пищеводом и просто гастротомированных, при открытой фистуле.
Итак, эффект мнимого кормления, хотя бы и кратковременного, затягивается. То же, конечно, необходимо допустить и при настоящей еде. Но нельзя не иметь в виду, что в то время как при мнимом кормлении есть все данные для продления его сокогонного эффекта (свежесть и реальность впечатления при отсутствии удовлетворения голода, т. е. разжигание желания, которое и есть действующая сила), при настоящей еде, наоборот, удовлетворения желания, чувство насыщения, как известно, наступающее гораздо ранее заканчивания процесса пищеварения, только вследствие наполнения желудка, его растяжения, должно прерывать желание еды, а с ним и его сокогонное действие. Потому представляется невероятным, чтобы весь отделительный процесс в желудке, при иных количествах и сортах еды затягивающийся до 10-12 часов, мог бы быть отнесен весь на счет исследуемого нами до сих пор фактора, тем более что 5-минутное мнимое кормление, при самых благоприятных условиях, вызывает отделение самое большее на 3-4 часа. Мы принуждены, таким образом, искать дальнейших возбудителей иннервационного прибора желудочных желез. Чем же и как продолжается отделение желудочного сока, начатое психическим моментом? Пее Первое, что придет всем в голову при этом вопросе, - это, конечно, действие пищи, находящейся велудке, на самые стенки желудка. Да, это так и есть, но, наверное, не в том грубом и простом виде, как это представляют многие физиологи, а за ними, конечно, и врачи. Когда я заявил, что хлеб и яичный белок, вложенные прямо в желудок собаки, в продолжение часов не вызывают на себя ни малейшего отделения, вероятно, многие из моих слушателей-врачей пришли в полное недоумение: «Как же тогда понять насильственное кормление чахоточных, психически больных и кормление людей с желудочными фистулами вследствие того или другого закрытия пищевода?». Начну мой ответ с довольно неожиданного положения.
Утверждение, что механическое раздражение пищей стенок желудка есть верный и действительный возбудитель отделительной работы желудка, утверждение, так резко выражаемое в многих физиологических учебниках и так крепко засевшие в головах врачей, представляет собою ни более, ни менее как печальное заблуждение, приобревшее характер упорного предрассудка. Наши неоднократные заявления о фантастичности этого утверждения в статьях, на докторских диспутах, в заседаниях медицинских обществ большею частью встречали покачивания головой, а то и прямой отпор, что этого не может быть. Я искренно сожалею, что господа упорные отрицатели не пожаловали сюда, чтобы все наше дело с ними публично подвергнуть суду фактов, к которым мы теперь и переходим. Этому пункту я придаю весьма большое значение; на нем, по моему мнению, должно разыграться генеральное сражение господствующего взгляда о способности слизистой оболочки желудка раздражаться чем попало с теорией о специфической, разборчивой раздражительности этой оболочки. Раз у защитников старого взгляда будет отбита эта позиция (действительность механического раздражения), им ничего не останется, как обратиться к новой точке зрения и признать существенными стороны железистой работы, бывшие прежде совершенно в тени. Нужно думать, что потому-то главным образом так мало было обращено внимания на опыт Биддера и Шмидта о психическом отделении желудочного сока, что сильно верили в грубое и простое механическое раздражение, казавшееся таким верным и непременным. Мы воспроизведем перед вами опыт с механическим раздражением слизистой оболочки желудка прежде всего в его старой, нами заученной, классической форме.
Перед вами собака, имеющая обыкновенную желудочную Фистулу и перерезанный на шее пищевод. При открытии фистулы, как вы видите, из желудка ничего не вытекает; за час до этого желудок был чисто промыт водой. Мы берем пресловутые бородку пера и довольно толстую стеклянную палочку, а также несколько листов пропускной бумаги - одни окрашенные красным, кислым лакмусом, другие синим, щелочным. Я даю моему помощнику поручение в продолжение каждых пяти минут беспрерывно производить движения в полости желудка во всевозможных направлениях попеременно то бородкой пера, то стеклянной палочкой. Через каждые 5 минут одно орудие сменяется другим и, вынутое, тщательно вытирается как синим, так и красным лакмусовыми листами. Вы все видели, господа, что эта процедура настойчиво продолжалась в течение получаса. Ни одной капли сока не показалось из отверстия фистульной трубки, вместе с тем на лакмусовых листах, которые я вам передавал в течение этого получаса, все мокрые места, как вы в этом убедились сами, имеют синий оттенок на красных листах и происходили, очевидно, от щелочной слизи желудка, между тем как синие листы стали только мокроватыми, не изменив цвета. Следовательно, и в полости самого желудка, при таком настойчивом механическом раздражении, не оказалось ни одного пункта, который представлял бы хоть сколько-нибудь заметную кислую реакцию.
Где же струи чистого желудочного сока, о которых мы читали в учебниках? Что сказать против убедительности этого опыта? По-моему, только одно - что мы имели дело с больной собакой, почему-либо неспособной к нормальной деятельности желудочных желез. Это единственное возражение мы имеем, однако, возможность совершенно обессилить на ваших глазах. После неудач с механическим раздражением желудка мы сейчас же приступаем на этой собаке к опыту с мнимым кормлением. Собака ест предлагаемую пищу с большим аппетитом, и вы видите, что ровно через 5 минут после начала кормления показывается из желудка первая капля чистого сока, за которой следуют дальнейшие все чаще и чаще. Я принимаю несколько капель на синий лакмусовый лист. Вы видите яркокрасные пятна сильнокислого сока на листе. К концу лекции, т. е. за 30 минут продолжающегося мнимого кормления, мы получили 150 куб. см совершение чистого, как дистиллированная вода, даже без всякого фильтрования, желудочного сока. Нельзя сомневаться, что, когда был приложен действительный раздражитель, желудочные железа этой собаки отвечали на него вполне нормально и вполне нормальным соком, а отсюда неотразимо следует, что для первой, отрицательной половины опыта никакой другой причины быть не могло, кроме той, что слизистая оболочка желудка действительно абсолютно индифферентна к механическому раздражителю, что касается ее отделительной деятельности. И, однако, это механическое раздражение как возбудителя желудочных желез демонстрируют на лекциях физиологии. Смею думать, что те лекционные опыты должны будут отныне уступить место тому, который был проделан перед вами.
Повидимому, совершенно простой опыт с механическим раздражением желудка может быть правильно поставлен только при строгом соблюдении известных, хотя и очень простых правил, которых физиологи ранее как-то не принимали во внимание, очевидно, главным образом в силу предвзятого доверия к механическому раздражению. Этих правил два. Во-первых, необходимо, чтобы желудок был совершенно чист и чтобы в него ничего не поступало со стороны. Это условие раньше не соблюдалось. Из желудка выпускали его содержимое, открывая пробку фистульной трубки, но тщательно не промывали до полного исчезания кислой реакции, и в складках его всегда могли оставаться скопления ранней кислой жидкости. Вместе с тем всегда могла поступать из полости рта слона, которая в недостаточно вымытом желудке быстро подкислялась. Не мудрено, что при таком условии стеклянная трубка, вызывая движение желудка (отношение механического раздражения к двигательной способности желудка есть совершенно особая вещь от того, о чем до сих пор была речь), вела к выбрасыванию из фистульной трубки известного количества кислой жидкости. Что все это так, что наше объяснение отвечает вполне действительности, несомненно доказывается тем, что никогда и никто не получил таким способом настоящего, чистого желудочного сока с кислотностью 0.5-0.6%. Довольно напомнить, что Гейденгайн, когда сок из уединенного им желудка впервые был испытан на кислотность, пришел в немалое изумление от ее размера (0.5 0.6 %) и, не доверяя результату, просил своего тогдашнего ассистента Гшейдлена проверить все титры. Кислотность раннего, самого чистого сока еле достигала 0.3%. Кроме того, как дальнейшее доказательство, что прежние исследователи при механическом раздражении не имели настоящего, ответноео на него отделения, можно привести то обстоятельство, что никем из них не отмечен всегда строгий 5-минутный латентный период. Его нельзя было бы не заметить, если бы действительно имелось настоящее возбуждение желез.
Не менее важно второе условие правильного опыта с механическим раздражением. Требуется, конечно, чтобы желудочные железы были в недеятельном состоянии до опыта и чтобы во время опыта не существовали моменты, сами по себе, независимо от механического раздражения, способнызывать настоящую работу желез. В прошлом нет никаких указаний на то, чтобы ждали часами и убеждались в прекращении секреторной деятельности желудка. С другой стороне, не имеется ни малейших намеков, чтобы авторы сколько-нибудь обеспечивали себя против вмешательства в результат опыта психического отделения желудочного сока, а последнее, как уже сказано выше, дается чрезвычайно трудно. Есть такие возбудимые собаки, у которых почти невозможно достигнуть полного покоя желез или требуются для этого многие часы выжидания. Нужно чрезвычайное внимание экспериментатора, чтобы сделать наш опыт в безупречно чистой форме. Стоило стоять около собаки какойнибудь еде, даже быть только запаху от рук служителя, приготовлявшего еду, и массе еще более незначительных обстоятельств, чтобы палочка оказалась без вины виноватой в возбуждении желудочных желез. На собаке, которую вы видите перед собою, оба условия, очевидно, выполнены, результат опыта с ней стоит в неумолимом противоречии с прежними лекционными и лабораторными опытами по этому предмету.
Выясненная выше важность этого опыта дает мне право злоупотребить вашим вниманием и представить вам еще два видоизменения того же самого опыта. Кто-либо все же мог бы сказать, что для успеха механического раздражения требуется одновременное прикосновение механического деятеля к большому числу точек внутренней поверхности желудка. Ввиду такого предположения я проделаю перед вами две новых формы нашего опыта.
Опять такая же собака, т. е. гастро- изофаготомированная. Желудок чисто промыт и находится в полном отделительном покое. Я ввожу в него толстую стеклянную трубку с концом, усеянным дырочками, в 2 3мм в диаметре: другой конец трубки припаян к большому баллону, содержащему в себе довольно крупный песок; через другую трубку баллона посредством каучукового насоса я произвожу сильный вихрь песчаных частичек. Ритмически работая каучуковым баллоном, я с силою выбрасываю песок в желудок в продолжение 10-15 минут. Никакого намека на отделение желудочного сока! Высыпающийся между стеклянной и фистульной трубкой песок или совсем сухой, или слегка мокроватый, но отнюдь не красящий синий лакмусовый лист в красный цвет. Очевидно, в этой форме опыта мы имеем дело и с сильным и с широко распространенным раздражением. Прошу посмотреть на работу нашего приборчика вне желудка. Вы видите, как через дырочки трубки (не один десяток) с силою вырываются песчаные струи. Подставляя руку под струю, вы отчетливо чувствуете силу ударов многочисленных песчинок. И теперь, по окончании опыта с песком, мы легко и бесспорно, посредством опыта с мнимым кормлением, удостоверяемся в полной нормальности отделительных отношений нашей собаки.
Еще опыт. Опять такая же собака. Этой в пустой и покойный желудок мы вводим каучуковый шар и будем его постепенно надувать спринцовкой, например, до размеров головки младенца; оставим его минуту-другую раздутым и затем дадим ему спасться. Повторяем это в продолжение 10-15 минут. За все это время из желудка не вытекает ни капли сока. Поверхность вынутого в конце опыта шара всюду представляет только щелочную реакцию. И здесь последовательное мнимое кормление резко свидетельствует о полной годности собаки. Относительно этого опыта следует только заметить, что для него нужно брать не особенно голодных животных (10--12 часов после еды), иначе легко получить возбуждение желез.
Если бы смотреть на механическое раздражение беспристрастными глазами, то фиктивность его подтверждается в лаборатории на каждом шагу и, собственно говоря, лежит в основании всех наших методов, касающихся желудочного отделения. У собаки с обыкновенной желудочной фистулой вне пищеварительного периода и без особых причин из желудка не вытекает ни капли сока. Как бы это могло быть, если 6ы механическое раздражение было действительно? Ведь внутренний диск фистульной трубки постоянно находится в соприкосновении с слизистой оболочкой желудка. То же самое наблюдается и на собаке с уединенным желудочком; в этот желудочек на время опыта вставляется на порядочную глубину стеклянная или каучуковая трубка для собирания сока, и, однако, через нее не вытекает ни капли сока, и поверхность ее отнюдь не делается кислой, если нет налицо настоящих отделительных условий, а трубка часто вынимается и поправляется. У собак обыкновенной желудочной фистулой, если операция сделана давно (год и больше), над внутренним диском трубки собираются складки слизистой оболочки, так что отверстие трубки ими совершенно закрывается; в таком случае приходится вводить через фистульное отверстие длинную и толстую металлическую трубку с дырчатыми стенками на значительную глубину, и это, однако, само по себе не влечет за собой ни одной капли сока. Далее - весьма обычная вещь, что в желудке собак встречаются большие комья волос, и все же нахождение их нисколько не мешает полному перерыву отделительной деятельности желудка вне часов пищеварения. Особенно ярким подобное отношение выступает неоднократно у нашей собаки с двойным желудочком, когда ей устраивали подстилку из древесных опилок с целью предупредить разъедание раны изливающимся соком. Сплошь и рядом в большом желудке ее находили тогда огромные массы этих опилок, иногда до полуфунта разом. Очевидно, собака облизывала рану и при этом глотала приставшие ко рту опилки. Однако эти опилки, лежа в желудке, сами по себе не возбуждали ни малейшего отделения, хотя механический эффект этих опилок, как каждый может себе представить, весьма значителен. Мне кажется, что этой длинной вереницы представленных фактов вполне достаточно, чтобы окончательно похоронить всякую мысль о возможности привести непосредственно в действие нервноотделительный прибор желудка путем механического раздражения его слизистой оболочки.
И, однако, до последнего времени в некоторых учебниках и книжках, касающихся деятельности желудка, продолжают фигурировать в роли возбудителей желудочных желез бородка пера и стеклянная палочка. Правда, найдется не мало физиологов, считающих механическое раздражение, по отношению к отделутельной деятельности желудка, не особенно сильным, скорее второстепенным, рядом с другими раздражителями. Но я сейчас не знаю физиолога-автора, который бы отрицал вообще его действительность и не верил в возможность получить посредством его хоть немного сока.
В заключение этой лекции мы остановимся несколько на одном пункте, стоящем в известном отношении к занимавшему нас вопросу. Если прикосновение пищи к слизистой оболочке желудка не вызывает отделения сока непосредственно, то не стоит ли, однако, поступление пищи в желудок в какой-нибудь косвенной связи с отделительным процессом?
Едва ли можно сомневаться, что и при нормальных условиях желудок является местом известных ощущений, т. е. его внутренняя оболочка обладает известной степенью чувства осязания. Эти ощущения в общем очень слабы, и большинство людей привыкает совершенно отвлекаться от них при нормальном ходе питания, так что материал их входит лишь бессознательно в чувство благополучия вообще и наслаждения едой в частности. Что все это действительно так, следует из факто известного ощущения при голоде, относимого именно к области желудка. С другой стороны, конечно, каждому приходилось встречать лиц, описывающих подробно и любовно, как кусок какой-нибудь любимой еды или глоток любимого питья, особенно на пустой желудок, чувствуется ими на всем ходу по пищеводу и желудку. Конечно, любитель еды, концентрируя постоянно внимание на акте еды, в конце концов будет отчетливо чувствовать и сознавать то, что у другого нормально заслонено другими ощущениями и впечатлениями. Нужно думать поэтому, что в наслаждение едой входят не только разнообразные раздражения полости рта и зева, как у наших животных при мнимом кормлении, но и раздражение пищей и возникновение ощущений в дальнейших отделах пищеварительного канала, до желудка включительно. Иначе сказать, пища, проходящая только через рот и зев, может вызывать меньшее наслаждение едой и отсюда меньшее желание еды, чем пища, проходящая весь путь, вплоть до желудка. Аппетит как страстное желание еды есть, конечно, сложное ощущение, и для наличности его во многих случаях требуются не только существование в организме потребности в новом питательном материале, но и состояние полного здоровья, ощущение этого здоровья во всех аппаратах пищеварительного канала. Таким образом будет понятным, что люди, испытывающие болезненные ощущения в этих аппаратах и сознательно или бессознательно помнящие их, хотя бы они в данный момент даже не имелись налицо, могут в некоторых случаях не чувствовать аппетита, не иметь потребности в еде. Известны невропатологические случаи, где такой потерей аппетита страдали люди c анестезией желудка: они как бы не чувствовали у себя желудка, и это восстановляло их против акта еды; пища, по их словам, словно проваливалась в пустой, чужой мешок. Точно так же можно себе представить у некоторых людей потерю аппетита вследствие долговременного закрытия, по той или другой причине, пищевода; они могли бы как бы забывать свой желудок, и в таком случае вкладывание пищи прямо в желудок после операции могло бы сопровождаться взрывом аппетита. Позволяю себе для дальнейшей иллюстрации того же привести факт из личной жизни.
После какой-то мимолетной, но сильной лихорадочной формы я, совершенно оправившись в остальном, потерял всякий позыв к еде. Было даже что-то забавное в этом полном равнодушии к пище. Совершенно здоровый, я, однако, резко отличался от других тем, что, повидимому, с легкостью мог обходиться совершенно без всякой еды. Боясь сильного истощения, я через дватри дня такого состояния решил, для возвращения аппетита, выпить вина. При первом же глотке я живо почувствовал движение его по пищеводу и в желудке и буквально моментально испытал приступ сильного аппетита. Смысл приведенных наблюдений состоит в том, что осязание, так сказать, желудком входящей пищи может служить или толчком к возбуждению аппетита, или условием, его усиливающим. Известно, что недостаток в организме питательных веществ, или, лучше, потребность еды, не ведет всегда и сейчас же к аппетиту, к страстному желанию есть. Как часто случается, что давно прошли часы обычной еды, а вы, занятые, отвлеченные чем-нибудь, не испытываете никакого делания есть. Всем хорошо известно, это вошло даже в пословицу, что еда во рту вызывает аппетит. Если это так, то в иных случаях первый толчок к пробуждению аппетита может быть дан в желудке, а не во рту. Конечно, ранее, когда говорилось о желании еды как возбудителе секреторных нервов желудочных желез, разумелось именно страстное и сознаваемое желание еды, именно то, что называется аппетитом, а не недостаток питательных веществ в организме, скрытая потребность еды, еще не перешедшая в определенное страстное желание. Лучший пример полной отдельности этих моментов представляют наши собаки в опытах с мнимым кормлением. Потребность в еде имеется у них и раньше опыта, однако сок течет только тогда, когда потребность эта выливается в форму страстного желания. Таким образом возможен случай, что у иных собак, при известных степенях голодания, прикосновение каких-нибудь тел к слизистой оболочке желудка, механическое раздражение желудка или растяжение его вкладываемыми массами могут подать повод к возбуждению аппетита, а возбудится он - появится и сок. Вот и третье основание для старого опыта с мнимой действительностью механического раздражения. Такая точка зрения была бы до некоторой степени примирением между моим утверждением о недействительности механического раздражения и всеобщею верой в его силу. И я допускаю, что механические свойства пищи иногда могут обусловить работу желудочных желез, но не прямо, простым физиологическим рефлексом, а косвенно, предварительно возбудив, оживив представление о еде и, таким образом, вызвав страстное желание еды. Надеюсь, что вто нисколько не путает предмета и только лишний раз оттеняет старую, грубую точку зрения на наши явления от подробного и конкретного анализа их. Конечно, и этот пункт, носящий у нас поневоле более предположительный характер, может быть подвергнут экспериментальному исследованию: стоит только сравнить силу эффекта мнимого кормления собак с перерезанным пищеводом и просто гастротомированных, при открытой фистуле.
 Этот опыт интересен в том отношении, что дает довольно существенную опору допущению, молчаливо желаемому до сих пор, именно, что все испытанные до сих пор вещества действовали на нервную систему желез, раздражая слизистую оболочку желудка, а не через то, что всасывались в кровь и тогда уже раздражали или нервную систему желез, или самые железы непосредственно. Ясно, что, будь действие на железы через кровь, один раствор экстракта действовал бы гораздо сильнее, чем связанный крахмалом и затрудненный в отношение всасывания.
Особенно подробным исследованиям был подвергнут жир, как растительный, так и животный. Он испытывался на собаке с уединенным желудочком, на собаках с обыкновенной желудочной фистулой и перерезанным пищеводом, наконец, на собаке, пережившей многие месяцы перерезке блуждающего нерва на шее. Во всех опытах жир вводился прямо в желудок, т. е. при исключении акта еды. Результат всегда оставался отрицательным.
Итак, из всех пищевых веществ в широком смысле слова, при исследовании их врозь, большинство их оназалось без сокогонного эффекта в отношении желудка, меньшинство составили вода и неизвестные еще точно составные части мяса, переходящие в раствор.
От этих опытов с элементарными веществами перейдем к опытам вкладывания в желудок, присключении акта еды, различных сортов обыкновенной пищи и зададимся вопросом: насколько действие этих сложных веществ слагается и объясняется из элементарных вышеуказанных действий?
Если вложим в большой желудок нашей собаки с уединенным желудком, незаметно для нее, известное количество молотого сырого мяса, то через 15-30 минут (в лучшем случае) начинается отделение сока, как уже сказано в предшествующей лекции. Считаю нелишним упомянуть здесь о выработанном нами (д-р Лобасов) способе введения мяса. При вкладывании мяса кусочками собака часто догадывается, что с неи делается, и это, конечно, может повести к психическому отделению. Если бы собака и спала, то процедура вкладывания всегда ее будит и оканчивается при бодром состоянии животного. Для избежания этих неудобств мы накладываем молотое мясо в широкую стеклянную трубку, а затем, во время сна собаки, введя ее на небольшую глубину в фистульную трубку, при помощи поршня быстро вталкиваем мясо в желудок. Проснувшись, собака, однако, не может догадаться, что с ней было, так как все уже кончено, и сплошь и рядом сейчас же засыпает опять. Но и теперь мясо всегда вызывает себя отделение сока. После сообщенного в начале этой лекции факт этот, разумеется, не представляет ничего неожиданного и непонятного. Очевидно, раствор некоторых химических веществ, находящихся в мясном соке, есть главнейшая причина начинающегося теперь отделения. Доктор Лобасов, занимавшийся этим вопросом, поставил много видоизменений опыта с целью как можно более усилить это заключение. Он, например, сильно вываривал мясо в продолжение нескольких дней и, вкладывая его в желудок, убеждался, что сокогонное действие теперь или очень ослаблалось, или исчезало совершенно. Но стоило прибавить к такому мясу либиховского экстракта, для того чтобы действие, свойственное сырому мясу, восстановилось.
Совершенно такие же опыты со вкладыванием хлеба и вареного яичного белка, т. е. при полном исключении психического момента, как уже сказано в предшествующей лекции, дают совершенно отрицательный результат; эти сорта еды остаются в желудке 2 часа (до тех пор, пока за ними наблюдали) без малейшего возбуждающего действия на желудочные железы. Мы можем представить себе, что частью их Физико-химическое состояние (связанная вода), частью отсутствие в них непосредственно раздражающих химических веществ обусловливают этот неожиданный факт. Что касается до яичного белка, то стоит только припомнить результаты опытов с вливанием жидкого белка; а относительно хлеба доктор Лобасов нашел, что продолжительные настои его с водой едва ли действуют в качестве возбудителей желудочных желез энергичнее воды.
Можно ли удовлетвориться полученными результатами? Дают ли они нам полное разъяснение отделительного процесса при нормальной еде? Очевидно, нет. Если с едой мяса дело может считаться более или менее законченным - сок льется на него частью вследствие психического момента, частью вследствие собственного химически раздражающего действия, - то того же нельзя сказать об отделительной работе желудка при еде хлеба и вареного яичного белка. При них оказывается объясненным только первый период отделения, имеющий свое основание в психическом моменте; дальнейший же период после трех-четырех часов остается темным в своем механизме, -так как только часть выливающегося на них сока покрывается из известного нам источника. Для лучшего уяснения смысла занимающего нас вопроса я нахожу полезным предварительно остановить ваше внимание на сопоставлении опытов вкладыванием и с едой крахмала.
Как уже раньше сказано, чистый крахмал, вложенный прямо в желудок, не обусловливает сколько-нибудь значительного отделения, при еде же его животным наступает отделение в продолжение двух-трех часов. Внимательный разбор этих опытов скоро раскрывает весь их механизм. Отделяющийся при еде крахмала сок вполне покрывается психическим отделением, как оно установлено в опыте с мнимым кормлением. Этой точности итога для массы сока и нет в случае хлеба и вареного яичного белка; только 1 / или 1 /2 сока, на них изливающегося, может быть отнесена на психический момент, а источник остальной части предстоит отыскать. Что действительно имеется другой источник, другой раздражитель, в частности при еде белка, следует и из того, что сок второго и третьего часа обладает не особенно высокой переваривающей силой, между тем как психический сок, как уже замечено выше, по переваривающей силе принадлежит к сильнейшим сортам. Всестественнее допустить, что при изменении хлеба и вареного яичного белка посредством психического аппетитного сока в начале нормального пищеварения в них возникает нечто химическое, производящее раздражающее действие на нервноотделительный прибор желудка. Вероятно, это какие-нибудь продукты переваривания, тождественные или близкие к тем веществам, которые являются возбудителями в мясе.
В пользу такого понимания наблюдаемых фактов можно привести несколько экспериментальных данных. Если собирать жидкий продукт переваривания твердого яичного белка, скопляющийся в желудке собаки, и ввести его прямо в большой желудок нашей собаки с уединенным желудочком, то нельзя не заметить гораздо более верного и несколько более значительного сокогонного действия сравнительно с такою же массою воды или жидкого белка (д-р Лобасов). Во всяком случае образование этих веществ не может быть очень значительным, так как и яичный белок и хлеб при их поедании животным, за вычетом психического отделения, которое продолжается 2 3 часа, представляют вообще скудное отделение по часам, начиная с третьего-четвертого часа. Дальнейшим доказательством нашего толкования может служить следующий опыт. Если в желудке имеется уже отделение желудочного сока, вызванное или психически, или вообще предшествующим пищеварением, то введение в желудок (незаметно для животного) жидкого яичного белка обыкновенно сопровождается значительным взмахом отделения (д-р Хижин). Как понять это, если не допустить, что при начинающемся переваривании белка из него образуется нечто раздражающее слизистую оболочку? Приняв такое толкование явлений относительно хлеба и яичного белка, мы можем с правом то же самое предположить и относительно белков мяса, т. е. что и в мясе, помимо находящихся в нем с самого начала раздражающих веществ, часть их образуется также во время процесса переваривания мяса.
Представленный анализ знакомит нас с совершенно особенным и очень большим значением психического, или аппетитного, сока. При мясе этот сок бесспорно сильно поддерживает раздражителя, заключенного в самом мясе, и таким образом ведет к скорейшему перевариванию мяса, значительно сокращая время пребывания его в необработанном виде в пищеварительном канале. При других же сортах еды, как, например- хлеб, этот сок есть необходимое условие того, чтобы вообще началось переваривание пищи. И хлеб и яичный белок, съеденные без аппетита, как бы вложенные в желудок незаметно для вас, останутся лежать там на долгое время камнем, без всякого намека на переваривание. В этих случаях аппетитный сок есть единственное начало отделительного процесса и неизбежное условие продолжения его, так как только начавшееся под его влиянием переваривание этих веществ дальше может поддерживать само себя. Психический сок служит как бы спичкой для зажигания горючих материалов, ввиду чего доктор Хижин и дал ему название запального. Потому-то, вероятно, психический сок при всякой еде обладает однообразную, более или менее высокою, переваривающею силой.
Ясно, что в случае съедания яичного белка и хлеба без аппетита роль запала могли бы сыграть как чистая вода, так еще лучше навар мяса или раствор либиховского экстракта. Пользуясь всеми этими сведениями, мы как бы проверили на практике предшествующий анализ отделительного процесса. Читая эти лекции, демонстрируя на них наши старые опыты с перерезкой блуждающих нервов в отношении к секреторному эффекту мнимого кормления, я получил собаку глубоким расстройством пищеварительной деятельности желудка, о чем я знал как по личному опыту, так и по заявлению многих других авторов (особенно Людвиг и Крель). Я решил ухаживать за пищеварением животного, опираясь на новые факты. Так как у собак с перерезанными блуждающими нервами совершенно и навсегда отсутствует психическое отделение желудочного сока, то я старался заменить этот недостающий механизм искусственным. Промыв желудок собаки, мы вливали него несколько сот (200300) куб. сантиметров мясного навара и ждали того времени, когда навар становился сильнокислым, т. е. желудочные железы приходили в состояние значительной работа. Тогда только мы вкладывали плотную пищу. Благодаря этому мы достигли того, что пища, без этого загнивавшая, теперь удовлетворительно переваривалась.
Наш анализ отделительного процесса до сих пор имел в виду почти исключительно только количество сока, изливающегося на различные роды пищи. Из второй лекции мы знаем, что при разных сортах еды чрезвычайно разнится и качество сока. Как происходит это различие? Как уже упоминалось неоднократно, психический сок при всех родах пищи обладает более или менее однообразной, высокой переваривающей силой. Следовательно, различие в переваривающей силе сока при различных сортах еды в последующие часы отделительной работы должно исходить из особенностей химического раздражения со стороны той или другой пищи.
B этом отношении исследование прежде всего обратилось к факту чрезвычайно высокой переваривающей силы хлебного сока сравнительно с мясным. На чем бы могло основываться это различие двух соков? Возможен ряд предположений: могли иметь значение физическое состояние, затем особенности белковых веществ в мясе и хлебе и, наконец, комбинация в хлебе белка с крахмалом. Первое предположение легко устраняется: мясо можно подсушить, хлеб, наоборот, смочить, а отношение переваривающих сил сока остается одно и то же (д-р Хижин). Затем было испытано (д-ром Лобасовым) третье предположение. Мешая мясо с чистым вареным крахмалом, по возможности в тех отношениях, в каких стоят в хлебе белок и крахмал, и давая есть такой, можно сказать, искусственный хлеб собаке, мы получили как раз такой сок по переваривающей силе, какой действительно изливается на натуральный хлеб. Выставляю для сравнения таблицу (15) с переваривающей силой по часам, как и валовой, при натуральном и искусственном хлебе. 200 г хлеба. Опыт 25 мая 1894 г. (д-ра Хижина). Смесь из 100 г крахмала, 100 г мяса и 150 куб. см воды. Опыт 10 мая 1895 г. (д-ра Лобасова).
Совершенное оправдание третьего предположения исключало необходимость особой проверки второго. Хотя сам факт многократно повторялся и представляет, как мне кажется, ценное приобретение в этой области, внутренний механизм его, однако, требует дальнейшего расследования. Значение комбинаций крахмала с мясными белками может быть понимаемо в различных смыслах. Роль крахмала можно представлять себе так, что он, не затрагивая секреторной иннервации, сильно возбуждает трофическую. Но возможна и другая точка зрения. Мы знаем уже из второй лекции, что при мясной пище сок со второго часа делается все более и более слабым пищеварительном отношении, концентрируясь снова лишь к концу отделительного периода. Так как психический сок, имевший место в первом часе этой еды, всегда обладает более или менее высокой переваривающей силой, то понижение переваривающей силы со второго часа должно быть приписано действию химических раздражителей мяса. Раз это так, то крахмал, смешанный с мясом, мог бы как-нибудь мешать этому влиянию составных частей мяса, понижающему переваривающую силу. Хотя настоящий материал лаборатории недостаточен, чтобы придти по этому пункту к какому-нибудь определенному решению, но нельзя не считать успехом и то, что мы можем ставить такие вопросы и имеем возможность подойти к их решению экспериментальным путем.
Этот опыт интересен в том отношении, что дает довольно существенную опору допущению, молчаливо желаемому до сих пор, именно, что все испытанные до сих пор вещества действовали на нервную систему желез, раздражая слизистую оболочку желудка, а не через то, что всасывались в кровь и тогда уже раздражали или нервную систему желез, или самые железы непосредственно. Ясно, что, будь действие на железы через кровь, один раствор экстракта действовал бы гораздо сильнее, чем связанный крахмалом и затрудненный в отношение всасывания.
Особенно подробным исследованиям был подвергнут жир, как растительный, так и животный. Он испытывался на собаке с уединенным желудочком, на собаках с обыкновенной желудочной фистулой и перерезанным пищеводом, наконец, на собаке, пережившей многие месяцы перерезке блуждающего нерва на шее. Во всех опытах жир вводился прямо в желудок, т. е. при исключении акта еды. Результат всегда оставался отрицательным.
Итак, из всех пищевых веществ в широком смысле слова, при исследовании их врозь, большинство их оназалось без сокогонного эффекта в отношении желудка, меньшинство составили вода и неизвестные еще точно составные части мяса, переходящие в раствор.
От этих опытов с элементарными веществами перейдем к опытам вкладывания в желудок, присключении акта еды, различных сортов обыкновенной пищи и зададимся вопросом: насколько действие этих сложных веществ слагается и объясняется из элементарных вышеуказанных действий?
Если вложим в большой желудок нашей собаки с уединенным желудком, незаметно для нее, известное количество молотого сырого мяса, то через 15-30 минут (в лучшем случае) начинается отделение сока, как уже сказано в предшествующей лекции. Считаю нелишним упомянуть здесь о выработанном нами (д-р Лобасов) способе введения мяса. При вкладывании мяса кусочками собака часто догадывается, что с неи делается, и это, конечно, может повести к психическому отделению. Если бы собака и спала, то процедура вкладывания всегда ее будит и оканчивается при бодром состоянии животного. Для избежания этих неудобств мы накладываем молотое мясо в широкую стеклянную трубку, а затем, во время сна собаки, введя ее на небольшую глубину в фистульную трубку, при помощи поршня быстро вталкиваем мясо в желудок. Проснувшись, собака, однако, не может догадаться, что с ней было, так как все уже кончено, и сплошь и рядом сейчас же засыпает опять. Но и теперь мясо всегда вызывает себя отделение сока. После сообщенного в начале этой лекции факт этот, разумеется, не представляет ничего неожиданного и непонятного. Очевидно, раствор некоторых химических веществ, находящихся в мясном соке, есть главнейшая причина начинающегося теперь отделения. Доктор Лобасов, занимавшийся этим вопросом, поставил много видоизменений опыта с целью как можно более усилить это заключение. Он, например, сильно вываривал мясо в продолжение нескольких дней и, вкладывая его в желудок, убеждался, что сокогонное действие теперь или очень ослаблалось, или исчезало совершенно. Но стоило прибавить к такому мясу либиховского экстракта, для того чтобы действие, свойственное сырому мясу, восстановилось.
Совершенно такие же опыты со вкладыванием хлеба и вареного яичного белка, т. е. при полном исключении психического момента, как уже сказано в предшествующей лекции, дают совершенно отрицательный результат; эти сорта еды остаются в желудке 2 часа (до тех пор, пока за ними наблюдали) без малейшего возбуждающего действия на желудочные железы. Мы можем представить себе, что частью их Физико-химическое состояние (связанная вода), частью отсутствие в них непосредственно раздражающих химических веществ обусловливают этот неожиданный факт. Что касается до яичного белка, то стоит только припомнить результаты опытов с вливанием жидкого белка; а относительно хлеба доктор Лобасов нашел, что продолжительные настои его с водой едва ли действуют в качестве возбудителей желудочных желез энергичнее воды.
Можно ли удовлетвориться полученными результатами? Дают ли они нам полное разъяснение отделительного процесса при нормальной еде? Очевидно, нет. Если с едой мяса дело может считаться более или менее законченным - сок льется на него частью вследствие психического момента, частью вследствие собственного химически раздражающего действия, - то того же нельзя сказать об отделительной работе желудка при еде хлеба и вареного яичного белка. При них оказывается объясненным только первый период отделения, имеющий свое основание в психическом моменте; дальнейший же период после трех-четырех часов остается темным в своем механизме, -так как только часть выливающегося на них сока покрывается из известного нам источника. Для лучшего уяснения смысла занимающего нас вопроса я нахожу полезным предварительно остановить ваше внимание на сопоставлении опытов вкладыванием и с едой крахмала.
Как уже раньше сказано, чистый крахмал, вложенный прямо в желудок, не обусловливает сколько-нибудь значительного отделения, при еде же его животным наступает отделение в продолжение двух-трех часов. Внимательный разбор этих опытов скоро раскрывает весь их механизм. Отделяющийся при еде крахмала сок вполне покрывается психическим отделением, как оно установлено в опыте с мнимым кормлением. Этой точности итога для массы сока и нет в случае хлеба и вареного яичного белка; только 1 / или 1 /2 сока, на них изливающегося, может быть отнесена на психический момент, а источник остальной части предстоит отыскать. Что действительно имеется другой источник, другой раздражитель, в частности при еде белка, следует и из того, что сок второго и третьего часа обладает не особенно высокой переваривающей силой, между тем как психический сок, как уже замечено выше, по переваривающей силе принадлежит к сильнейшим сортам. Всестественнее допустить, что при изменении хлеба и вареного яичного белка посредством психического аппетитного сока в начале нормального пищеварения в них возникает нечто химическое, производящее раздражающее действие на нервноотделительный прибор желудка. Вероятно, это какие-нибудь продукты переваривания, тождественные или близкие к тем веществам, которые являются возбудителями в мясе.
В пользу такого понимания наблюдаемых фактов можно привести несколько экспериментальных данных. Если собирать жидкий продукт переваривания твердого яичного белка, скопляющийся в желудке собаки, и ввести его прямо в большой желудок нашей собаки с уединенным желудочком, то нельзя не заметить гораздо более верного и несколько более значительного сокогонного действия сравнительно с такою же массою воды или жидкого белка (д-р Лобасов). Во всяком случае образование этих веществ не может быть очень значительным, так как и яичный белок и хлеб при их поедании животным, за вычетом психического отделения, которое продолжается 2 3 часа, представляют вообще скудное отделение по часам, начиная с третьего-четвертого часа. Дальнейшим доказательством нашего толкования может служить следующий опыт. Если в желудке имеется уже отделение желудочного сока, вызванное или психически, или вообще предшествующим пищеварением, то введение в желудок (незаметно для животного) жидкого яичного белка обыкновенно сопровождается значительным взмахом отделения (д-р Хижин). Как понять это, если не допустить, что при начинающемся переваривании белка из него образуется нечто раздражающее слизистую оболочку? Приняв такое толкование явлений относительно хлеба и яичного белка, мы можем с правом то же самое предположить и относительно белков мяса, т. е. что и в мясе, помимо находящихся в нем с самого начала раздражающих веществ, часть их образуется также во время процесса переваривания мяса.
Представленный анализ знакомит нас с совершенно особенным и очень большим значением психического, или аппетитного, сока. При мясе этот сок бесспорно сильно поддерживает раздражителя, заключенного в самом мясе, и таким образом ведет к скорейшему перевариванию мяса, значительно сокращая время пребывания его в необработанном виде в пищеварительном канале. При других же сортах еды, как, например- хлеб, этот сок есть необходимое условие того, чтобы вообще началось переваривание пищи. И хлеб и яичный белок, съеденные без аппетита, как бы вложенные в желудок незаметно для вас, останутся лежать там на долгое время камнем, без всякого намека на переваривание. В этих случаях аппетитный сок есть единственное начало отделительного процесса и неизбежное условие продолжения его, так как только начавшееся под его влиянием переваривание этих веществ дальше может поддерживать само себя. Психический сок служит как бы спичкой для зажигания горючих материалов, ввиду чего доктор Хижин и дал ему название запального. Потому-то, вероятно, психический сок при всякой еде обладает однообразную, более или менее высокою, переваривающею силой.
Ясно, что в случае съедания яичного белка и хлеба без аппетита роль запала могли бы сыграть как чистая вода, так еще лучше навар мяса или раствор либиховского экстракта. Пользуясь всеми этими сведениями, мы как бы проверили на практике предшествующий анализ отделительного процесса. Читая эти лекции, демонстрируя на них наши старые опыты с перерезкой блуждающих нервов в отношении к секреторному эффекту мнимого кормления, я получил собаку глубоким расстройством пищеварительной деятельности желудка, о чем я знал как по личному опыту, так и по заявлению многих других авторов (особенно Людвиг и Крель). Я решил ухаживать за пищеварением животного, опираясь на новые факты. Так как у собак с перерезанными блуждающими нервами совершенно и навсегда отсутствует психическое отделение желудочного сока, то я старался заменить этот недостающий механизм искусственным. Промыв желудок собаки, мы вливали него несколько сот (200300) куб. сантиметров мясного навара и ждали того времени, когда навар становился сильнокислым, т. е. желудочные железы приходили в состояние значительной работа. Тогда только мы вкладывали плотную пищу. Благодаря этому мы достигли того, что пища, без этого загнивавшая, теперь удовлетворительно переваривалась.
Наш анализ отделительного процесса до сих пор имел в виду почти исключительно только количество сока, изливающегося на различные роды пищи. Из второй лекции мы знаем, что при разных сортах еды чрезвычайно разнится и качество сока. Как происходит это различие? Как уже упоминалось неоднократно, психический сок при всех родах пищи обладает более или менее однообразной, высокой переваривающей силой. Следовательно, различие в переваривающей силе сока при различных сортах еды в последующие часы отделительной работы должно исходить из особенностей химического раздражения со стороны той или другой пищи.
B этом отношении исследование прежде всего обратилось к факту чрезвычайно высокой переваривающей силы хлебного сока сравнительно с мясным. На чем бы могло основываться это различие двух соков? Возможен ряд предположений: могли иметь значение физическое состояние, затем особенности белковых веществ в мясе и хлебе и, наконец, комбинация в хлебе белка с крахмалом. Первое предположение легко устраняется: мясо можно подсушить, хлеб, наоборот, смочить, а отношение переваривающих сил сока остается одно и то же (д-р Хижин). Затем было испытано (д-ром Лобасовым) третье предположение. Мешая мясо с чистым вареным крахмалом, по возможности в тех отношениях, в каких стоят в хлебе белок и крахмал, и давая есть такой, можно сказать, искусственный хлеб собаке, мы получили как раз такой сок по переваривающей силе, какой действительно изливается на натуральный хлеб. Выставляю для сравнения таблицу (15) с переваривающей силой по часам, как и валовой, при натуральном и искусственном хлебе. 200 г хлеба. Опыт 25 мая 1894 г. (д-ра Хижина). Смесь из 100 г крахмала, 100 г мяса и 150 куб. см воды. Опыт 10 мая 1895 г. (д-ра Лобасова).
Совершенное оправдание третьего предположения исключало необходимость особой проверки второго. Хотя сам факт многократно повторялся и представляет, как мне кажется, ценное приобретение в этой области, внутренний механизм его, однако, требует дальнейшего расследования. Значение комбинаций крахмала с мясными белками может быть понимаемо в различных смыслах. Роль крахмала можно представлять себе так, что он, не затрагивая секреторной иннервации, сильно возбуждает трофическую. Но возможна и другая точка зрения. Мы знаем уже из второй лекции, что при мясной пище сок со второго часа делается все более и более слабым пищеварительном отношении, концентрируясь снова лишь к концу отделительного периода. Так как психический сок, имевший место в первом часе этой еды, всегда обладает более или менее высокой переваривающей силой, то понижение переваривающей силы со второго часа должно быть приписано действию химических раздражителей мяса. Раз это так, то крахмал, смешанный с мясом, мог бы как-нибудь мешать этому влиянию составных частей мяса, понижающему переваривающую силу. Хотя настоящий материал лаборатории недостаточен, чтобы придти по этому пункту к какому-нибудь определенному решению, но нельзя не считать успехом и то, что мы можем ставить такие вопросы и имеем возможность подойти к их решению экспериментальным путем.
 Как бы то ни было, выдвинут новый факт, что вещество, само по себе не возбуждающее отделения желудочного сока, в комбинации с раздражающими веществами мяса, резко и своеобразно изменяет работу желез.
С вопросом о значении крахмала для отделительной работы желудка в естественной связи по многим отношениям стоит вопрос о таком же значении жира.
Как и при крахмале, отрицательные опыты относительно возбуждающего действия жира на желудочные железы далеко не исчерпывают всей его роли в том случае, когда он появляется в пищеварительном канале предшественником или спутником других веществ. При тщательном изучении действия жира выступила совершенно новая и важная сторона в отделительном процессе, как исследование комбинации крахмала с мясом принесло нам интересный факт влияния крахмала на качество сока.
Если ввести собаке зондом или влить в фистулу большого желудка, например, 100 г прованского масла (причем, как вы уже знаете, никакого отделения не наступает), а затем, спустя полчаса-час дать собаке съесть обычную порцию 400 г мяса, то получается совершенно новая картина отделительной работы желудка, чем если бы дать ту же еду без предварительного жира (д-р Хижин). Начала отделения теперь вместо обычных 5 10 минут приходится ждать полчаса-час и больше. Начавшееся, наконец, отделение весьма слабо, часа вы получаете для часовой величины 3-5 куб. см вместо 10-15 куб. см, и лишь позже перед вами идут количества сока, отвечающие нормальным. Подобное же искажение кривой нормального отделения при мясе наблюдается в том случае, если вы вводите жир прямо после еды мяса; разница будет только в том, что теперь отделение может начаться в свое время после еды (5--10 минут) и с нормальною энергией, и лишь через 10-15 минут разыграется прежняя история, т. е. задержка нормального отделения. Наконец совершенно то же самое оказывается и тогда, когда собаке дают мясо, перемешанное с жиром. Во всех втих случаях (опыты д-ра Лобасова) с уменьшением количества сока часто наблюдается и уменьшение его переваривающей силы. Представляю в таблицах (16 и 17) один пример из этих опытов, причем для резкости сопоставляю нормальное отделение при мясе с измененным под влиянием жира.
Как бы то ни было, выдвинут новый факт, что вещество, само по себе не возбуждающее отделения желудочного сока, в комбинации с раздражающими веществами мяса, резко и своеобразно изменяет работу желез.
С вопросом о значении крахмала для отделительной работы желудка в естественной связи по многим отношениям стоит вопрос о таком же значении жира.
Как и при крахмале, отрицательные опыты относительно возбуждающего действия жира на желудочные железы далеко не исчерпывают всей его роли в том случае, когда он появляется в пищеварительном канале предшественником или спутником других веществ. При тщательном изучении действия жира выступила совершенно новая и важная сторона в отделительном процессе, как исследование комбинации крахмала с мясом принесло нам интересный факт влияния крахмала на качество сока.
Если ввести собаке зондом или влить в фистулу большого желудка, например, 100 г прованского масла (причем, как вы уже знаете, никакого отделения не наступает), а затем, спустя полчаса-час дать собаке съесть обычную порцию 400 г мяса, то получается совершенно новая картина отделительной работы желудка, чем если бы дать ту же еду без предварительного жира (д-р Хижин). Начала отделения теперь вместо обычных 5 10 минут приходится ждать полчаса-час и больше. Начавшееся, наконец, отделение весьма слабо, часа вы получаете для часовой величины 3-5 куб. см вместо 10-15 куб. см, и лишь позже перед вами идут количества сока, отвечающие нормальным. Подобное же искажение кривой нормального отделения при мясе наблюдается в том случае, если вы вводите жир прямо после еды мяса; разница будет только в том, что теперь отделение может начаться в свое время после еды (5--10 минут) и с нормальною энергией, и лишь через 10-15 минут разыграется прежняя история, т. е. задержка нормального отделения. Наконец совершенно то же самое оказывается и тогда, когда собаке дают мясо, перемешанное с жиром. Во всех втих случаях (опыты д-ра Лобасова) с уменьшением количества сока часто наблюдается и уменьшение его переваривающей силы. Представляю в таблицах (16 и 17) один пример из этих опытов, причем для резкости сопоставляю нормальное отделение при мясе с измененным под влиянием жира.
 Перед нами новый и в высшей степени резкий факт: жир подавляет, тормозит нормальную энергию отделительного процесса. Как толковать его? Принимая во внимание обстановку наших опытов, т. е. наблюдение отделения уединенном желудочке, можно объяснять его на два лада. Мир задерживает отделение или грубо механически тем, чтобволакивая слизистую оболочку большого желудка, мешает раздражению ее, или тем, что рефлекторно обусловливает торможение отделительной работы, вследствие ли раздражения задерживающих нервов желез или вследствие задерживания центров секреторных нервов. Строго оценивая факты, мы останавливаемся главным образом на втором механизме действия жира. Уже доказано выше, отделительная работа при еде мяса начинается непрерывно с психического отделения, т. е. с центрального, а оно-то и задерживается прежде всего жиром, как вы отчетливо видели на предъявленном опыте.
Ввиду большого значения задерживающего влияния жира мы (д-р Лобасов) старались всячески разнообразить опыты. У собак с гастро- и эзофаготомией производилось мнимое кормление короткого продолжения, например одноминутное, и замечались точно как время наступления отделения, так количество и качество выливавшегося сока. Затем у тех же собак вливали в желудок 50-100 куб. см жира и, спустя 44--часа и более, вновь устраивали мнимое кормление совершенно тех же размеров как относительно продолжительности, так и количества скормленной еды. При этом иногда перед самым мнимым кормлением выливали жир из желудка, в другой же раз оставляли его там и при мнимом кормлении, наблюдая отделение сока в толстой стеклянной трубке, вправленной в желудочную фистулу. Понятно, что сок, как более тяжелый сравнительно с маслом, должен был опускаться на дно нашей трубки. Без исключения во всех случаях наблюдалось резкое ослабление психического отделения: сока иногда не бывало совсем; когда был, то отделение его начиналось гораздо позже, в резко меньшем количестве и с сильно уменьшенной переваривающей силой. Особенно убедительным опыт с задерживающим действием жира становится на собаке с уединенным желудочком и с эзофаготомией (см. табл. 18).
Влито в желудок 100 г прованского масла. Через 30 минут мнимое кормление в продолжение 6 минут. За 2 часа из уединенного желудочка ничего не выделилось. Снова мнимое кормление в продолжение 6 минут. За час после него собрано 1.8 куб. см сока с переваривающей силой 4.0 мм.
Очень интересно, что мнимое кормление, продолжающееся долго, совершенно пересиливает задерживающее влияние жира.
Если даже вообще сильный раздражитель, психический момент, значительно ослабляется со стороны жира, то тем более это можно допустить относительно раздражителей, действующих на слизистую оболочку желудка. Остается ли что-нибудь на долю обволакивания слизистой оболочки жирным слоем в смысле задерживания от длительной работы с этой стороны, сказать определенно на основании нашего материала мы не можем.
Мнимое кормление в продолжение 6 минут (табл. 18).
Разъясненное действие жира может, кажется, до известной степени помочь нам понять картину отделительной работы при молоке, разумея при этом вялый ход отделения и сравнительно низкую переваривающую силу молочного сока. самом деле не жир ли молока тому причиной? Мы думали получить для решения этого вопроса некоторый экспериментальный материал, проделав опыт на сливках, т. е. на молоке с большим содержанием жира. Если жир вообще причинизкой переваривающей силы молочного сока, то сок на сливки должен оказаться еще слабее по пищеварительной силе. Это в действительности и оказалось. Опять сопоставляю отделение при молоке и сливках (опыт д-ра Лобасова) (табл. 19).
Перед нами новый и в высшей степени резкий факт: жир подавляет, тормозит нормальную энергию отделительного процесса. Как толковать его? Принимая во внимание обстановку наших опытов, т. е. наблюдение отделения уединенном желудочке, можно объяснять его на два лада. Мир задерживает отделение или грубо механически тем, чтобволакивая слизистую оболочку большого желудка, мешает раздражению ее, или тем, что рефлекторно обусловливает торможение отделительной работы, вследствие ли раздражения задерживающих нервов желез или вследствие задерживания центров секреторных нервов. Строго оценивая факты, мы останавливаемся главным образом на втором механизме действия жира. Уже доказано выше, отделительная работа при еде мяса начинается непрерывно с психического отделения, т. е. с центрального, а оно-то и задерживается прежде всего жиром, как вы отчетливо видели на предъявленном опыте.
Ввиду большого значения задерживающего влияния жира мы (д-р Лобасов) старались всячески разнообразить опыты. У собак с гастро- и эзофаготомией производилось мнимое кормление короткого продолжения, например одноминутное, и замечались точно как время наступления отделения, так количество и качество выливавшегося сока. Затем у тех же собак вливали в желудок 50-100 куб. см жира и, спустя 44--часа и более, вновь устраивали мнимое кормление совершенно тех же размеров как относительно продолжительности, так и количества скормленной еды. При этом иногда перед самым мнимым кормлением выливали жир из желудка, в другой же раз оставляли его там и при мнимом кормлении, наблюдая отделение сока в толстой стеклянной трубке, вправленной в желудочную фистулу. Понятно, что сок, как более тяжелый сравнительно с маслом, должен был опускаться на дно нашей трубки. Без исключения во всех случаях наблюдалось резкое ослабление психического отделения: сока иногда не бывало совсем; когда был, то отделение его начиналось гораздо позже, в резко меньшем количестве и с сильно уменьшенной переваривающей силой. Особенно убедительным опыт с задерживающим действием жира становится на собаке с уединенным желудочком и с эзофаготомией (см. табл. 18).
Влито в желудок 100 г прованского масла. Через 30 минут мнимое кормление в продолжение 6 минут. За 2 часа из уединенного желудочка ничего не выделилось. Снова мнимое кормление в продолжение 6 минут. За час после него собрано 1.8 куб. см сока с переваривающей силой 4.0 мм.
Очень интересно, что мнимое кормление, продолжающееся долго, совершенно пересиливает задерживающее влияние жира.
Если даже вообще сильный раздражитель, психический момент, значительно ослабляется со стороны жира, то тем более это можно допустить относительно раздражителей, действующих на слизистую оболочку желудка. Остается ли что-нибудь на долю обволакивания слизистой оболочки жирным слоем в смысле задерживания от длительной работы с этой стороны, сказать определенно на основании нашего материала мы не можем.
Мнимое кормление в продолжение 6 минут (табл. 18).
Разъясненное действие жира может, кажется, до известной степени помочь нам понять картину отделительной работы при молоке, разумея при этом вялый ход отделения и сравнительно низкую переваривающую силу молочного сока. самом деле не жир ли молока тому причиной? Мы думали получить для решения этого вопроса некоторый экспериментальный материал, проделав опыт на сливках, т. е. на молоке с большим содержанием жира. Если жир вообще причинизкой переваривающей силы молочного сока, то сок на сливки должен оказаться еще слабее по пищеварительной силе. Это в действительности и оказалось. Опять сопоставляю отделение при молоке и сливках (опыт д-ра Лобасова) (табл. 19).

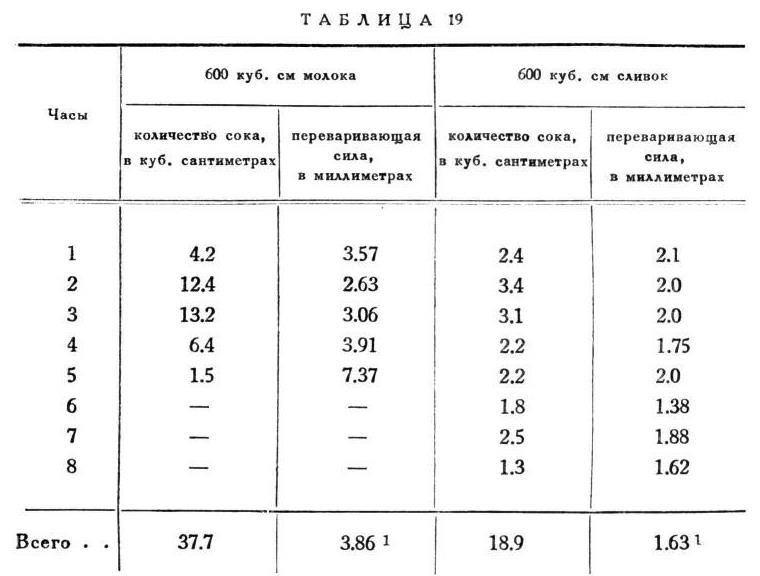 Приятно видеть успехи анализа отделительного процесса: выше мы объяснили себе первые пункты кривой молочного отделения (небольшого отделения первого часа вследствие отсутствия психического раздражения), теперь приближаемся к пониманию качества молочного сока.
В настоящем пункте моих лекций я нахожу вполне удобным заняться, наконец, решением двух важнейших для нас вопросов, требовавших уже давно своего ответа: один со времени первой лекции и другой с начала текущей лекции. Первый вопрос вопрос о праве нашего маленького уединенного желудка представлять собою во всех случаях от длительной работы большой желудок, служить как бы его зеркалом (как выразился доктор Хижин в своей диссертации). Второй вопрос состоит в том, можно ли различные вещества, возбуждающие отделение желудочного сока или как-нибудь его варьирующие, считать действующими именно на слизистую оболочку желудка, возбуждая находящиеся в ней периферические окончания центростремительных нервов нервноотделительного прибора желудка. Эти вопросы тесно связаны друг с другом, и потому разрешение их приурочено мною к одному месту. Начну с первого.
Всякому, знакомящемуся впервые с нашими опытами над отделительной деятельностью желудка, бросается в глаза, повидимому, весьма существенное обстоятельство. В то время как большой желудок обычным порядком при нормальной еде наполняется пищей, наш маленький всегда остается свободным от нес. Казалось бы, что прикосновение пищи в одном случае и отсутствие этого в другом должны образовать огромное различие в условиях работы того и другого. Однако при тщательном анализе всех отношений, с кучею фактов в руках, можно решительно заявить, что это, повидимому, важное обстоятельство - в сушности безразлично. Когда при начале еды сок течет из нашего желудочка, то деятельность его в этот момент должна быть признана совершенно тождественной с деятельностью большого желудка. После всего сообщенного в этой и двух предыдущих лекциях такое утверждение понятно само собой: дело начинается с психического раздражения центров секреторных нервов, и это раздражение, конечно, одинаково проводится ко всем пунктам слизистой оболочки с ее железами как большого, так и маленького желудочка. А раз доказано это, мы уже должны быть расположены и в дальнейших фазах отделительного периода, ради единства объяснения, видеть деятельность нервной системы. Положение дела сейчас коренным образом изменилось против раннего, когда, несмотря на усиленные старания, иннервация желудочных желез никак не хотела даваться исследователям. Теперь она есть, сложная, и должна быть занята работой. Если начало отделительного процесса тождественно в обоих желудках, то как стоит дело затем, когда разыгрывается отделение, имеющее в основании своем, по нашему предположению, местное раздражающее действие пищи, или, выражаясь объективнее, просто в дальнейший период отделительного процесса, когда израсходуется действие психического раздражения? Во всяком случае мы имеем перед собою факт, что отделение в маленьком желудочке существует и тогда, когда психическое отделение закончилось или даже и совсем не существовало, как, например, при вкладывании мяса в большой желудок незаметно для собаки. Естественно, этот факт должен лечь в основание всего нашего рассуждения. Как он может существовать? Каким образом местные явления в большом желудке могут отразиться на маленьком желудке? Эту фактическую связь между обоими желудками можно представить себе только на два лада, посредством двух известных связующих, объединяющих систем организма - кровеносной и нервной.
Можно представить себе, что химические вещества, вызывающие отделение, всасываясь в пищеварительном канале, с кровью приносятся или к центрам отделительных нервов, или к самим пепсиновым железам, раздражая или то, или другое. Такое предположение доступно легкой проверке. Если предположение верно, то при введении этих веществ в кровеносную систему каким-нибудь другим путем, а не из желудка, мы должна получить ту же работу желез. Опыт решительно оказывался потив этого предположения. Многие авторы вводили прямую кишку навары мяса и растворы либиховского экстракта, но никогда ни малейшего проявления деятельности желез не видали. Особенно настойчиво разрабатывал этот пункт доктор Лобасов, вводивший в прямую кишку во много раз большие дозы либиховского экстракта, чем какие вводились в желудок для возбуждения сока. Последовательным вымыванием rectum и анализом химическим и физиологическим он убеждался в исчезании из rectum возбуждающих желудочное отделение веществ либиховского экстракта. Таким образом путем исключения мы приходим к необходимому заключению, что наш маленький желудок и в позднейших фазах отделительного процесса возбуждается со стороны большого желудка нервным путем, путем рефлекса, Следовательно, заключаем мы дальше, и в самом большом желудке отделение существует также на основании рефлекторного раздражения. Раз это - рефлекс, то легко понятно, ввиду условий переваривания пищи в желудке, что рефлекс этот должен быть разлитьм, а не локализированным, т. е. раздражение данной точки слизистой оболочки не обусловливает отделения именно в той же точке слизистой оболочки, а вообще ведет к работе безразлично всех желудочных желез. Первое в самом деле не имело бы никакого смысла, так как пища в желудке находится в постоянном движении, переходя с одного пункта на другие. Поэтому вполне естественно, что раздражение поверхности большого желудка постоянно и точно передается по нервам на вырезанный из него кусок в виде нашего уединенного желудочка, раз только нервные отношения нашего куска остались неприкосновенными. Последнее заключение очень приобретает в силе, когда мы сравниваем деятельность уединенных желудочков по нашему способу (с сохранением блуждающих нервов) и по гейденгайновскому (при перерезке этих нервов). Собака, оперированная по нашему способу, теперь уже два с полавиною года тому назад, неизменно представляет в уединенном желудочке один и тот же ход отделительной работы, при определенных условиях. Желудочки же, уединенные по способу Гейденгайна, резко меняют с течением времени характер своей отделительной работы. Вначале их работа очень энергична, отделение при обильной еде продолжается много часов, в большом размере (Гейденгайн и Саноцкий). Но если животное живет больше, то замечается постепенное ограничение работы этого желудочка, и спустя один-полтора месяца после операции даже на очень большую еду отделение наблюдается лишь 3-5 часов, делаясь все скуднее и скуднее, что касается часовой порции.
Все сделанные выше выводы не есть, однако, единственное основание для убеждения в правильности нашей методики. Полная параллельность работы большого и маленького желудков в настоящее время доказана для нас прямыми наблюдениями, т. е. имеется полное оправдание тех выводов. Нам нужно только припомнить и собрать воедино все относящиеся сюда факты. В пятой лекции был описан и показана цифрах опыт на нашей собаке с уединенным желудочком и эзофаготомией при мнимом кормлении. Как помните, отделение в обоих желудках, при этих условиях, было совершенно одинаково во всех пунктах. Отсутст•вие эффекта мнимого кормления на собаках с уединенным желудочком по способу Гейденгайна (т. е. с перерезанными блуждающими нервами) вполне совпадает с бесплодностью мнимого кормления у собак с цельным желудком и с перерезанными блуждающими нервами. То же сходство в деятельности обоих желудочков наблюдается и при раздражителях, действующих из желудка. Вода возбуждает отделение как в большом, так и в малом. То же надо сказать и относительно растворов либиховского экстракта, причем одинаково на обоих желудках замечается, что последний раздражитель гораздо энергичнее первого. Яичной белок и крахмал как в жидком виде, так и в форме кусков, оставляют в покое и тот и другой желудки. Жир также в обоих желудках не только не дает отделения, но развивает задерживающее действие. Короче сказать, мы не знаем ни одного случая, где бы маленький и большой желудки расходились в их секреторной деятельности. Считаю важным тут же прибавить, что многие факты, собранные нами на нашей собаке с двумя желудочками, проделаны врозь и подтверждены на многих и различных собаках с простыми желудочными фистулами и эзофаготомией. В самое последнее время другая собака, оперированная по нашему способу изоляции части желудка, стереотипно воспроизводит все главнейшие факты, собранные на первой нашей собаке.
Нетрудно заметить, что сопоставленными фактами решается и наш второй вопрос - о пункте действия веществ, обусловли- ивающих отделение из желудка. Если доказано теперь, что вся отделительная работа желудка - нервная и, исключая период психического возбуждения, - рефлекторная, то этим прочно устанавливается, что раздражитель прикладывается к плоскости периферических окончаний центростремительных нервов, т. е. к слизистой оболочке пищеварительного канала, вероятно, желудка. Помимо отсутствия действия при введении веществ в rectum, за это может быть приведен и следующий факт, сообщенный доктором Фремоном. При перерезке блуждающих нервов целый изолированный желудок остается в полном отделительном покое, несмотря на поступление пищи в кишечный канал через рот. Так как при изоляции только куска желудка по способу Гейденгайна, т. е. с перерезкой блуждающих нервов этого куска, отделение в нем все же существует, лишь только пища находится в остальном желудке, то для отрицательного факта Фремона мыслимо только одно основание - полное отсутствие раздражения именно слизистой оболочки желудка. Конечно, остается еще крайняя возможность, что, помимо рефлекторного раздражения, пища оказывает и какое-то ближайшее, более непосредственное действие на желудочные железы. Нужно признаться, что это предположение, бывшее в большом ходу прежде, при отсутствии элементов иннервации желудочных желез, так сказать, предположение поневоле, сейчас представляется даже трудно мыслимым. Надо допустить какое-то пропитывание (совсем уже не физиологический акт!) толщи слизистой оболочки пищевыми веществами для того, чтобы они могли раздражать желудочные железы. В настоящее время это предположение тем невероятнее, что последние эксперименты очень ограничивают всасывающую способность внутренней поверхности желудка, а при этом надо еще иметь в виду, что, будь всасывание, оно все же не ведет непосредственно к прониканию веществ в пепсиновые железы. Допустить проникакие веществ через отверстие желез - так же нелегко, так как во время отделительного периода ток жидкости непрерывно направляется из полости желез полость желудка. Как подтверждение разбираемого нами предположения приводилась аналогия с насекомоядными растениями, но аналогия едва ли законная. У растений нервная система еще не выделена в особую ткань и принцип, функция ее распределена, разлита по всем клеткам; в нашем же случае, при желудочных железах, мы располагаем деятельностью чрезвычайно сложного нервного прибора. К сожалению, это невероятное предположение, сколько я могу сообразить, нельзя окончательно похоронить каким-нибудь прямым опытом, оно позабудется лишь постепенно, уступая место изучению нервно-железистых явлений, которые, наверное, будут привлекать к себе все большее и большее внимание.
Доказав нервный характер всего отделительного периода при желудочных железах, я должен весь предшествующий материал представить вам еще раз как картину работы иннервационного прибора этих желез. В огромном большинстве случаев при желудочном пищеварении дело начинается с сильного центрального (автоматического) раздражения как секреторных, так и трофических волокон желудочных желез. Спустя более или менее продолжительное время после акта еды, начинают давать себя знать рефлекторные раздражители в желудке при постепенном угасании автоматического, психического раздражителя. Если съедено мясо, то центр секреторных волокон продолжает сильно раздражаться и рефлекторно со стороны желудка экстрактивными веществами мяса, между тем как центры трофических волокон, с соответственных периферических окончаний, получают только слабые толчки. В случаях хлеба совершенно наоборот: после автоматического раздражения центры секреторных волокон слабо возбуждаются со стороны периферических окончаний, но зато продолжают энергично раздражаться центры трофических со своих окончаний. В случае примеси жира к пище в центры посылаются рефлекторно-задерживающие влияния, ограничивающие деятельность как секреторных, так и трофических волокон.
Я описал железистую работу желудка такою, какою мы ее видели в наших опытах, какою она выходила из-под наших рук. Нова ли эта картина? В подробностях - да, но не в основных ее чертах. Как это ни странно, общий ее очерк имелся в науке уже целых 50 лет тому назад. Лишний повод боящимся новизны примкнуть к нашим взглядам.
У талантливого автора «Traite analytique de la digestion» (1843), Блондло, совершенно ясно говорится как о значении акта еды, так и о специфической раздражимости слизистой оболочки желудка. Конечно, фактическая разработка темы далеко не достаточна, но не надо забывать, что это были первые опыты с искусственным свищом у собаки. Поистине непостижимо, каким образом опыты Блондло и его воззрения на желудочную деятельность в продолжение протекших пятидесяти лет не только не умножились, не развились, а скорее пришли забвение благодаря ошибочным опытам и грубым взглядам последующих авторов. Отзвук труда Блондло только изредка чувствуется в физиологических сочинениях французских авторов. Из других авторов должно указать еще на Гейденгайна, так обогатившего вообще физиологию отделения и, в частности, относительно желудка сообщившего несколько верных фактов и пустившего в обращение несколько плодотворных идей. От него вышли некоторые факты и идея о расчленении отделительного процесса по периодам и по раздражителям и мысль о важном значении исследований различных сортов еды, взятых врозь, на отделительную работу желудка. [25] Книга Блондло и статья Гейденгайна, собственно говоря, исчерпывают все существенное, что имеется в физиологии за 50 с лишком лет относительно условий и механизма отделительной работы желудка во время пищеварения. Роковым для нашего вопроса, очевидно, оказался научный промах с мнимою действительностью механического момента как возбудителя желудочных желез, а он в свою очередь был грехом недостаточной методики.
Приятно видеть успехи анализа отделительного процесса: выше мы объяснили себе первые пункты кривой молочного отделения (небольшого отделения первого часа вследствие отсутствия психического раздражения), теперь приближаемся к пониманию качества молочного сока.
В настоящем пункте моих лекций я нахожу вполне удобным заняться, наконец, решением двух важнейших для нас вопросов, требовавших уже давно своего ответа: один со времени первой лекции и другой с начала текущей лекции. Первый вопрос вопрос о праве нашего маленького уединенного желудка представлять собою во всех случаях от длительной работы большой желудок, служить как бы его зеркалом (как выразился доктор Хижин в своей диссертации). Второй вопрос состоит в том, можно ли различные вещества, возбуждающие отделение желудочного сока или как-нибудь его варьирующие, считать действующими именно на слизистую оболочку желудка, возбуждая находящиеся в ней периферические окончания центростремительных нервов нервноотделительного прибора желудка. Эти вопросы тесно связаны друг с другом, и потому разрешение их приурочено мною к одному месту. Начну с первого.
Всякому, знакомящемуся впервые с нашими опытами над отделительной деятельностью желудка, бросается в глаза, повидимому, весьма существенное обстоятельство. В то время как большой желудок обычным порядком при нормальной еде наполняется пищей, наш маленький всегда остается свободным от нес. Казалось бы, что прикосновение пищи в одном случае и отсутствие этого в другом должны образовать огромное различие в условиях работы того и другого. Однако при тщательном анализе всех отношений, с кучею фактов в руках, можно решительно заявить, что это, повидимому, важное обстоятельство - в сушности безразлично. Когда при начале еды сок течет из нашего желудочка, то деятельность его в этот момент должна быть признана совершенно тождественной с деятельностью большого желудка. После всего сообщенного в этой и двух предыдущих лекциях такое утверждение понятно само собой: дело начинается с психического раздражения центров секреторных нервов, и это раздражение, конечно, одинаково проводится ко всем пунктам слизистой оболочки с ее железами как большого, так и маленького желудочка. А раз доказано это, мы уже должны быть расположены и в дальнейших фазах отделительного периода, ради единства объяснения, видеть деятельность нервной системы. Положение дела сейчас коренным образом изменилось против раннего, когда, несмотря на усиленные старания, иннервация желудочных желез никак не хотела даваться исследователям. Теперь она есть, сложная, и должна быть занята работой. Если начало отделительного процесса тождественно в обоих желудках, то как стоит дело затем, когда разыгрывается отделение, имеющее в основании своем, по нашему предположению, местное раздражающее действие пищи, или, выражаясь объективнее, просто в дальнейший период отделительного процесса, когда израсходуется действие психического раздражения? Во всяком случае мы имеем перед собою факт, что отделение в маленьком желудочке существует и тогда, когда психическое отделение закончилось или даже и совсем не существовало, как, например, при вкладывании мяса в большой желудок незаметно для собаки. Естественно, этот факт должен лечь в основание всего нашего рассуждения. Как он может существовать? Каким образом местные явления в большом желудке могут отразиться на маленьком желудке? Эту фактическую связь между обоими желудками можно представить себе только на два лада, посредством двух известных связующих, объединяющих систем организма - кровеносной и нервной.
Можно представить себе, что химические вещества, вызывающие отделение, всасываясь в пищеварительном канале, с кровью приносятся или к центрам отделительных нервов, или к самим пепсиновым железам, раздражая или то, или другое. Такое предположение доступно легкой проверке. Если предположение верно, то при введении этих веществ в кровеносную систему каким-нибудь другим путем, а не из желудка, мы должна получить ту же работу желез. Опыт решительно оказывался потив этого предположения. Многие авторы вводили прямую кишку навары мяса и растворы либиховского экстракта, но никогда ни малейшего проявления деятельности желез не видали. Особенно настойчиво разрабатывал этот пункт доктор Лобасов, вводивший в прямую кишку во много раз большие дозы либиховского экстракта, чем какие вводились в желудок для возбуждения сока. Последовательным вымыванием rectum и анализом химическим и физиологическим он убеждался в исчезании из rectum возбуждающих желудочное отделение веществ либиховского экстракта. Таким образом путем исключения мы приходим к необходимому заключению, что наш маленький желудок и в позднейших фазах отделительного процесса возбуждается со стороны большого желудка нервным путем, путем рефлекса, Следовательно, заключаем мы дальше, и в самом большом желудке отделение существует также на основании рефлекторного раздражения. Раз это - рефлекс, то легко понятно, ввиду условий переваривания пищи в желудке, что рефлекс этот должен быть разлитьм, а не локализированным, т. е. раздражение данной точки слизистой оболочки не обусловливает отделения именно в той же точке слизистой оболочки, а вообще ведет к работе безразлично всех желудочных желез. Первое в самом деле не имело бы никакого смысла, так как пища в желудке находится в постоянном движении, переходя с одного пункта на другие. Поэтому вполне естественно, что раздражение поверхности большого желудка постоянно и точно передается по нервам на вырезанный из него кусок в виде нашего уединенного желудочка, раз только нервные отношения нашего куска остались неприкосновенными. Последнее заключение очень приобретает в силе, когда мы сравниваем деятельность уединенных желудочков по нашему способу (с сохранением блуждающих нервов) и по гейденгайновскому (при перерезке этих нервов). Собака, оперированная по нашему способу, теперь уже два с полавиною года тому назад, неизменно представляет в уединенном желудочке один и тот же ход отделительной работы, при определенных условиях. Желудочки же, уединенные по способу Гейденгайна, резко меняют с течением времени характер своей отделительной работы. Вначале их работа очень энергична, отделение при обильной еде продолжается много часов, в большом размере (Гейденгайн и Саноцкий). Но если животное живет больше, то замечается постепенное ограничение работы этого желудочка, и спустя один-полтора месяца после операции даже на очень большую еду отделение наблюдается лишь 3-5 часов, делаясь все скуднее и скуднее, что касается часовой порции.
Все сделанные выше выводы не есть, однако, единственное основание для убеждения в правильности нашей методики. Полная параллельность работы большого и маленького желудков в настоящее время доказана для нас прямыми наблюдениями, т. е. имеется полное оправдание тех выводов. Нам нужно только припомнить и собрать воедино все относящиеся сюда факты. В пятой лекции был описан и показана цифрах опыт на нашей собаке с уединенным желудочком и эзофаготомией при мнимом кормлении. Как помните, отделение в обоих желудках, при этих условиях, было совершенно одинаково во всех пунктах. Отсутст•вие эффекта мнимого кормления на собаках с уединенным желудочком по способу Гейденгайна (т. е. с перерезанными блуждающими нервами) вполне совпадает с бесплодностью мнимого кормления у собак с цельным желудком и с перерезанными блуждающими нервами. То же сходство в деятельности обоих желудочков наблюдается и при раздражителях, действующих из желудка. Вода возбуждает отделение как в большом, так и в малом. То же надо сказать и относительно растворов либиховского экстракта, причем одинаково на обоих желудках замечается, что последний раздражитель гораздо энергичнее первого. Яичной белок и крахмал как в жидком виде, так и в форме кусков, оставляют в покое и тот и другой желудки. Жир также в обоих желудках не только не дает отделения, но развивает задерживающее действие. Короче сказать, мы не знаем ни одного случая, где бы маленький и большой желудки расходились в их секреторной деятельности. Считаю важным тут же прибавить, что многие факты, собранные нами на нашей собаке с двумя желудочками, проделаны врозь и подтверждены на многих и различных собаках с простыми желудочными фистулами и эзофаготомией. В самое последнее время другая собака, оперированная по нашему способу изоляции части желудка, стереотипно воспроизводит все главнейшие факты, собранные на первой нашей собаке.
Нетрудно заметить, что сопоставленными фактами решается и наш второй вопрос - о пункте действия веществ, обусловли- ивающих отделение из желудка. Если доказано теперь, что вся отделительная работа желудка - нервная и, исключая период психического возбуждения, - рефлекторная, то этим прочно устанавливается, что раздражитель прикладывается к плоскости периферических окончаний центростремительных нервов, т. е. к слизистой оболочке пищеварительного канала, вероятно, желудка. Помимо отсутствия действия при введении веществ в rectum, за это может быть приведен и следующий факт, сообщенный доктором Фремоном. При перерезке блуждающих нервов целый изолированный желудок остается в полном отделительном покое, несмотря на поступление пищи в кишечный канал через рот. Так как при изоляции только куска желудка по способу Гейденгайна, т. е. с перерезкой блуждающих нервов этого куска, отделение в нем все же существует, лишь только пища находится в остальном желудке, то для отрицательного факта Фремона мыслимо только одно основание - полное отсутствие раздражения именно слизистой оболочки желудка. Конечно, остается еще крайняя возможность, что, помимо рефлекторного раздражения, пища оказывает и какое-то ближайшее, более непосредственное действие на желудочные железы. Нужно признаться, что это предположение, бывшее в большом ходу прежде, при отсутствии элементов иннервации желудочных желез, так сказать, предположение поневоле, сейчас представляется даже трудно мыслимым. Надо допустить какое-то пропитывание (совсем уже не физиологический акт!) толщи слизистой оболочки пищевыми веществами для того, чтобы они могли раздражать желудочные железы. В настоящее время это предположение тем невероятнее, что последние эксперименты очень ограничивают всасывающую способность внутренней поверхности желудка, а при этом надо еще иметь в виду, что, будь всасывание, оно все же не ведет непосредственно к прониканию веществ в пепсиновые железы. Допустить проникакие веществ через отверстие желез - так же нелегко, так как во время отделительного периода ток жидкости непрерывно направляется из полости желез полость желудка. Как подтверждение разбираемого нами предположения приводилась аналогия с насекомоядными растениями, но аналогия едва ли законная. У растений нервная система еще не выделена в особую ткань и принцип, функция ее распределена, разлита по всем клеткам; в нашем же случае, при желудочных железах, мы располагаем деятельностью чрезвычайно сложного нервного прибора. К сожалению, это невероятное предположение, сколько я могу сообразить, нельзя окончательно похоронить каким-нибудь прямым опытом, оно позабудется лишь постепенно, уступая место изучению нервно-железистых явлений, которые, наверное, будут привлекать к себе все большее и большее внимание.
Доказав нервный характер всего отделительного периода при желудочных железах, я должен весь предшествующий материал представить вам еще раз как картину работы иннервационного прибора этих желез. В огромном большинстве случаев при желудочном пищеварении дело начинается с сильного центрального (автоматического) раздражения как секреторных, так и трофических волокон желудочных желез. Спустя более или менее продолжительное время после акта еды, начинают давать себя знать рефлекторные раздражители в желудке при постепенном угасании автоматического, психического раздражителя. Если съедено мясо, то центр секреторных волокон продолжает сильно раздражаться и рефлекторно со стороны желудка экстрактивными веществами мяса, между тем как центры трофических волокон, с соответственных периферических окончаний, получают только слабые толчки. В случаях хлеба совершенно наоборот: после автоматического раздражения центры секреторных волокон слабо возбуждаются со стороны периферических окончаний, но зато продолжают энергично раздражаться центры трофических со своих окончаний. В случае примеси жира к пище в центры посылаются рефлекторно-задерживающие влияния, ограничивающие деятельность как секреторных, так и трофических волокон.
Я описал железистую работу желудка такою, какою мы ее видели в наших опытах, какою она выходила из-под наших рук. Нова ли эта картина? В подробностях - да, но не в основных ее чертах. Как это ни странно, общий ее очерк имелся в науке уже целых 50 лет тому назад. Лишний повод боящимся новизны примкнуть к нашим взглядам.
У талантливого автора «Traite analytique de la digestion» (1843), Блондло, совершенно ясно говорится как о значении акта еды, так и о специфической раздражимости слизистой оболочки желудка. Конечно, фактическая разработка темы далеко не достаточна, но не надо забывать, что это были первые опыты с искусственным свищом у собаки. Поистине непостижимо, каким образом опыты Блондло и его воззрения на желудочную деятельность в продолжение протекших пятидесяти лет не только не умножились, не развились, а скорее пришли забвение благодаря ошибочным опытам и грубым взглядам последующих авторов. Отзвук труда Блондло только изредка чувствуется в физиологических сочинениях французских авторов. Из других авторов должно указать еще на Гейденгайна, так обогатившего вообще физиологию отделения и, в частности, относительно желудка сообщившего несколько верных фактов и пустившего в обращение несколько плодотворных идей. От него вышли некоторые факты и идея о расчленении отделительного процесса по периодам и по раздражителям и мысль о важном значении исследований различных сортов еды, взятых врозь, на отделительную работу желудка. [25] Книга Блондло и статья Гейденгайна, собственно говоря, исчерпывают все существенное, что имеется в физиологии за 50 с лишком лет относительно условий и механизма отделительной работы желудка во время пищеварения. Роковым для нашего вопроса, очевидно, оказался научный промах с мнимою действительностью механического момента как возбудителя желудочных желез, а он в свою очередь был грехом недостаточной методики.
 Влито 250 куб. см воды; за 30 минут отделения не было. Опять влито 250 куб. того же раствора соляной кислоты. Сок собирается в 10-минутные промежутки.
Влито 250 куб. см воды; за 30 минут отделения не было. Опять влито 250 куб. того же раствора соляной кислоты. Сок собирается в 10-минутные промежутки.
 Никакой особенной разницы в раздражающем действии разных кислот замечено не было. Пробовались фосфорная, лимонная, молочная и уксусная кислоты.
Постоянство и резкость факта делали его замечательным и исключительным. Невольно напрашивалась мысль, что в кислоте нами открыт специфический раздражитель иннервационного прибора поджелудочной железы. Сейчас же приходило в голову, что нормальное содержимое желудка имеет кислую реакцию, и эта реакция является связующим звеном между двумя соседними отделениями пищеварительного канала. Представлялось необходимым все эти интересные и важные предположения проверить, подкрепить фактами. Прежде всего были испытаны концентрации от 0.5 до 0.05%, причем получились следующие цифры.
Влито по 250 куб. см раствора соляной кислоты
Никакой особенной разницы в раздражающем действии разных кислот замечено не было. Пробовались фосфорная, лимонная, молочная и уксусная кислоты.
Постоянство и резкость факта делали его замечательным и исключительным. Невольно напрашивалась мысль, что в кислоте нами открыт специфический раздражитель иннервационного прибора поджелудочной железы. Сейчас же приходило в голову, что нормальное содержимое желудка имеет кислую реакцию, и эта реакция является связующим звеном между двумя соседними отделениями пищеварительного канала. Представлялось необходимым все эти интересные и важные предположения проверить, подкрепить фактами. Прежде всего были испытаны концентрации от 0.5 до 0.05%, причем получились следующие цифры.
Влито по 250 куб. см раствора соляной кислоты
 Можно быть уверенным, что с 0.5 % кислоты мы не достигли крайней степени напряжения железы. С другое стороны, насколько позволяют судить некоторые, не проведенные систематически, опыты, чувствительность pancreas к кислоте совпадает приблизительно с чувствительностью нашего вкусового аппарата: еле кислая на вкус жидкость еще отчетливо действовала раздражающим образом на железу. Уже пропорциональность и чувствительность значительно подкрепляли наше предположение, что кислота не есть общий, заурядный, а именно специфический раздражитель панкреатической железы. Это заключение еще более выигрывало в силе благодаря факту, что та же кислота оставалась без малейшего эффекта относительно желудочных желез.
Но у нас имеются и более убедительные опыты. Мы (проф. Широких) много раз сравнивали действие на железе, с одной стороны, перца и горчицы, с другой - кислоты. Упервых не замечалось ни следа раздражающего действия на железу. Брались навар красного перца и смесь воды с горчичным маслом, брались такой силы, которая только что была совместима с выносливостью пищеварительного канала, т. е. только что переносилась животным без рвоты. Такие жидкости, взятые в рот, обусловливали в нем отчетливое чувство жжения, и, однако, ни намека на раздражающий эффект по отношению pancreas; между тем рядом с ними слабые растворы кислоты сейчас же, как и всегда, гнали сок. Опыты доктора Готлиба, поставленные с теми же веществами (перец и горчица) на кроликах и давшие результаты, противоположные нашим, должны быть понимаемы иначе, чем понимает их автор. Очевидно, благодаря слишком большим дозам веществ имело место разрушение слизистой оболочки пищеварительного канала, и, таким образом, подвергались раздражению сами центростремительные нервы, а не периферические окончания их, которые только и обладают специфической раздражительностью.
Мне кажется, что ряд приведенных данных вполне достаточен для утвердительного ответа на вопрос специфичности кислоты как раздражителя pancreas. Как логический вывод из изученного факта являлось дальнейшее предположение, что содержимое желудка уже по одному тому должно возбуждать поджелудочную железу, что оно обладает кислой реакцией. Проверка предположения, конечно, не представляла ничего трудного. Прежде всего легко убедились в том, что чистый желудочный сок - столь же сильный раздражитель pancreas, как и соответствующий ему раствор соляной кислоты. Далее - растворы разных сортов сахара, пептона, белков, введенные в желудок зондом, оказывали значительное действие на панкреатическое отделение только в том случае, если они обладали резко кислой реакцией; в случае же нейтральной реакции или щелочной их сокогонное действие равнялось воде или же было меньше. Ocoбенно убедительным наше рассуждение стало бы в том случае, если бы удалось у пищи, находящейся желудке и возбуждающей обыкновенным порядком панкреатическую железу, отнять это действие посредством нейтрализации. Это и есть. Если животному, находящемуся в периоде пищеварения и обильно отделяющему панкреатический сок, ввести в желудок через зонд или через обыкновенную желудочную фистулу соду, известковую воду и самый панкреатический сок, во всех этих случаях, спустя несколько минут, начинается задерживание нормального отделения, заходящее иногда очень далеко. Для примера привожу опыт.
Отделение собирается и отмечается в 5-минутные промежутки.
Можно быть уверенным, что с 0.5 % кислоты мы не достигли крайней степени напряжения железы. С другое стороны, насколько позволяют судить некоторые, не проведенные систематически, опыты, чувствительность pancreas к кислоте совпадает приблизительно с чувствительностью нашего вкусового аппарата: еле кислая на вкус жидкость еще отчетливо действовала раздражающим образом на железу. Уже пропорциональность и чувствительность значительно подкрепляли наше предположение, что кислота не есть общий, заурядный, а именно специфический раздражитель панкреатической железы. Это заключение еще более выигрывало в силе благодаря факту, что та же кислота оставалась без малейшего эффекта относительно желудочных желез.
Но у нас имеются и более убедительные опыты. Мы (проф. Широких) много раз сравнивали действие на железе, с одной стороны, перца и горчицы, с другой - кислоты. Упервых не замечалось ни следа раздражающего действия на железу. Брались навар красного перца и смесь воды с горчичным маслом, брались такой силы, которая только что была совместима с выносливостью пищеварительного канала, т. е. только что переносилась животным без рвоты. Такие жидкости, взятые в рот, обусловливали в нем отчетливое чувство жжения, и, однако, ни намека на раздражающий эффект по отношению pancreas; между тем рядом с ними слабые растворы кислоты сейчас же, как и всегда, гнали сок. Опыты доктора Готлиба, поставленные с теми же веществами (перец и горчица) на кроликах и давшие результаты, противоположные нашим, должны быть понимаемы иначе, чем понимает их автор. Очевидно, благодаря слишком большим дозам веществ имело место разрушение слизистой оболочки пищеварительного канала, и, таким образом, подвергались раздражению сами центростремительные нервы, а не периферические окончания их, которые только и обладают специфической раздражительностью.
Мне кажется, что ряд приведенных данных вполне достаточен для утвердительного ответа на вопрос специфичности кислоты как раздражителя pancreas. Как логический вывод из изученного факта являлось дальнейшее предположение, что содержимое желудка уже по одному тому должно возбуждать поджелудочную железу, что оно обладает кислой реакцией. Проверка предположения, конечно, не представляла ничего трудного. Прежде всего легко убедились в том, что чистый желудочный сок - столь же сильный раздражитель pancreas, как и соответствующий ему раствор соляной кислоты. Далее - растворы разных сортов сахара, пептона, белков, введенные в желудок зондом, оказывали значительное действие на панкреатическое отделение только в том случае, если они обладали резко кислой реакцией; в случае же нейтральной реакции или щелочной их сокогонное действие равнялось воде или же было меньше. Ocoбенно убедительным наше рассуждение стало бы в том случае, если бы удалось у пищи, находящейся желудке и возбуждающей обыкновенным порядком панкреатическую железу, отнять это действие посредством нейтрализации. Это и есть. Если животному, находящемуся в периоде пищеварения и обильно отделяющему панкреатический сок, ввести в желудок через зонд или через обыкновенную желудочную фистулу соду, известковую воду и самый панкреатический сок, во всех этих случаях, спустя несколько минут, начинается задерживание нормального отделения, заходящее иногда очень далеко. Для примера привожу опыт.
Отделение собирается и отмечается в 5-минутные промежутки.

 Перед нами поучительный, уже и раньше намечавшийся, факт преемственности и связи работы одноготдела пищеварительного канала с работой последующего отдела. Слюна, увлажняя сухое, могла фигурировать в желудке в качестве раздражителя, как вода. В самом желудке психическое отделение, начиная пищеварение, тем самым, как мы видели, обеспечивает его продолжение. Этот принцип в случае кислоты как раздражителя pancreas выступает с особенной ясностью.
Итак, мы с правом можем сказать, чтолота есть специфический раздражитель поджелудочной железы. Но где пункт его приложения? Возможны два предположения. Наш агент или действует местно, раздражая периферические окончания центростремительных нервов слизистой оболочки, или, вступая в кровь, раздражает центр секреторных нервов или непосредственно сами секреторные клетки. Анализ приведенных фактов и некоторые новые опыты дают хорошее основание признать: справедливым первое предположение. Продумаем внимательнее факты раздражения кислотой. Вопрос о действии кислоты в крови представляется простым: кислота уменьшает более или менее щелочность крови. Следовательно, если панкреатический сок гонится под влиянием извне введенной кислоты, то щелочность крови, конечно, должна быть уменьшена, и это единственное изменение, которое могло бы при этом наступить в крови. При нормальной же еде, когда находящаяся в желудке пищевая смесь обусловливает отделение панкреатического сока соляной кислотой, образовавшейся из крови, в крови имеется увеличение щелочности, что доказывается известным фактом повышения щелочности мочи во время пищеварительного периода. Таким образом мы имеем в крови два совершенно противоположных химических состояния при возбуждении панкреатической железы кислотой, извне введенной, и кислотой желудка. Стало быть, уже по одному этому теоретическому расчету абсолютно исключается возмощность объяснения возбуждающего действия кислоты на pancreas через кровь. Но мы можем привести и прямые опыты, говорящие в том же смысле. Кислые жидкости, введенные в прямую кишку, всегда оставляют панкреатическую железу в совершенном покое. Точно так же кислота не действует на панкреатическую железу и в том случае, если остается в желудке, не переходя в кишки. На последнее указал доктор Готлиб, но более полно обработал этот вопрос в последнее время доктор Попельский. Вот его опыт на собаке с постоянной поджелудочной фистулой.
Перед нами поучительный, уже и раньше намечавшийся, факт преемственности и связи работы одноготдела пищеварительного канала с работой последующего отдела. Слюна, увлажняя сухое, могла фигурировать в желудке в качестве раздражителя, как вода. В самом желудке психическое отделение, начиная пищеварение, тем самым, как мы видели, обеспечивает его продолжение. Этот принцип в случае кислоты как раздражителя pancreas выступает с особенной ясностью.
Итак, мы с правом можем сказать, чтолота есть специфический раздражитель поджелудочной железы. Но где пункт его приложения? Возможны два предположения. Наш агент или действует местно, раздражая периферические окончания центростремительных нервов слизистой оболочки, или, вступая в кровь, раздражает центр секреторных нервов или непосредственно сами секреторные клетки. Анализ приведенных фактов и некоторые новые опыты дают хорошее основание признать: справедливым первое предположение. Продумаем внимательнее факты раздражения кислотой. Вопрос о действии кислоты в крови представляется простым: кислота уменьшает более или менее щелочность крови. Следовательно, если панкреатический сок гонится под влиянием извне введенной кислоты, то щелочность крови, конечно, должна быть уменьшена, и это единственное изменение, которое могло бы при этом наступить в крови. При нормальной же еде, когда находящаяся в желудке пищевая смесь обусловливает отделение панкреатического сока соляной кислотой, образовавшейся из крови, в крови имеется увеличение щелочности, что доказывается известным фактом повышения щелочности мочи во время пищеварительного периода. Таким образом мы имеем в крови два совершенно противоположных химических состояния при возбуждении панкреатической железы кислотой, извне введенной, и кислотой желудка. Стало быть, уже по одному этому теоретическому расчету абсолютно исключается возмощность объяснения возбуждающего действия кислоты на pancreas через кровь. Но мы можем привести и прямые опыты, говорящие в том же смысле. Кислые жидкости, введенные в прямую кишку, всегда оставляют панкреатическую железу в совершенном покое. Точно так же кислота не действует на панкреатическую железу и в том случае, если остается в желудке, не переходя в кишки. На последнее указал доктор Готлиб, но более полно обработал этот вопрос в последнее время доктор Попельский. Вот его опыт на собаке с постоянной поджелудочной фистулой.
 С другой стороны, у нас (у д-ра Попельского) имелась собака, у которой желудок в привратниковой части был разрезан пополам и затем на местах разреза в обе части введены обыкновенные фистульные трубки. Вливание кислоты в главный желудок оставляло pancreas в покое, вливание в привратниковую часть обусловливало отделение поджелудочного сока, но лишь тогда, когда кислота переходила из нее в кишки.
Нужно быть любителем очень натянутых догадок, чтобы и при этих фактах продолжать еще думать о другой связи кислоты c pancreas, кроме рефлекторной. Нельзя при этом случае не заметить, что при pancreas, как уже сказано выше, мы совершенно свободны от необходимости обсуждать какое-нибудь непосредственное проникновение пищевых веществ в ее полость.
Затем нам остается еще один интересный вопрос: как понимать самый факт раздражающего действия кислоты? Кислота, как мы сказали, есть связующее звено между желудочным и двенадцатиперстным пищеварением. Это, конечно, бесспорный факт, но почему связью между этими отделами служит кислота, а не что-нибудь другое? Само собою разумеется, что в настоящее время нельзя претендовать на вполне научное решение этого вопроса; можно высказать только несколько предположений. Как известно, ферменты панкреатического сока действуют вполне хорошо при щелочной реакции, при слабокислой реакции это действие уже очень ослаблено, а при несколько более значительной кислой реакции оно становится нулевым. Можно поэтому представить себе, что панкреатический сок, возбуждаясь кислотой желудка и сам будучи щелочным, приготовляет для себя подходящую реакцию, нейтрализуя кислоту. Вместе с тем при этой нейтрализации, неблагоприятной для пепсина, панкреатический сок предохраняет себя от разрушающего действия этого фермента. Плодотворная мысль Брюкке о механизме прекращения желудочного пищеварения в кишках и создания благоприятных условий для кишечного пищеварения посредством желчи, таким образом, распространяется и на самый панкреатический сок.
Вместе с тем представляется вероятным и другой смысл быясняемой нами связи. Для какой-то цели, еще не вполне понятой, желудочный сок представляет собою весьма концентрированный раствор соляной кислоты. Этот раствор приготовляется, по учению современной физиологии, посредством разложения хлористого натрия крови, вследствие чего в крови получается избыток щелочей, который, ради сохранения постоянства состава крови, должен быть удален из организма. Соляная же кислота, исполнив свои обязанности в пищеварительном канале, подлежала бы всасыванию в кровь и повела бы в свою очередь к сильному понижению щелочности крови. Таким образом щелочность крови, в связи с пищеварительным актом, должна была бы проделывать большие размахи в противоположные стороны, щелочность же крови, как известно, есть существеннейший фактор химического процесса тела. Все это легко устраняется при теперешнем соотношении главных пищеварительных соков, когда кислый желудочный сок, именно вследствие его кислотности, пропорционально этой кислотности гонит щелочный панкреатический сок, т. е. в то время как из хлористого натрия крови кислотный элемент идет в пепсиновые железы и оттуда в полость желудка, элемент щелочи в виде соды служит панкреатической железе для образования панкреатического сока. Таким образом обе составные части хлористого натрия в кишках встречаются вновь, чтобы восстановить его. В последнее время доктор Вальтер дал этому объяснению солидную экспериментальную опору. Если кислота в самом деле, по крайней мере отчасти, гонит панкреатический сок потому, что должна нейтрализоваться его щелочью, то нужно ожидать в известных случаях самостоятельных изменений щелочности сока, независимо от его фереентной части и в связи именно с кислотным раздражителем. Это и оказалось в действительности. Количественные определения золы сока, ее титрование, как и титрование всего панкреатического сока, указали на резкую связь возбудителя отделения с неорганическими составными частями сока. Сок, вытекающий на раствор кислоты, всегда содержит крайне незначительное количество органического вещества, максимальное количество неорганического, так что в этом соке неорганического вещества бывает в 2-3 раза больше, чем органического, и представляет высшую щелочность золы, как и свою собственную. При этом быстрота отделения не имеет главного значения: кислотный сок сохраняет свои характерные качества при различных величинах часовой энергии. Факт совершенно аналогичный ранним фактам. Как ранее, во второй лекции, мы видели приспособление ферментов сока к сортам пищи: для хлеба увеличивался крахмальный фермент, для молока жирный, так здесь для чистой кислоты накопляется в соке щелочь и крайне уменьшается ненужная для нее его органическая часть. Однако сок, вытекающий на кислоту, никогда не бывает лишен совершенно ферментных свойств, что указывает только на частичное значение того факта, которым мы занимались: сок, следовательно, всегда рассчитывается на переваривание пищевых веществ, а не только для нейтрализации кислоты.
Вероятно, благодаря примененному способу исследования можно будет в общем потоке панкреатического сока отделять или угадывать струю обусловленного кислотой сока или, иначе сказать, узнавать между раздражителями в данный момент кислотного агента. Оказалось, что мясной сок, т. е. вытекающий при еде мяса, особенно в первые часы, в отношении неорганического остатка весьма приближается к кислотному. И это совпадает c тем, что при мясе в первые часы происходит чрезвычайно сильное отделение желудочного сока, кислота которого и является в качестве главного возбудителя панкреатического сока.
Таким образом сильнейшим раздражителей нервноотделительного прибора pancreas оказалось непитательное вещество кислота. Однако это не исключает возможности действия других раздражителей, или тождественных с раздражителями желудочных желез, или особых от них, так как ферментное действие панкреатического сока гораздо шире действия желудочного сока. Поэтому естественно представлялся вопрос: не являются ли раздражителями pancreas крахмал и жир как вещества, стоящие в специальном отношении к панкреатическому соку? По сделанным до сих пор опытам мы не могли убедиться в сокогонном действии крахмала. Разные смеси вареного крахмала с водой не гнали сока больше, чем одна вода. Предмет требует дальнейшей разработки, и весьма возможно, что какое-нибудь тонкое условие пока ускользает из наших глаз. Но, может быть, и здесь, как при желудочном соке, от крахмала исходит только трофическое действие, т. е. усиливается содержание фермента, как мы уже видели это во второй лекции, но не увеличивается масса сока. Не исключается, наконец, возможность, что пищеварительный интерес крахмала, если можно так выразиться, связан с какимнибудь другим условием, например, хоть с постоянным развитием в пищеварительном канале, при совершенно нормальном пищеварении, молочной кислоты, в особенности из углеводной пищи. Может быть, в этом лежит разгадка этого физиологохимического факта, остающегося пока совершенно темным относительно его целей и смысла. Наука еще не пробовала, да и не могла до сих пор приступать к синтезу реального пищеварения, т. е. к объединению иногда расходящихся интересов как всех пищевых веществ между собою, так и всего пищеварительного канала в отношении ко всему организму. Для понимания последней фразы прошу вспомнить историю жира в отношении желудочного отделения и вероятное предположение относительно смысла факта раздражающего действия кислоты.
Положительнее и гораздо проще вышли опыты с жиром относительно панкреатической железы. Уже одно сопоставление известных фактов делает в высшей степени вероятным, что жир должен быть самостоятельным раздражителем панкреатической железы. Мир тормозит отделение желудочного сока, следовательно, в норме нельзя ждать при жире косвенного возбуждения pancreas кислотой; остается одно прямое. Доктор Долинский при вливании зондом жидкого масла в желудок собаки постоянно наблюдал более или менее значительное отделение панкреатического сока. Хотя постоянство факта, при сильном задерживающем влиянии жира на желудочный сок, служило хорошей порукой, что мы имеем дело в этих опытах с прямым раздражением жиром панкреатической железы, однако скептик мог бы или допустить накопление в желудке кислой жидкости ранее нашего опыта, или держать в голове наше прежнее заявление, что сильное психическое возбуждение может пересилить задерживающее действие жира. Но вот вам удовлетворяющий требованию опыт, поставленный доктором Дамаскиным.
Совершенно здоровая собака, имеющая две фистулы желудочную и поджелудочную - кормлена за 20 часов до опыта; над отверстием протока фиксирована металлическая воронка с калибрированным цилиндром. Желудочная фистула посредством пробки и каучуковой трубки соединена с воронкой, поставленной на известной высоте и содержащей 110 115 куб. см жира. Каучук прерывается Т-образной трубкой, на одиночном колене которой прикрепляется также каучуковая трубка. При начале опыта моровский зажим запирает каучук между Т-образной трубкой и воронкой, т. е. не допускает выливаться жиру из воронки. Каучук, надетый на одиночную часть Т-образной трубки, открыт, т. е. содержимое желудка свободно может вытекать кнаружи. Часто в начале опыта течет из желудка чистая кислая жидкость, очевидно психические желудочный сок, реже желудок заключает в себе только немного щелочной слезы. Производящий опыт уединяется с собакой в отдельную комнату, и собака, потеряв надежду получить что-нибудь съестное и соскучившись, большею частью засыпает. Экспериментатор ждет полного прекращения кислого отделения из желудка и затем возможно осторожно открывает зажим от масла и закрывает зажим кнаружи. Если ранее, при открытом желудке, панкреатический сок или совсем не отделялся, или вытекал со скоростью 0.5 1.0 куб. см в 15 минут, то теперь, спустя 3-5 минут после вливания жира, отделение его отчетливо усиливается, достигая через 15--30 минут величины 7-10 куб. см в те же 15 минут. При этом в нижнюю каучуковую трубку спускается из желудка только небольшое количество щелочной слизи. Следовательно, отделение панкреатического сока под влиянием жира происходит при безусловном отсутствии в желудке кислого желудочного сока.
Иногда опыт видоизменялся таким образом: 15-30 минут спустя после введения жира открывается зажим на нижней трубке, и содержимое желудка выливается вон; большею частью это 15--20 куб. см жира и 3-5 куб. см щелочной слизи. Слизь маленькими порциями, несколько смешанная с каплями жира, продолжает вытекать и потом. В другом раз, рано или поздно, вместе с этой слизью из желудка выбрасывается желчь (или, чаще, желчью окрашенная жидкость), содержащая жир в щелочной реакции, очевидно, забрасываемая антиперистальтически из кишек в желудок. Тем не менее во все это время из панкреатической фистулы обильно течет сок. Сообщенные наблюдения исключают также мысль, чтобы в кишках при разложении жиров быстро развивалась и начинала преобладать кислая реакция, которая бы и была раздражителем pancreas в этих опытах: содержимое кишек в продолжение часа и больше совершенно не представляет и следов кислой реакции. Эти опыты дают право заключить, что жир есть второй и самостоятельный раздражитель панкреатической железы.
Прибавлю несколько второстепенных замечаний. При этих опытах, как сказано, час-два течет из желудка эмульсированная жидкость. Факт производит такое впечатление, что двенадцатиперстная кишка представляет такую же отдельную полость, как и желудок, постоянно перегоняя из одного конца в другой свое содержимое - в данном случае жир, смешанный с действующими на него реактивами - желчью и панкреатическим соком и подвергающийся в этой полости сечением времени, и довольно длинного, соответственному изменения, т. е. эмульсированию и разложению; через 1--2 часа эмульсированная жидкость действительно становится кислой. При пустом желудке полость двенадцатиперстной кишки как бы расширяется на счет его. Второе замечание относится ко сну животного во время опыта. Как я уже упоминал выше, сон не оказывает ни малейшего влияния на ход отделительной работы желудочных желез. Однако относительно панкреатической железы имелось категорическое заявление из нашей лаборатории, что сон чуть не прекращает совершенно панкреатического отделения, даже в самый его разгар. Дальнейшие наблюдения поправили эту ошибку. Механизм ошибки не лишен интереса. Автор, утверждавший это, повидимому, был совершенно прав: с засыпанием собаки отделение сока круто падало почти до нуля. Почему? Кроме связи. принятой автором, могла быть случайная, внешняя связь между актом засыпания и прекращением вытекания сока из протока, например хотя такая. Животное стоит в станке в особых так называемых сапогах. Когда оно засыпает, оно, естественно, принимает пассивное положение - виснет на сапогах; при этом происходит сильное смещение и натяжение наружных покровов. В таком случае проток поджелудочной железы, идущий сквозь эти покровы, часто оказываются сжатым, придавленным, и вот вам точная, хотя и внешняя, связь сна с панкреатическим отделением. Этот факт свидетельствует еще раз о необходимости ежеминутного и широкого внимания при производстве физиологических опытов.
Что касается до пункта приложения возбуждающего действия жира на pancreas, то на основании приведенных опытов можно думать, что раздражение прикладывается к слизистой оболочке двенадцатиперстной кишки; у нас часто наблюдалось сильное и продолжительное отделение поджелудочного сока при уже совершенно свободном от жира желудке. Механизм действия жира как раздражителя поджелудочной железы едва ли может представлять собою вопрос. При химической индифферентности жира нельзя и думать о раздражении через кровь. Другое дело, конечно, периферические окончания, которые специально устроены для того, чтобы реагировать на всевозможные условия - химические, механические и всякие другие.
Теперь обратимся к раздражителяорые оказались действительными при желудочном отделении: не будут ли они действовать и при панкреатическом? Это - психическое возбуждение, вода и экстрактивные вещества. Теоретически представляются вероятными как положительный, так и отрицательный ответы. Если аппетит и вода нужны, чтобы обеспечить начало отделительной работы желудка, то ведь то же, может быть, иногда нужно и для поджелудочной железы, хотя панкреатическое отделение в главном (кислота) зависит от желудочного. При заболевании желудка поджелудочная железа осталась бы без ее главного импульса, и мы действительно знаем клинические случаи, где соляной кислоты в желудке не бывает месяцами, а пищеварение в целом идет довольно удовлетворительно. Стало быть, в интересах большей самостоятельности поджелудочной железы представляется вероятной действительность показанных выше условий и в качестве ее раздражителей. Значит, дело в фактах. Мы с умыслом отодвинули этот вопрос к концу лекции, так как решение его тесно связано с вышеустановленными фактами. Опыты над условиями, действующими в качестве специальных раздражителей pancreas, просты, как мы видели; в совершенно другом положении опыты над агентами, являющимися вместе с тем возбудителями желудочного отделения. Они будут, конечно, возбудителями и pancreas в силу кислотности желудочного сока, но это не есть решение вопроса. Необходимо разъяснить, не действуют ли они самостоятельно, прямо, независимо от кислотности желудочного сока, а это сделать не легко.
Доктор Кувшинский давно уже показал, что поддразнивание голодного животного видом пищи может обусловить иногда чрезвычайно обильное отделение панкреатического сока. Но вывод его из этого факта о психическом раздражении нервов pancreas, в свое время совершенно законный, в настоящее время, очевидно, нуждается в поверке: не имеем ли мы здесь дела с раздражающим действием кислоты желудочного сока, накопившегося в желудке под влиянием психического раздражителя? Являлось неизбежным повторение опыта в такой обстановке, чтобы косвенное влияние кислоты желудочного сока было исключено. Для этого вначале рассчитывали было на сложное оперирование животного: у собаки производились операции желудочной, панкреатической фистулы эзофаготомии. Ставя опыт с мнимым кормлением при открытой желудочной фистуле, наблюдали наступление или усиление панкреатического отделения под влиянием кормления. При таком исходе опыта смысл его оставался неопределенным; он был бы ясным только в том случае, если бы при нашей обстановке сок не потек. Теперь же опять можно было предполагать разно. Может быть, часть желудочного сока и при открытой желудочной вистуле все же успевала переходить в двенадцатиперстную кишку. Но тут же оказался и новый путь для решения нашего вопроса - это наблюдение скрытого периода мнимого кормления для pancreas. Скрытый период желудочного отделения у собаки представляет, как уже заявлено мною выше, строгую границу в одну сторону, он никогда не бывает короче 4.5 минуты. Поджелудочный сок обыкновенно начинает выделяться через 2-3 минуты при введении раздражителя, например кислоты. При поддразнивании сок начинает усиливаться также всего чаще через 2 -3 минуты после начала кормления. Это, как мне кажется, в настоящее время всего больше располагает к допущению мысли о непосредственном психическом раздражении секреторных нервов реncreas, как оно уже давно установлено для центра желудочных секреторных нервов.
Вероятно, в связи с этим фактом стоит и часто делаемое наблюдение, когда долго следишь за панкреатическим отделением голодного животного: вслед за урчанием в кишках наступает более или менее кратковременная работа железы, при покое желудочных желез. Можно думать, что быстролетное желание еды легко и одновременно захватывает центры как двигательных нервов кишек, так и секреторных нервов pancreas, будучи недостаточным для возбуждения более косного (более длинный скрытый период) желудочного отделения. Возможно, кроме того, что центр нервов pancreas как принадлежащий к кишечному отделу пищеварения более или менее тесно ассоциирован с центрами двигательных нервов кишек. Психическое же возбуждение кишечного движения - факт общеизвестный, уже вошло в пословицу о перебирании кишек при сильном аппетите или голоде. Во всяком случае над вопросом о психическом возбуждении pancreas надо еще поработать.
Подобные же вопросы, как с психическим возбуждением, приходится ставить и при воде, в ее отношении к pancreas. Влитая в желудок вода гонит панкреатический сок. Но почему прямо ли, самостоятельно возбуждая эту железу или благодаря предварительному подкислению желудочным соком? При опытах для решения этого вопроса (д-р Дамаскин) применялась та же самая методика, что описана при жире. Вливая вобаке незаметно для нее, при совершенно затихшем желудочном отделении, 150 куб. см воды, мы замечаем через 2-3 минуты начало или отчетливое усиление панкреатического отделения; ждем еще 12 минуты, когда отделение все усиливается, и теперь опоражниваем желудок. Обыкновенно в желудке еще остается некоторое количество воды, нейтральной или щелочной жидкости. Случается, что отделение pancreas продолжается некоторое время и после опорожнения желудка, хотя желудке так дело и не доходит до появления сока. В положительном случае отделение желудочного сока не начинается ранее десяти минут. Вывод ясен и бесспорен: вода есть самостоятельный, прямой раздражитель иннервационного прибора pancreas. Наконец последний вопрос относительно действия химических раздражителей желудочных желез, которые найдены нами между экстрактивными веществами мяса. Соответствующий опыт был поставлен совершенно так же, как опыт с чистой водой. Ни малейшей разницы при этом от воды не наблюдалось; отделение начиналось в тот же срок после вливания раствора либиховского экстракта, как и при воде, и отнюдь не было более энергично, чем при последнем.
Подводя итоги всем приведенным фактам, мы можем сказать, что есть несколько раздражителей, общих для желудочных и поджелудочной желез; это может быть психический момент, страстное желание еды и, несомненно, вода; а затем как те, так и другая имеют своих собственных раздражителей: желудочные - экстрактивные вещества, pancreas - кислоту и жир.
Мы должны еще несколько остановиться на явлениях задерживания, которые в некоторых случаях обнаруживаются при деятельности поджелудочной железы. Как уже упомянуто, растворы нейтральных и щелочных солей щелочных металлов не только не возбуждают панкреатического отделения, а скорее задерживают его. Я опишу эти опыты подробнее.
Сравнивалось сокогонное действие чистой воды и растворов упомянутых веществ; всякий раз отделение поджелудочного сока во втором случае было резко меньше. Представляю из работы доктора Беккера несколько цифр, сюда относящихся.
Сок собирается и отмечается в получасовые промежутки.
С другой стороны, у нас (у д-ра Попельского) имелась собака, у которой желудок в привратниковой части был разрезан пополам и затем на местах разреза в обе части введены обыкновенные фистульные трубки. Вливание кислоты в главный желудок оставляло pancreas в покое, вливание в привратниковую часть обусловливало отделение поджелудочного сока, но лишь тогда, когда кислота переходила из нее в кишки.
Нужно быть любителем очень натянутых догадок, чтобы и при этих фактах продолжать еще думать о другой связи кислоты c pancreas, кроме рефлекторной. Нельзя при этом случае не заметить, что при pancreas, как уже сказано выше, мы совершенно свободны от необходимости обсуждать какое-нибудь непосредственное проникновение пищевых веществ в ее полость.
Затем нам остается еще один интересный вопрос: как понимать самый факт раздражающего действия кислоты? Кислота, как мы сказали, есть связующее звено между желудочным и двенадцатиперстным пищеварением. Это, конечно, бесспорный факт, но почему связью между этими отделами служит кислота, а не что-нибудь другое? Само собою разумеется, что в настоящее время нельзя претендовать на вполне научное решение этого вопроса; можно высказать только несколько предположений. Как известно, ферменты панкреатического сока действуют вполне хорошо при щелочной реакции, при слабокислой реакции это действие уже очень ослаблено, а при несколько более значительной кислой реакции оно становится нулевым. Можно поэтому представить себе, что панкреатический сок, возбуждаясь кислотой желудка и сам будучи щелочным, приготовляет для себя подходящую реакцию, нейтрализуя кислоту. Вместе с тем при этой нейтрализации, неблагоприятной для пепсина, панкреатический сок предохраняет себя от разрушающего действия этого фермента. Плодотворная мысль Брюкке о механизме прекращения желудочного пищеварения в кишках и создания благоприятных условий для кишечного пищеварения посредством желчи, таким образом, распространяется и на самый панкреатический сок.
Вместе с тем представляется вероятным и другой смысл быясняемой нами связи. Для какой-то цели, еще не вполне понятой, желудочный сок представляет собою весьма концентрированный раствор соляной кислоты. Этот раствор приготовляется, по учению современной физиологии, посредством разложения хлористого натрия крови, вследствие чего в крови получается избыток щелочей, который, ради сохранения постоянства состава крови, должен быть удален из организма. Соляная же кислота, исполнив свои обязанности в пищеварительном канале, подлежала бы всасыванию в кровь и повела бы в свою очередь к сильному понижению щелочности крови. Таким образом щелочность крови, в связи с пищеварительным актом, должна была бы проделывать большие размахи в противоположные стороны, щелочность же крови, как известно, есть существеннейший фактор химического процесса тела. Все это легко устраняется при теперешнем соотношении главных пищеварительных соков, когда кислый желудочный сок, именно вследствие его кислотности, пропорционально этой кислотности гонит щелочный панкреатический сок, т. е. в то время как из хлористого натрия крови кислотный элемент идет в пепсиновые железы и оттуда в полость желудка, элемент щелочи в виде соды служит панкреатической железе для образования панкреатического сока. Таким образом обе составные части хлористого натрия в кишках встречаются вновь, чтобы восстановить его. В последнее время доктор Вальтер дал этому объяснению солидную экспериментальную опору. Если кислота в самом деле, по крайней мере отчасти, гонит панкреатический сок потому, что должна нейтрализоваться его щелочью, то нужно ожидать в известных случаях самостоятельных изменений щелочности сока, независимо от его фереентной части и в связи именно с кислотным раздражителем. Это и оказалось в действительности. Количественные определения золы сока, ее титрование, как и титрование всего панкреатического сока, указали на резкую связь возбудителя отделения с неорганическими составными частями сока. Сок, вытекающий на раствор кислоты, всегда содержит крайне незначительное количество органического вещества, максимальное количество неорганического, так что в этом соке неорганического вещества бывает в 2-3 раза больше, чем органического, и представляет высшую щелочность золы, как и свою собственную. При этом быстрота отделения не имеет главного значения: кислотный сок сохраняет свои характерные качества при различных величинах часовой энергии. Факт совершенно аналогичный ранним фактам. Как ранее, во второй лекции, мы видели приспособление ферментов сока к сортам пищи: для хлеба увеличивался крахмальный фермент, для молока жирный, так здесь для чистой кислоты накопляется в соке щелочь и крайне уменьшается ненужная для нее его органическая часть. Однако сок, вытекающий на кислоту, никогда не бывает лишен совершенно ферментных свойств, что указывает только на частичное значение того факта, которым мы занимались: сок, следовательно, всегда рассчитывается на переваривание пищевых веществ, а не только для нейтрализации кислоты.
Вероятно, благодаря примененному способу исследования можно будет в общем потоке панкреатического сока отделять или угадывать струю обусловленного кислотой сока или, иначе сказать, узнавать между раздражителями в данный момент кислотного агента. Оказалось, что мясной сок, т. е. вытекающий при еде мяса, особенно в первые часы, в отношении неорганического остатка весьма приближается к кислотному. И это совпадает c тем, что при мясе в первые часы происходит чрезвычайно сильное отделение желудочного сока, кислота которого и является в качестве главного возбудителя панкреатического сока.
Таким образом сильнейшим раздражителей нервноотделительного прибора pancreas оказалось непитательное вещество кислота. Однако это не исключает возможности действия других раздражителей, или тождественных с раздражителями желудочных желез, или особых от них, так как ферментное действие панкреатического сока гораздо шире действия желудочного сока. Поэтому естественно представлялся вопрос: не являются ли раздражителями pancreas крахмал и жир как вещества, стоящие в специальном отношении к панкреатическому соку? По сделанным до сих пор опытам мы не могли убедиться в сокогонном действии крахмала. Разные смеси вареного крахмала с водой не гнали сока больше, чем одна вода. Предмет требует дальнейшей разработки, и весьма возможно, что какое-нибудь тонкое условие пока ускользает из наших глаз. Но, может быть, и здесь, как при желудочном соке, от крахмала исходит только трофическое действие, т. е. усиливается содержание фермента, как мы уже видели это во второй лекции, но не увеличивается масса сока. Не исключается, наконец, возможность, что пищеварительный интерес крахмала, если можно так выразиться, связан с какимнибудь другим условием, например, хоть с постоянным развитием в пищеварительном канале, при совершенно нормальном пищеварении, молочной кислоты, в особенности из углеводной пищи. Может быть, в этом лежит разгадка этого физиологохимического факта, остающегося пока совершенно темным относительно его целей и смысла. Наука еще не пробовала, да и не могла до сих пор приступать к синтезу реального пищеварения, т. е. к объединению иногда расходящихся интересов как всех пищевых веществ между собою, так и всего пищеварительного канала в отношении ко всему организму. Для понимания последней фразы прошу вспомнить историю жира в отношении желудочного отделения и вероятное предположение относительно смысла факта раздражающего действия кислоты.
Положительнее и гораздо проще вышли опыты с жиром относительно панкреатической железы. Уже одно сопоставление известных фактов делает в высшей степени вероятным, что жир должен быть самостоятельным раздражителем панкреатической железы. Мир тормозит отделение желудочного сока, следовательно, в норме нельзя ждать при жире косвенного возбуждения pancreas кислотой; остается одно прямое. Доктор Долинский при вливании зондом жидкого масла в желудок собаки постоянно наблюдал более или менее значительное отделение панкреатического сока. Хотя постоянство факта, при сильном задерживающем влиянии жира на желудочный сок, служило хорошей порукой, что мы имеем дело в этих опытах с прямым раздражением жиром панкреатической железы, однако скептик мог бы или допустить накопление в желудке кислой жидкости ранее нашего опыта, или держать в голове наше прежнее заявление, что сильное психическое возбуждение может пересилить задерживающее действие жира. Но вот вам удовлетворяющий требованию опыт, поставленный доктором Дамаскиным.
Совершенно здоровая собака, имеющая две фистулы желудочную и поджелудочную - кормлена за 20 часов до опыта; над отверстием протока фиксирована металлическая воронка с калибрированным цилиндром. Желудочная фистула посредством пробки и каучуковой трубки соединена с воронкой, поставленной на известной высоте и содержащей 110 115 куб. см жира. Каучук прерывается Т-образной трубкой, на одиночном колене которой прикрепляется также каучуковая трубка. При начале опыта моровский зажим запирает каучук между Т-образной трубкой и воронкой, т. е. не допускает выливаться жиру из воронки. Каучук, надетый на одиночную часть Т-образной трубки, открыт, т. е. содержимое желудка свободно может вытекать кнаружи. Часто в начале опыта течет из желудка чистая кислая жидкость, очевидно психические желудочный сок, реже желудок заключает в себе только немного щелочной слезы. Производящий опыт уединяется с собакой в отдельную комнату, и собака, потеряв надежду получить что-нибудь съестное и соскучившись, большею частью засыпает. Экспериментатор ждет полного прекращения кислого отделения из желудка и затем возможно осторожно открывает зажим от масла и закрывает зажим кнаружи. Если ранее, при открытом желудке, панкреатический сок или совсем не отделялся, или вытекал со скоростью 0.5 1.0 куб. см в 15 минут, то теперь, спустя 3-5 минут после вливания жира, отделение его отчетливо усиливается, достигая через 15--30 минут величины 7-10 куб. см в те же 15 минут. При этом в нижнюю каучуковую трубку спускается из желудка только небольшое количество щелочной слизи. Следовательно, отделение панкреатического сока под влиянием жира происходит при безусловном отсутствии в желудке кислого желудочного сока.
Иногда опыт видоизменялся таким образом: 15-30 минут спустя после введения жира открывается зажим на нижней трубке, и содержимое желудка выливается вон; большею частью это 15--20 куб. см жира и 3-5 куб. см щелочной слизи. Слизь маленькими порциями, несколько смешанная с каплями жира, продолжает вытекать и потом. В другом раз, рано или поздно, вместе с этой слизью из желудка выбрасывается желчь (или, чаще, желчью окрашенная жидкость), содержащая жир в щелочной реакции, очевидно, забрасываемая антиперистальтически из кишек в желудок. Тем не менее во все это время из панкреатической фистулы обильно течет сок. Сообщенные наблюдения исключают также мысль, чтобы в кишках при разложении жиров быстро развивалась и начинала преобладать кислая реакция, которая бы и была раздражителем pancreas в этих опытах: содержимое кишек в продолжение часа и больше совершенно не представляет и следов кислой реакции. Эти опыты дают право заключить, что жир есть второй и самостоятельный раздражитель панкреатической железы.
Прибавлю несколько второстепенных замечаний. При этих опытах, как сказано, час-два течет из желудка эмульсированная жидкость. Факт производит такое впечатление, что двенадцатиперстная кишка представляет такую же отдельную полость, как и желудок, постоянно перегоняя из одного конца в другой свое содержимое - в данном случае жир, смешанный с действующими на него реактивами - желчью и панкреатическим соком и подвергающийся в этой полости сечением времени, и довольно длинного, соответственному изменения, т. е. эмульсированию и разложению; через 1--2 часа эмульсированная жидкость действительно становится кислой. При пустом желудке полость двенадцатиперстной кишки как бы расширяется на счет его. Второе замечание относится ко сну животного во время опыта. Как я уже упоминал выше, сон не оказывает ни малейшего влияния на ход отделительной работы желудочных желез. Однако относительно панкреатической железы имелось категорическое заявление из нашей лаборатории, что сон чуть не прекращает совершенно панкреатического отделения, даже в самый его разгар. Дальнейшие наблюдения поправили эту ошибку. Механизм ошибки не лишен интереса. Автор, утверждавший это, повидимому, был совершенно прав: с засыпанием собаки отделение сока круто падало почти до нуля. Почему? Кроме связи. принятой автором, могла быть случайная, внешняя связь между актом засыпания и прекращением вытекания сока из протока, например хотя такая. Животное стоит в станке в особых так называемых сапогах. Когда оно засыпает, оно, естественно, принимает пассивное положение - виснет на сапогах; при этом происходит сильное смещение и натяжение наружных покровов. В таком случае проток поджелудочной железы, идущий сквозь эти покровы, часто оказываются сжатым, придавленным, и вот вам точная, хотя и внешняя, связь сна с панкреатическим отделением. Этот факт свидетельствует еще раз о необходимости ежеминутного и широкого внимания при производстве физиологических опытов.
Что касается до пункта приложения возбуждающего действия жира на pancreas, то на основании приведенных опытов можно думать, что раздражение прикладывается к слизистой оболочке двенадцатиперстной кишки; у нас часто наблюдалось сильное и продолжительное отделение поджелудочного сока при уже совершенно свободном от жира желудке. Механизм действия жира как раздражителя поджелудочной железы едва ли может представлять собою вопрос. При химической индифферентности жира нельзя и думать о раздражении через кровь. Другое дело, конечно, периферические окончания, которые специально устроены для того, чтобы реагировать на всевозможные условия - химические, механические и всякие другие.
Теперь обратимся к раздражителяорые оказались действительными при желудочном отделении: не будут ли они действовать и при панкреатическом? Это - психическое возбуждение, вода и экстрактивные вещества. Теоретически представляются вероятными как положительный, так и отрицательный ответы. Если аппетит и вода нужны, чтобы обеспечить начало отделительной работы желудка, то ведь то же, может быть, иногда нужно и для поджелудочной железы, хотя панкреатическое отделение в главном (кислота) зависит от желудочного. При заболевании желудка поджелудочная железа осталась бы без ее главного импульса, и мы действительно знаем клинические случаи, где соляной кислоты в желудке не бывает месяцами, а пищеварение в целом идет довольно удовлетворительно. Стало быть, в интересах большей самостоятельности поджелудочной железы представляется вероятной действительность показанных выше условий и в качестве ее раздражителей. Значит, дело в фактах. Мы с умыслом отодвинули этот вопрос к концу лекции, так как решение его тесно связано с вышеустановленными фактами. Опыты над условиями, действующими в качестве специальных раздражителей pancreas, просты, как мы видели; в совершенно другом положении опыты над агентами, являющимися вместе с тем возбудителями желудочного отделения. Они будут, конечно, возбудителями и pancreas в силу кислотности желудочного сока, но это не есть решение вопроса. Необходимо разъяснить, не действуют ли они самостоятельно, прямо, независимо от кислотности желудочного сока, а это сделать не легко.
Доктор Кувшинский давно уже показал, что поддразнивание голодного животного видом пищи может обусловить иногда чрезвычайно обильное отделение панкреатического сока. Но вывод его из этого факта о психическом раздражении нервов pancreas, в свое время совершенно законный, в настоящее время, очевидно, нуждается в поверке: не имеем ли мы здесь дела с раздражающим действием кислоты желудочного сока, накопившегося в желудке под влиянием психического раздражителя? Являлось неизбежным повторение опыта в такой обстановке, чтобы косвенное влияние кислоты желудочного сока было исключено. Для этого вначале рассчитывали было на сложное оперирование животного: у собаки производились операции желудочной, панкреатической фистулы эзофаготомии. Ставя опыт с мнимым кормлением при открытой желудочной фистуле, наблюдали наступление или усиление панкреатического отделения под влиянием кормления. При таком исходе опыта смысл его оставался неопределенным; он был бы ясным только в том случае, если бы при нашей обстановке сок не потек. Теперь же опять можно было предполагать разно. Может быть, часть желудочного сока и при открытой желудочной вистуле все же успевала переходить в двенадцатиперстную кишку. Но тут же оказался и новый путь для решения нашего вопроса - это наблюдение скрытого периода мнимого кормления для pancreas. Скрытый период желудочного отделения у собаки представляет, как уже заявлено мною выше, строгую границу в одну сторону, он никогда не бывает короче 4.5 минуты. Поджелудочный сок обыкновенно начинает выделяться через 2-3 минуты при введении раздражителя, например кислоты. При поддразнивании сок начинает усиливаться также всего чаще через 2 -3 минуты после начала кормления. Это, как мне кажется, в настоящее время всего больше располагает к допущению мысли о непосредственном психическом раздражении секреторных нервов реncreas, как оно уже давно установлено для центра желудочных секреторных нервов.
Вероятно, в связи с этим фактом стоит и часто делаемое наблюдение, когда долго следишь за панкреатическим отделением голодного животного: вслед за урчанием в кишках наступает более или менее кратковременная работа железы, при покое желудочных желез. Можно думать, что быстролетное желание еды легко и одновременно захватывает центры как двигательных нервов кишек, так и секреторных нервов pancreas, будучи недостаточным для возбуждения более косного (более длинный скрытый период) желудочного отделения. Возможно, кроме того, что центр нервов pancreas как принадлежащий к кишечному отделу пищеварения более или менее тесно ассоциирован с центрами двигательных нервов кишек. Психическое же возбуждение кишечного движения - факт общеизвестный, уже вошло в пословицу о перебирании кишек при сильном аппетите или голоде. Во всяком случае над вопросом о психическом возбуждении pancreas надо еще поработать.
Подобные же вопросы, как с психическим возбуждением, приходится ставить и при воде, в ее отношении к pancreas. Влитая в желудок вода гонит панкреатический сок. Но почему прямо ли, самостоятельно возбуждая эту железу или благодаря предварительному подкислению желудочным соком? При опытах для решения этого вопроса (д-р Дамаскин) применялась та же самая методика, что описана при жире. Вливая вобаке незаметно для нее, при совершенно затихшем желудочном отделении, 150 куб. см воды, мы замечаем через 2-3 минуты начало или отчетливое усиление панкреатического отделения; ждем еще 12 минуты, когда отделение все усиливается, и теперь опоражниваем желудок. Обыкновенно в желудке еще остается некоторое количество воды, нейтральной или щелочной жидкости. Случается, что отделение pancreas продолжается некоторое время и после опорожнения желудка, хотя желудке так дело и не доходит до появления сока. В положительном случае отделение желудочного сока не начинается ранее десяти минут. Вывод ясен и бесспорен: вода есть самостоятельный, прямой раздражитель иннервационного прибора pancreas. Наконец последний вопрос относительно действия химических раздражителей желудочных желез, которые найдены нами между экстрактивными веществами мяса. Соответствующий опыт был поставлен совершенно так же, как опыт с чистой водой. Ни малейшей разницы при этом от воды не наблюдалось; отделение начиналось в тот же срок после вливания раствора либиховского экстракта, как и при воде, и отнюдь не было более энергично, чем при последнем.
Подводя итоги всем приведенным фактам, мы можем сказать, что есть несколько раздражителей, общих для желудочных и поджелудочной желез; это может быть психический момент, страстное желание еды и, несомненно, вода; а затем как те, так и другая имеют своих собственных раздражителей: желудочные - экстрактивные вещества, pancreas - кислоту и жир.
Мы должны еще несколько остановиться на явлениях задерживания, которые в некоторых случаях обнаруживаются при деятельности поджелудочной железы. Как уже упомянуто, растворы нейтральных и щелочных солей щелочных металлов не только не возбуждают панкреатического отделения, а скорее задерживают его. Я опишу эти опыты подробнее.
Сравнивалось сокогонное действие чистой воды и растворов упомянутых веществ; всякий раз отделение поджелудочного сока во втором случае было резко меньше. Представляю из работы доктора Беккера несколько цифр, сюда относящихся.
Сок собирается и отмечается в получасовые промежутки.
 Задерживающее действие исследовалось и вругой форме опыта, причем специально имелось в виду выяснить, как долго продолжается это действие.
Собаке вливался зондом испытуемый раствор. Спустя час животному давалась обычная еда, наступающее отделение сравнивалось с нормальным отделением после еды. Сравнение обнаруживало отчетливое уменьшение отделения в первом случае. Опять привожу пример из работы доктора Беккера (табл. 21).
Отделение отмечается каждый час (в куб. сантиметрах).
Задерживающее действие исследовалось и вругой форме опыта, причем специально имелось в виду выяснить, как долго продолжается это действие.
Собаке вливался зондом испытуемый раствор. Спустя час животному давалась обычная еда, наступающее отделение сравнивалось с нормальным отделением после еды. Сравнение обнаруживало отчетливое уменьшение отделения в первом случае. Опять привожу пример из работы доктора Беккера (табл. 21).
Отделение отмечается каждый час (в куб. сантиметрах).
 Здесь прошу вас еще раз припомнить то я говорил в первой лекции о хроническом прибавление соды к пище. Такое прибавление надолго и очень сильно понижает отделительную деятельность поджелудочной железы, дело доходит до совершенно необычного minimum работы.
Итак, факт понижающего действия наших веществ на pancreas в высшей степени ярок и, конечно, достоин внимания. Как представлять себе механизм задерживания, остается еще невполне разъясненным: местное ли это только действие на периферические окончания рефлекторных нервов или сюда вмешивается и действие через кровь, в настоящее время с положительностью сказать трудно. Что местное действие не остается без участия в разбираемом явлении, явствует уже из того факта, что задерживающее действие принадлежит не только упомянутым растворам, но и растворам других, легко растворяющихся веществ, например сахара, как то показывают опыты доктора Дамаскина. Все имеет такой вид, что легко растворяющиеся в воде вещества что-то отнимают у воды из ее обычных свойств и этим лишают ее постоянного местнораздражающего действия.
Вот все факты, которые собрала наша лаборатория относительно нормальных раздражителей поджелудочной железы. Мы считаем себя вправе признавать их новыми, хотя мысль о специально раздражающем действии кислоты и кислой пищевой массы желудка высказывалась уже давно. Но одно дело мысль и другое - ясные и точные факты. Что мысль эта, как не основанная на точных фактах, не нашла себе распространения, следует из того, что в позднейших работах и учебниках постоянно говорится лишь о раздражающем действии пищи в целом.
Я кончил, господа, с фактической частью лекций и вместе с тем чрезвычайно далек от мысли, что наш предмет исчерпан по существу. Многое, очень многое еще должно быть добыто, чтобы можно было поздравить себя с настоящей победой в этой области, но приобретенное дорсго уже по одному тому, что оно служит ясной программой для ближайшего исследования. У нас сейчас гораздо больше вопросов, чем сколько их было еще недавно. И это обилие вопросов есть успех исследования, потому что оно свидетельствует об ознакомлении с обширной областью, понятой с общей точки зрения и распланированной по отдельным участкам для производства частных работ. Вопросов так много, что о них можно говорить только группами. Во второй лекции мы познакомились с крайнею сложностью, характеризующеюся, однако, постоянством и точностью, работы желудочных и поджелудочной желез. Предстоит объяснить каждый пункт этой сложной работы с точки зрения интересов составных частей пищи, условий благосостояния пищеварительного канала, как и всего организма. Говоря частно ответить на вопросы: почему в данный момент сока то, а не другое количество, с такими, а не с другими качествами, чем эти колебания количества и свойств полезны пище для переваривания, а пищеварительному каналу и всему организму для их целостности и нормальности? За этим рядом мы имеем другой ряд вопросов: как происходят все эти колебания железистой деятельности? Мы разложили пищу на ее отдельные элементы, но приведенные далеко не обнимают собою всего количества реальных элементов. Нужно, конечно, определить все. Из действия элементов должен быть объяснен каждый пункт кривой отдаления при каждом сорте более сложной пищи. Для решения задачи должны быть применены как постепенное соединение изученных элементов, синтезы шаг за шагом все более и более сложной пищи, так и подробный анализ свойств сока в каждый момент отделительной работы. При сложной еде последнее нужно для заключения из свойств сока о характере раздражителей, как, например, из щелочности панкреатического сока можно заключать о действии кислоты в роли раздражителя. Согласие результатов по обоим способам (синтетическому и аналитическому) явится лучшим мерилом правильности заключения. Конечно, это систематическое исследование элементов пищи поведет к открытию многих неожиданных соотношений: с одной стороны - между пищевыми веществами, с другой - между пищеварительными железами. Полный ответ на две приведенные группы вопросов, зачем и каким образом колеблется железистая деятельность, мы получим только тогда, когда с приемом исследования отделительной работы соединится подробное исследование содержимого пищеварительного канала во все время пищеварительного периода, на каждом пункте всего его протяжения, когда мы будем точно знать, где какая составная часть пищи находится и каким изменениям подвергается в каждый данный момент. Последняя группа вопросов касается действия элементов пищи, т. е. точной локализации, характера действия и результата комбинирования местных специальных раздражителей, как и хода центральных иннервационных процессов, обусловливаемых не только периферическими толчками со стороны пищеварительного кала, но и влияниями с других органов. Конечно, вопросы всех этих трех групп взаимно и тесно переплетаются друг с другом.
Само собой разумеется, что все указанные вопросы имеют полную силу и для тех реактивов пищеварительного канала, которые не вошли в наше изложение, как желчь и кишечный сок, ввиду полной неудовлетворительности их физиологии с развитой в этих лекциях точки зрения. Но как ни много осталось сделать, можно быть довольным тем, что получено. Добытыми данными изгнана из нашего отдела - можно надеяться, безвозвратно грубая и бесплодная идея общей раздражительности пищеварительного канала всякими механическими, химическими и термическими агентами, безотносительно к подробностям каждой частной пищеварительной задачи. При теперешнем положении дела эти агенты, при интенсивности их действия, могли бы быть лишь благоприятствующими или возмущающими обстоятельствами, а никак не главными и нормальными определителями секреторной работы пищеварительного канала. На месте грубой подделки недостаточного знания отчетливо очерчиваются контуры художественного механизма, исполненного тонкости и внутренней целесообразности, как и все в природе, поскольку мы с ней ближе знакомимся.
Существенная польза для процесса пищеварения извлекается уже из самого инстинкта еды, так как он составляет, кроме импульса к исканию и введению в организм пищи, первый и сильнейший раздражитель многих пищеварительных желез. Выделенная таким образом масса жидкостей разных реакций переводит значительную часть принятой пищи в растворимое или полужидкое состояние, чем дается возможность проявиться химическим свойствам пищевой массы. Поэтому первоначально возбужденная деятельность железатем видоизменяется и направляется сообразно свойствам введенных пищевых веществ, как они дают себя знать периферическим окончаниям нервноотделительного прибора. В интересах всех веществ устанавливается известное равновесие в количестве и силе реактивов: одно усиливается, другое ослабляется и тормозится до известной степени, т. е. получается то содействие, то борьба отдельных элементов пищи в отношении к реактивам. Начатая актом еды отделительная работа развивается и распространяется вдоль пищеварительного канала благодаря целесообразному сцеплению одной пищеварительной инстанции с другой.
В своей речи в Обществе русских врачей, о которой упомянуто в начале этих лекций, я высказал уверенность, что через какие-нибудь 10 лет мы будем так же хорошо знать химическую работу пищеварительного канала, как знаем сейчас физический прибор нашего глаза. После этих слов прошло два года, и, оглядываясь на сделанное в течение этих лет, я не вижу причины брать моих слов назад. Оживление в разбираемом отделе знаний за последний год становится заметным и на Западе; с нашими работниками соединяются за тем же делом многочисленные езролейские товарищи, и наш предмет, раз он вышел на настоящую дорогу, по сущности дела, подлежит скорома и полному изучению. Это не вопрос о сущности жизни, о механизме или химизме деятельности клеток, окончательное решение которого останется еще на долю бесчисленного ряда научных поколений как постоянно увлекающее, но никогда вполне не удовлетворяемое желание. На нашем, так сказать, ярусе жизни, в органной физиологии (в противоположность клеточной), во многих отделах ее, уже с правом, трезво можно надеяться на возможность совершенного уяснения нормальной связи всех отдельных частей прибора (в нашем случае пищеварительного канала) между собою и с объектами внешней природы, стоящиеи к ним в специальном отношении (в данном случае с пищей). На ступени органной физиологии мы как бы абстрагируемся от вопросов, что такое периферическое окончание рефлекторных нервов и каким образом оно воспринимает того или другого раздражителя, что такое нервный процесс, как, в силу каких реакций и какого молекулярного устройства возникают в секреторной клетке те или другие ферменты и приготовляетот или другой пищеварительный реактив. Мы принимаем эти свойства и эти элементарные деятельности как готовые данные и, улавливая празила, законы их деятельности в целом приборе, можем в известных пределах управлять прибором, властвовать над ним.
Здесь прошу вас еще раз припомнить то я говорил в первой лекции о хроническом прибавление соды к пище. Такое прибавление надолго и очень сильно понижает отделительную деятельность поджелудочной железы, дело доходит до совершенно необычного minimum работы.
Итак, факт понижающего действия наших веществ на pancreas в высшей степени ярок и, конечно, достоин внимания. Как представлять себе механизм задерживания, остается еще невполне разъясненным: местное ли это только действие на периферические окончания рефлекторных нервов или сюда вмешивается и действие через кровь, в настоящее время с положительностью сказать трудно. Что местное действие не остается без участия в разбираемом явлении, явствует уже из того факта, что задерживающее действие принадлежит не только упомянутым растворам, но и растворам других, легко растворяющихся веществ, например сахара, как то показывают опыты доктора Дамаскина. Все имеет такой вид, что легко растворяющиеся в воде вещества что-то отнимают у воды из ее обычных свойств и этим лишают ее постоянного местнораздражающего действия.
Вот все факты, которые собрала наша лаборатория относительно нормальных раздражителей поджелудочной железы. Мы считаем себя вправе признавать их новыми, хотя мысль о специально раздражающем действии кислоты и кислой пищевой массы желудка высказывалась уже давно. Но одно дело мысль и другое - ясные и точные факты. Что мысль эта, как не основанная на точных фактах, не нашла себе распространения, следует из того, что в позднейших работах и учебниках постоянно говорится лишь о раздражающем действии пищи в целом.
Я кончил, господа, с фактической частью лекций и вместе с тем чрезвычайно далек от мысли, что наш предмет исчерпан по существу. Многое, очень многое еще должно быть добыто, чтобы можно было поздравить себя с настоящей победой в этой области, но приобретенное дорсго уже по одному тому, что оно служит ясной программой для ближайшего исследования. У нас сейчас гораздо больше вопросов, чем сколько их было еще недавно. И это обилие вопросов есть успех исследования, потому что оно свидетельствует об ознакомлении с обширной областью, понятой с общей точки зрения и распланированной по отдельным участкам для производства частных работ. Вопросов так много, что о них можно говорить только группами. Во второй лекции мы познакомились с крайнею сложностью, характеризующеюся, однако, постоянством и точностью, работы желудочных и поджелудочной желез. Предстоит объяснить каждый пункт этой сложной работы с точки зрения интересов составных частей пищи, условий благосостояния пищеварительного канала, как и всего организма. Говоря частно ответить на вопросы: почему в данный момент сока то, а не другое количество, с такими, а не с другими качествами, чем эти колебания количества и свойств полезны пище для переваривания, а пищеварительному каналу и всему организму для их целостности и нормальности? За этим рядом мы имеем другой ряд вопросов: как происходят все эти колебания железистой деятельности? Мы разложили пищу на ее отдельные элементы, но приведенные далеко не обнимают собою всего количества реальных элементов. Нужно, конечно, определить все. Из действия элементов должен быть объяснен каждый пункт кривой отдаления при каждом сорте более сложной пищи. Для решения задачи должны быть применены как постепенное соединение изученных элементов, синтезы шаг за шагом все более и более сложной пищи, так и подробный анализ свойств сока в каждый момент отделительной работы. При сложной еде последнее нужно для заключения из свойств сока о характере раздражителей, как, например, из щелочности панкреатического сока можно заключать о действии кислоты в роли раздражителя. Согласие результатов по обоим способам (синтетическому и аналитическому) явится лучшим мерилом правильности заключения. Конечно, это систематическое исследование элементов пищи поведет к открытию многих неожиданных соотношений: с одной стороны - между пищевыми веществами, с другой - между пищеварительными железами. Полный ответ на две приведенные группы вопросов, зачем и каким образом колеблется железистая деятельность, мы получим только тогда, когда с приемом исследования отделительной работы соединится подробное исследование содержимого пищеварительного канала во все время пищеварительного периода, на каждом пункте всего его протяжения, когда мы будем точно знать, где какая составная часть пищи находится и каким изменениям подвергается в каждый данный момент. Последняя группа вопросов касается действия элементов пищи, т. е. точной локализации, характера действия и результата комбинирования местных специальных раздражителей, как и хода центральных иннервационных процессов, обусловливаемых не только периферическими толчками со стороны пищеварительного кала, но и влияниями с других органов. Конечно, вопросы всех этих трех групп взаимно и тесно переплетаются друг с другом.
Само собой разумеется, что все указанные вопросы имеют полную силу и для тех реактивов пищеварительного канала, которые не вошли в наше изложение, как желчь и кишечный сок, ввиду полной неудовлетворительности их физиологии с развитой в этих лекциях точки зрения. Но как ни много осталось сделать, можно быть довольным тем, что получено. Добытыми данными изгнана из нашего отдела - можно надеяться, безвозвратно грубая и бесплодная идея общей раздражительности пищеварительного канала всякими механическими, химическими и термическими агентами, безотносительно к подробностям каждой частной пищеварительной задачи. При теперешнем положении дела эти агенты, при интенсивности их действия, могли бы быть лишь благоприятствующими или возмущающими обстоятельствами, а никак не главными и нормальными определителями секреторной работы пищеварительного канала. На месте грубой подделки недостаточного знания отчетливо очерчиваются контуры художественного механизма, исполненного тонкости и внутренней целесообразности, как и все в природе, поскольку мы с ней ближе знакомимся.
Существенная польза для процесса пищеварения извлекается уже из самого инстинкта еды, так как он составляет, кроме импульса к исканию и введению в организм пищи, первый и сильнейший раздражитель многих пищеварительных желез. Выделенная таким образом масса жидкостей разных реакций переводит значительную часть принятой пищи в растворимое или полужидкое состояние, чем дается возможность проявиться химическим свойствам пищевой массы. Поэтому первоначально возбужденная деятельность железатем видоизменяется и направляется сообразно свойствам введенных пищевых веществ, как они дают себя знать периферическим окончаниям нервноотделительного прибора. В интересах всех веществ устанавливается известное равновесие в количестве и силе реактивов: одно усиливается, другое ослабляется и тормозится до известной степени, т. е. получается то содействие, то борьба отдельных элементов пищи в отношении к реактивам. Начатая актом еды отделительная работа развивается и распространяется вдоль пищеварительного канала благодаря целесообразному сцеплению одной пищеварительной инстанции с другой.
В своей речи в Обществе русских врачей, о которой упомянуто в начале этих лекций, я высказал уверенность, что через какие-нибудь 10 лет мы будем так же хорошо знать химическую работу пищеварительного канала, как знаем сейчас физический прибор нашего глаза. После этих слов прошло два года, и, оглядываясь на сделанное в течение этих лет, я не вижу причины брать моих слов назад. Оживление в разбираемом отделе знаний за последний год становится заметным и на Западе; с нашими работниками соединяются за тем же делом многочисленные езролейские товарищи, и наш предмет, раз он вышел на настоящую дорогу, по сущности дела, подлежит скорома и полному изучению. Это не вопрос о сущности жизни, о механизме или химизме деятельности клеток, окончательное решение которого останется еще на долю бесчисленного ряда научных поколений как постоянно увлекающее, но никогда вполне не удовлетворяемое желание. На нашем, так сказать, ярусе жизни, в органной физиологии (в противоположность клеточной), во многих отделах ее, уже с правом, трезво можно надеяться на возможность совершенного уяснения нормальной связи всех отдельных частей прибора (в нашем случае пищеварительного канала) между собою и с объектами внешней природы, стоящиеи к ним в специальном отношении (в данном случае с пищей). На ступени органной физиологии мы как бы абстрагируемся от вопросов, что такое периферическое окончание рефлекторных нервов и каким образом оно воспринимает того или другого раздражителя, что такое нервный процесс, как, в силу каких реакций и какого молекулярного устройства возникают в секреторной клетке те или другие ферменты и приготовляетот или другой пищеварительный реактив. Мы принимаем эти свойства и эти элементарные деятельности как готовые данные и, улавливая празила, законы их деятельности в целом приборе, можем в известных пределах управлять прибором, властвовать над ним.










 О пыт 2 - й. Собака, весом 25.1 кг, в течение восьми дней до операции получает ежедневно 800 г мяса и неопределенное количество овсянки. В день операции, в 7 часов утра, собака получает еще один фунт мяса. В 9 часов утра взято 200 куб. см крови для анализа, после чего наложен жуковский свищ. Затем удалена вся моча (17 куб. см) из мочевого пузыря и экстирпированапечень, вес которой равен 551 г. Взвешенные после смерти животного остатки печени, приставшие к сосудам, весили 19 г. Непосредственно после операции собака бродит и в течение полутора часов остается, видимо, нормальной; затем у ней наступает сначала коматозное состояние, а затем клонические и тетанические судороги, в которых она и околела спустя 3 1/4 часа после опорожнения мочевого пузыря. Незадолго перед смертью взято из art. carotis 200 куб. см крови для анализа. В брюшной полости найдено 150 куб. см жидкой крови, желудок и кишки наполнены пищей, в мочевом пузыре лишь 11.5 куб. смочи слабокислой реакции. При столь небольшом количестве материала нельзя было сделать качественных проб. Для количественного анализа взято было 5 куб. см мочи для определения аммиака, 2.5 куб. см - для определения мочевины и 2.5 куб. см - для общего содержания азота. Анализы дали следующие числа.
О пыт 2 - й. Собака, весом 25.1 кг, в течение восьми дней до операции получает ежедневно 800 г мяса и неопределенное количество овсянки. В день операции, в 7 часов утра, собака получает еще один фунт мяса. В 9 часов утра взято 200 куб. см крови для анализа, после чего наложен жуковский свищ. Затем удалена вся моча (17 куб. см) из мочевого пузыря и экстирпированапечень, вес которой равен 551 г. Взвешенные после смерти животного остатки печени, приставшие к сосудам, весили 19 г. Непосредственно после операции собака бродит и в течение полутора часов остается, видимо, нормальной; затем у ней наступает сначала коматозное состояние, а затем клонические и тетанические судороги, в которых она и околела спустя 3 1/4 часа после опорожнения мочевого пузыря. Незадолго перед смертью взято из art. carotis 200 куб. см крови для анализа. В брюшной полости найдено 150 куб. см жидкой крови, желудок и кишки наполнены пищей, в мочевом пузыре лишь 11.5 куб. смочи слабокислой реакции. При столь небольшом количестве материала нельзя было сделать качественных проб. Для количественного анализа взято было 5 куб. см мочи для определения аммиака, 2.5 куб. см - для определения мочевины и 2.5 куб. см - для общего содержания азота. Анализы дали следующие числа.

 Относительно методики наших опытов мы должны сделать небольшое замечание. Известно, что собаки лишь несколько часов переживают полную экстирпацию печени. Имея это в виду, мы надеялись получить более правильную картину изменений в обмене веществ, сравнивая мочу, взятую непосредственно перед операцией, с мочой, взятой после смерти, чем при сравнении нормальной мочи, собранной в течение суток, с мочой посмертной. Мочевина в крови и в моче определялась, по Шендорфу, [30] после первого осаждения фосфорновольфрамовой и соляной кислотой, аммиак - по способу Ненцкого и Залеского [31] отгонкой в вакууме и, наконец, общее содержание азота -- по способу Кьелдаля.
Полученные нами числа обоих опытов прежде всего подтверждают ранее высказанные другими авторами (Мейстер) [32] и нами положения, именно: прибыль аммиака в крови и в моче, увеличение других азотистых составных частей мочи и уменьшение мочевины в моче. Особенно резко выступает это во 2-м опыте, где оперированная собака, хотя, правда, и в короткое время, чувствовала себя относительно хорошо, и лишь потом обнаружились судороги. В этом 2-м опыте видели также, что не только выделение мочи, но и общее содержание азота значительно уменьшилось.
Что касается содержания мочевины в крови, то в 1-м опыте кровь содержит почти одинаковое количество мочевины как до, так и после операции, а во 2-м - кровь после экстирпации печени содержит даже больше мочевины. Количество аммиака в крови после операции в обоих случаях повышенное, хотя повышение это не настолько велико, чтобы ему одному приписать причину интоксикации и смерти животного. Вероятно, существуют какието другие причины, способствующие столь быстрой смерти. Чтобы испытать, не переходят ли токсические вещества в мочу, мы испробовали токсические свойства мочи, взятой после смерти животного в 1-м опыте и оставшейся от анализа. 10 куб. см такой мочи были в 10 часов 30 минут утра подкожно впрыснуты кролику, весом 1925 г. Температура животного до инъекции равнялась 39.6°, to 4 часа после инъекции -- 39.5°, спустя 7 часов -- 40.1°. Весь день кролик был вялым и отказывался есть предлагаемую пищу. На следующий день температура упала до 39.1°, кролик совершенно оправился остался здоровым. Впрыснутая моча содержала лишь немного свертывающихся при нагревании белков. При стоянии образовался большой осадок уратов; азотная кислота, прибавленная к охлажденной моче, дала кристаллический осадок азотнокислой мочевины.
Милая, по возможности, продлить жизнь собаки после операции и, таким образом, лучше изучить последствия удаления печени из круга кровообращения, мы поставили 3-й опыт, в котором у собаки после наложения венной фистулы печень не экстирпировалась, но перевязывалась
О пыт 3 - й. Большая собака, весом 36 кг, в течение шести дней перед операцией получает ежедневно по 1.2 кг мяса. Овсянка не дается. Операция произведена в 9 часов 15 минут вечера, после того как предварительно рзято из art. cruralis 250 куб. см крови для анализа. Вслед за наложением венной фистулы и перевязкой печеночной артерии из мочевого пузыря взята вся моча. Ее количество равнялось 140 куб. см, удельный вес 1036, реакция слабощелочная. После операции животное вскоре оправилось, и только около 5 часов утра обнаружились первые клонические судороги, перешедшие постепенно в тетанические. После 6 часов животное впало в сопор и оставалось в этом состоянии до смерти, последовавшей около 8 часов 30 минут утра. Незадолго перед смертью, уже в агонии, было выпущено из art. carotis 500 куб. см крови для анализа. При вскрытии полости живота печень оказалась в первых стадиях влажной гангрены. В желудке находилось недостаточно жидкое содержание с кислой реакцией, все тонкие кишки наполнены пищевой кашицей, почки сильно гиперемиины, в мочевом пузыре 115 куб. см мочи удельного веса 1.042. Реакция мочи кислая. Качественные пробы обнаружили содержание желчного пигмента, много мочевой кислоты и белок. Количественное определение белка дало 0.39% свертывающегося при нагревании белка. Анализы крови мочи дали следующие числа.
Относительно методики наших опытов мы должны сделать небольшое замечание. Известно, что собаки лишь несколько часов переживают полную экстирпацию печени. Имея это в виду, мы надеялись получить более правильную картину изменений в обмене веществ, сравнивая мочу, взятую непосредственно перед операцией, с мочой, взятой после смерти, чем при сравнении нормальной мочи, собранной в течение суток, с мочой посмертной. Мочевина в крови и в моче определялась, по Шендорфу, [30] после первого осаждения фосфорновольфрамовой и соляной кислотой, аммиак - по способу Ненцкого и Залеского [31] отгонкой в вакууме и, наконец, общее содержание азота -- по способу Кьелдаля.
Полученные нами числа обоих опытов прежде всего подтверждают ранее высказанные другими авторами (Мейстер) [32] и нами положения, именно: прибыль аммиака в крови и в моче, увеличение других азотистых составных частей мочи и уменьшение мочевины в моче. Особенно резко выступает это во 2-м опыте, где оперированная собака, хотя, правда, и в короткое время, чувствовала себя относительно хорошо, и лишь потом обнаружились судороги. В этом 2-м опыте видели также, что не только выделение мочи, но и общее содержание азота значительно уменьшилось.
Что касается содержания мочевины в крови, то в 1-м опыте кровь содержит почти одинаковое количество мочевины как до, так и после операции, а во 2-м - кровь после экстирпации печени содержит даже больше мочевины. Количество аммиака в крови после операции в обоих случаях повышенное, хотя повышение это не настолько велико, чтобы ему одному приписать причину интоксикации и смерти животного. Вероятно, существуют какието другие причины, способствующие столь быстрой смерти. Чтобы испытать, не переходят ли токсические вещества в мочу, мы испробовали токсические свойства мочи, взятой после смерти животного в 1-м опыте и оставшейся от анализа. 10 куб. см такой мочи были в 10 часов 30 минут утра подкожно впрыснуты кролику, весом 1925 г. Температура животного до инъекции равнялась 39.6°, to 4 часа после инъекции -- 39.5°, спустя 7 часов -- 40.1°. Весь день кролик был вялым и отказывался есть предлагаемую пищу. На следующий день температура упала до 39.1°, кролик совершенно оправился остался здоровым. Впрыснутая моча содержала лишь немного свертывающихся при нагревании белков. При стоянии образовался большой осадок уратов; азотная кислота, прибавленная к охлажденной моче, дала кристаллический осадок азотнокислой мочевины.
Милая, по возможности, продлить жизнь собаки после операции и, таким образом, лучше изучить последствия удаления печени из круга кровообращения, мы поставили 3-й опыт, в котором у собаки после наложения венной фистулы печень не экстирпировалась, но перевязывалась
О пыт 3 - й. Большая собака, весом 36 кг, в течение шести дней перед операцией получает ежедневно по 1.2 кг мяса. Овсянка не дается. Операция произведена в 9 часов 15 минут вечера, после того как предварительно рзято из art. cruralis 250 куб. см крови для анализа. Вслед за наложением венной фистулы и перевязкой печеночной артерии из мочевого пузыря взята вся моча. Ее количество равнялось 140 куб. см, удельный вес 1036, реакция слабощелочная. После операции животное вскоре оправилось, и только около 5 часов утра обнаружились первые клонические судороги, перешедшие постепенно в тетанические. После 6 часов животное впало в сопор и оставалось в этом состоянии до смерти, последовавшей около 8 часов 30 минут утра. Незадолго перед смертью, уже в агонии, было выпущено из art. carotis 500 куб. см крови для анализа. При вскрытии полости живота печень оказалась в первых стадиях влажной гангрены. В желудке находилось недостаточно жидкое содержание с кислой реакцией, все тонкие кишки наполнены пищевой кашицей, почки сильно гиперемиины, в мочевом пузыре 115 куб. см мочи удельного веса 1.042. Реакция мочи кислая. Качественные пробы обнаружили содержание желчного пигмента, много мочевой кислоты и белок. Количественное определение белка дало 0.39% свертывающегося при нагревании белка. Анализы крови мочи дали следующие числа.
 К сожалению, определения аммиака в моче до и после операции в этом опыте не удались. Что касается определений мочевины, то, приняв общее содержание азота за 100, находим, что в виде мочевины выделилось 81.5% азота до операции и 47.8% после операции. А так как общее содержание азота до и после операции было почти одно и то же, то очевидно, что и в этом случае удаление печени вызвало резкое уменьшение мочевины в моче. Странно в этом опыте, что кровь до и после операции содержит одинаковые количества аммиака. Одинаковое же содержание мочевины в крови до и после операции находится в соответствии с результатами двух первых опытов. Если, таким образом, удаление печени не вызывает никаких изменений в содержании мочевины в крови и собаки, как в нашем последнем опыте, живут более 10 часов после исключения печени из круга кровообращения, выделяя при этом мочевину мочою (4.13%), то мы не можем не признать, что печень является не единственным местом образования мочевины. К совершенно подобному выводу приходит Кауфман [33] в своих исследованиях по тому же вопросу. В его опытах кровь голодавших собак содержала в среднем 32 мг мочевины в 100 г, печень - в среднем 109 мг, мозг ----- [34] 62 мг. Таким образом все названные органы содержали мочевины больше, чем кровь, и Кауфман принимает, что все они участвуют в образовании мочевины. Образуется ли мочевина в этих органах только из карбаминовокислого аммиака или посредством гидролиза из более сложных соединений, остается невыясненным. Нам лично кажется более вероятным первое предположение, так как в наших опытах у собак после мясной пищи во всех органах было найдено значительно большее содержание аммиака, чем в крови, в то время как содержание аммиака в органах при голодании сводилось к минимуму. Несомненно, что после удаления печени выделение мочи заметно уменьшается. При этом содержание мочевины в моче уменьшается не потому, что первая задерживается почками, а просто потому, что после удаления печени в теле образуется меньше мочевины. Во всех трех опытах содержание мочевины в крови до и после операции было почти одно и то же. При задержании ее после операции количество мочевины в крови должно бы было быть значительно большим. А так как на самом деле содержание мочевины остается при этом почти неизменным, то это прямо указывает на то, что, кроме печени, и другие органы участвуют в образовании мочевины и что переход мочевины из органов в кровь зависит от содержания ее в органах и регулируется определенными условиями.
В соответствии с результатами физиологических опытов, приведших нас к заключению, что печень является не единственным местом образования мочевины, стоят также клинические наблюдения при циррозе печени, острой атрофии печени и фосфорном отравлении. Теперь становятся понятными случаи, когда при тяжелых болезнях печени содержание мочевины в моче лишь ничтожно понижалось или даже оставалось без изменения. Хотя клиницисты и утверждали, [35] что трудно найти более веский аргумент против мочевинообразовательной функции печени и что вопрос о месте образования мочевины в организме млекопитающих остается открытым, тем не менее все это совершенно неверно. Мы еще раз повторяем здесь то, что было уже сказано нами в последнем сообщении: «Что печень обладает мочевинообразовательной функцией, с несомненностью явствует: 1) из трансфузионных опытов Шредера и Соломона, 2) из факта задерживания печенью приносимого воротной веной аммиака и 3) из значительного уменьшения содержания мочевины в моче после возможно полной экстирпации печени». 2 [36]
Исследованные Мюнцером и Рихтером случаи острой атрофии печени доказывают как раз противоположное тому, что утверждают названные авторы. В двух сообщаемых Мюнцером 3 [37] случаях, когда при микроскопическом исследовании не было вовсе найдено нормальной печеночной паренхимы, моча содержала в случае № 11 (общее содержание азота равнялось 100) 52.4% в виде мочевины, 36.7% в виде аммиака и 10.9% 0/ в виде других азотистых соединений. В случае № 13 азот мочи распределятся следующим образом: 52.9% мочевинного азота, 17.3% аммиачного и 29.8% азота в виде других составных частей мочи. Эти числа очень близки к тем, которые мы получали при возможно полной экстирпации печени, как, например, в нашем опыте втором. В случае № 12, где микроскопическое исследование обнаружило присутствие довольно большого числа хорошо сохранившихся печеночных долек, Мюнцер получил почти нормальные числа: 91.8 % мочевинного азота, 6.9% аммиачного и только 1.3% азота в виде других составных частей мочи. В первом случае, описанном Рихтером, где микроскопически было констатировано почти полное разрушение печеночных клеток, моча за последние два дня перед смертью имела следующий состав (общее содержание N равнялось 100): 61% мочевинного азота, 10 % аммиачного и 5.9% 5 0/ азота аллоксуровых веществ, а также: 72 мочевинного азота, 16% аммиачного и 6.6 % азота аллоксуровых веществ. Таким образом и здесь замечалось ясное увеличение содержания аммиака и уменьшение мочевины. Второй случай, приводимый Рихтером, не имеет отношения к интересующему нас вопросу, так как за последние два дня перед смертью моча не была собрана.
Из приведенных чисел видно, что увеличение аммиака и уменьшение мочевины в моче тем значительнее, чем полнее разрушение паренхимы печени. Если же хотя небольшая часть паренхимы сохранилась, то изменения в составе мочи так ничтожны, что разницу можно всегда приписать погрешностям определений.
Неизменность состава мочи в данном случае объясняется, во-первых, тем обстоятельством, что оставшиеся нормальные печеночные клетки работают с усиленной энергией, а во-вторых, тем, что с накоплением аммиака в крови, по всей вероятности, и остальные органы образуют больше мочевины. Но при этом они могут восполнять функции печени лишь до известной степени и на непродолжительное время, что мы наглядно видели на наших собаках с венной фистулой.
В нашей первой работе, произведенной совместно с Ганом и Массеном, [38] мы показали, что резкие изменения в составе мочи наступают лишь тогда, когда у животного обнаруживаются тяжелые симптомы отравления карбаминовой кислотою. Магнамини в своей недавно появившейся работа 1 [39] занялся повторением наших опытов. Он делал анализ мочи за несколько дней до наложения венной фистулы и затем 2-3 дня после операции. Уже вследствие одного этого можно было предполагать, что его определения окажутся непригодными для решения вопроса об образовании мочевины в печени. Кроме того, Магнамини налагал фистулы по способу, видоизмененному проф. Квейроло, что в данном случае явилось не улучшением, а ухудшением метода, употребляемого нами. Квейроло [40] сшивает воротную вену с нижней полой не тотчас под печенью, а несколько ниже, под впадением vena pancreaticoduodenalis, которую он перевязывает. Мы видели в наших опытах, что в этих случаях явления интоксикации могут не наступать, так как кровь vena pancreaticoduodenalis, этой чрезвычайно важной ветви системы воротной вены, попадает в печень при помощи малых сосудов, расположенных в ligamentum hepatogastroduodenale и образующих коллатеральное кровообращение. Применением модификации Квейроло при оперировании собак объясняются также застой в венной системе и альбуминурия у собак Магнамини. У наших собак с ванной фистулой все время до самой смерти моча оставалась безбелковой; точно так же причиной альбуминурии нельзя признать применение морфия. Наконец в работе Магнамини встречаются ошибки в подсчетах, так что заключения, выводимые автором на основании полученных чисел, не всегда отвечают истинному положению дела.
Так, в своем третьем опыте Магнамини следующим образом высчитывает состав мочи по азоту (принев общее количество азота равным 100).
К сожалению, определения аммиака в моче до и после операции в этом опыте не удались. Что касается определений мочевины, то, приняв общее содержание азота за 100, находим, что в виде мочевины выделилось 81.5% азота до операции и 47.8% после операции. А так как общее содержание азота до и после операции было почти одно и то же, то очевидно, что и в этом случае удаление печени вызвало резкое уменьшение мочевины в моче. Странно в этом опыте, что кровь до и после операции содержит одинаковые количества аммиака. Одинаковое же содержание мочевины в крови до и после операции находится в соответствии с результатами двух первых опытов. Если, таким образом, удаление печени не вызывает никаких изменений в содержании мочевины в крови и собаки, как в нашем последнем опыте, живут более 10 часов после исключения печени из круга кровообращения, выделяя при этом мочевину мочою (4.13%), то мы не можем не признать, что печень является не единственным местом образования мочевины. К совершенно подобному выводу приходит Кауфман [33] в своих исследованиях по тому же вопросу. В его опытах кровь голодавших собак содержала в среднем 32 мг мочевины в 100 г, печень - в среднем 109 мг, мозг ----- [34] 62 мг. Таким образом все названные органы содержали мочевины больше, чем кровь, и Кауфман принимает, что все они участвуют в образовании мочевины. Образуется ли мочевина в этих органах только из карбаминовокислого аммиака или посредством гидролиза из более сложных соединений, остается невыясненным. Нам лично кажется более вероятным первое предположение, так как в наших опытах у собак после мясной пищи во всех органах было найдено значительно большее содержание аммиака, чем в крови, в то время как содержание аммиака в органах при голодании сводилось к минимуму. Несомненно, что после удаления печени выделение мочи заметно уменьшается. При этом содержание мочевины в моче уменьшается не потому, что первая задерживается почками, а просто потому, что после удаления печени в теле образуется меньше мочевины. Во всех трех опытах содержание мочевины в крови до и после операции было почти одно и то же. При задержании ее после операции количество мочевины в крови должно бы было быть значительно большим. А так как на самом деле содержание мочевины остается при этом почти неизменным, то это прямо указывает на то, что, кроме печени, и другие органы участвуют в образовании мочевины и что переход мочевины из органов в кровь зависит от содержания ее в органах и регулируется определенными условиями.
В соответствии с результатами физиологических опытов, приведших нас к заключению, что печень является не единственным местом образования мочевины, стоят также клинические наблюдения при циррозе печени, острой атрофии печени и фосфорном отравлении. Теперь становятся понятными случаи, когда при тяжелых болезнях печени содержание мочевины в моче лишь ничтожно понижалось или даже оставалось без изменения. Хотя клиницисты и утверждали, [35] что трудно найти более веский аргумент против мочевинообразовательной функции печени и что вопрос о месте образования мочевины в организме млекопитающих остается открытым, тем не менее все это совершенно неверно. Мы еще раз повторяем здесь то, что было уже сказано нами в последнем сообщении: «Что печень обладает мочевинообразовательной функцией, с несомненностью явствует: 1) из трансфузионных опытов Шредера и Соломона, 2) из факта задерживания печенью приносимого воротной веной аммиака и 3) из значительного уменьшения содержания мочевины в моче после возможно полной экстирпации печени». 2 [36]
Исследованные Мюнцером и Рихтером случаи острой атрофии печени доказывают как раз противоположное тому, что утверждают названные авторы. В двух сообщаемых Мюнцером 3 [37] случаях, когда при микроскопическом исследовании не было вовсе найдено нормальной печеночной паренхимы, моча содержала в случае № 11 (общее содержание азота равнялось 100) 52.4% в виде мочевины, 36.7% в виде аммиака и 10.9% 0/ в виде других азотистых соединений. В случае № 13 азот мочи распределятся следующим образом: 52.9% мочевинного азота, 17.3% аммиачного и 29.8% азота в виде других составных частей мочи. Эти числа очень близки к тем, которые мы получали при возможно полной экстирпации печени, как, например, в нашем опыте втором. В случае № 12, где микроскопическое исследование обнаружило присутствие довольно большого числа хорошо сохранившихся печеночных долек, Мюнцер получил почти нормальные числа: 91.8 % мочевинного азота, 6.9% аммиачного и только 1.3% азота в виде других составных частей мочи. В первом случае, описанном Рихтером, где микроскопически было констатировано почти полное разрушение печеночных клеток, моча за последние два дня перед смертью имела следующий состав (общее содержание N равнялось 100): 61% мочевинного азота, 10 % аммиачного и 5.9% 5 0/ азота аллоксуровых веществ, а также: 72 мочевинного азота, 16% аммиачного и 6.6 % азота аллоксуровых веществ. Таким образом и здесь замечалось ясное увеличение содержания аммиака и уменьшение мочевины. Второй случай, приводимый Рихтером, не имеет отношения к интересующему нас вопросу, так как за последние два дня перед смертью моча не была собрана.
Из приведенных чисел видно, что увеличение аммиака и уменьшение мочевины в моче тем значительнее, чем полнее разрушение паренхимы печени. Если же хотя небольшая часть паренхимы сохранилась, то изменения в составе мочи так ничтожны, что разницу можно всегда приписать погрешностям определений.
Неизменность состава мочи в данном случае объясняется, во-первых, тем обстоятельством, что оставшиеся нормальные печеночные клетки работают с усиленной энергией, а во-вторых, тем, что с накоплением аммиака в крови, по всей вероятности, и остальные органы образуют больше мочевины. Но при этом они могут восполнять функции печени лишь до известной степени и на непродолжительное время, что мы наглядно видели на наших собаках с венной фистулой.
В нашей первой работе, произведенной совместно с Ганом и Массеном, [38] мы показали, что резкие изменения в составе мочи наступают лишь тогда, когда у животного обнаруживаются тяжелые симптомы отравления карбаминовой кислотою. Магнамини в своей недавно появившейся работа 1 [39] занялся повторением наших опытов. Он делал анализ мочи за несколько дней до наложения венной фистулы и затем 2-3 дня после операции. Уже вследствие одного этого можно было предполагать, что его определения окажутся непригодными для решения вопроса об образовании мочевины в печени. Кроме того, Магнамини налагал фистулы по способу, видоизмененному проф. Квейроло, что в данном случае явилось не улучшением, а ухудшением метода, употребляемого нами. Квейроло [40] сшивает воротную вену с нижней полой не тотчас под печенью, а несколько ниже, под впадением vena pancreaticoduodenalis, которую он перевязывает. Мы видели в наших опытах, что в этих случаях явления интоксикации могут не наступать, так как кровь vena pancreaticoduodenalis, этой чрезвычайно важной ветви системы воротной вены, попадает в печень при помощи малых сосудов, расположенных в ligamentum hepatogastroduodenale и образующих коллатеральное кровообращение. Применением модификации Квейроло при оперировании собак объясняются также застой в венной системе и альбуминурия у собак Магнамини. У наших собак с ванной фистулой все время до самой смерти моча оставалась безбелковой; точно так же причиной альбуминурии нельзя признать применение морфия. Наконец в работе Магнамини встречаются ошибки в подсчетах, так что заключения, выводимые автором на основании полученных чисел, не всегда отвечают истинному положению дела.
Так, в своем третьем опыте Магнамини следующим образом высчитывает состав мочи по азоту (принев общее количество азота равным 100).
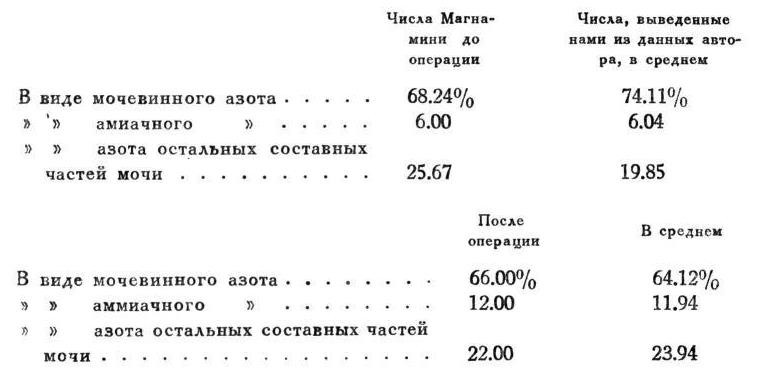 Так Такие же погрешности допущены и в четвертом, последнем спыте.
Так Такие же погрешности допущены и в четвертом, последнем спыте.
 Из своих опытов Магнамини выводит заключение, что количество мочевины после операции слегка уменьшается. Тем не менее, такого вывода из данных Магнамини сделать нельзя. В первом опыте уменьшение достигает 2.46 % (разница между 76.40 и 73.94, а не между 77.10 и 73.23, как полагает Магнамини). Опыт второй не входит в расчет, так как нет чисел состава мочи до операции. В опыте третьем уменьшение достигает 10 % (разница между 74.11 и 64.12). Напротив, в опыте четвертом мочевинный азот увеличивался на 4% после операции. В зависимости от этого нарушаются и отношения всех других азотистых составных частей мочи. Единственное заключение, которое можно вывести из данных Магнамини, это то, что собаки, оперированные по Квейроло, кроме белка, выделяют также аммиак в ненормально большом количестве.
В вышеописанных наших опытах содержание аммиака в крови после удаления печени лишь немного повышалось, так что было бы трудно считать причиной смерти животных лишь одно накопление в их организме карбаминовокислого аммиака. Однако мы не вполне согласны с замечаниями Либлейна, [41] что картины болезни, получающиеся после острого удаления печени и при наложении венной фистулы, совершенно различны. Имеющиеся особенности носят характер скорее количественной, чем качественной разницы. А известно, что тот или другой темп отравления оказывает большое влияние на картину отравления. Специально относительно аммиачных солей указываем на работу д-ра Юринского.2 2 [42] Точно так же Либлейн не прав, полагая, что симптомы интоксикации у собак с венной фистулой являются результатом скорее болезненных изменений важных и в особенности нервных аппаратов, чем последствием беспрерывно возо5новляющихся токсических инсультов. Ли блейн упускает из виду, что эти симптомы интоксикации при венной фистуле отсутствуют, как скоро перевязывается не воротная, а нижняя полая вена, 3 [43] и, кроме того, могут быть произвольно вызываемы при помощи кормления мясом или приема аммиачных солей в дозах, совершенно безвредных для неоперированных собак таких же размеров. Что сильные физические или психические раздражения могут способствовать наступлению явлений интоксикации, мы сами имели случай наблюдать, 4 [44] но, после того как было найдено, что кровь собак с венной фистулой в период интоксикации содержит более чем тройное количество аммиака по сравнению с кровью нормальной собаки, мы полагаем, что именно это накопление в крови карбаминовокислого аммиака и следует признать за причину интоксикации.
Наша собака, в крови которой было констатировано накопление карбаминовокислого аммиака, обнаруживала симптомы отравления после введения в желудок лимоннокислого аммиака. Тем не менее нам казалось интересным определить, как велико будет содержание аммиака в крови собаки, которая будет заболевать, так сказать, самопроизвольно, после обильного приема мясной пищи. С этой целью д-ром Лундбергом при нашем содействии был произведен следующий опыт.
Собака, 33.7 кг весом, 2 дня перед операцией получает ежедневно по 600 куб. см молока и по 800 г хлеба. За день перед операцией из небольшой артерии на бедре выпущено немного крови, в которой определено содержание аммиака. В 84 куб. см крови найдено 1.89 мг NH3 2.2 мг в 100 куб. см. В день операции, 8 февраля, утром собака получает только молоко. Около 12 часов произведена операция под хлороформным наркозом после того, как незадолго перед этим собаке было впрыснуто в вену 14 куб. см 1%-го раствора морфия. Венная фистула сделана возможно большей. Кровоизлияние празрезании ножницами было незначительное и притом тотчас остановлено. Операция, произведенная описанным выше способом, прошла гладко. На следующий день собака была в хорошем состоянии; до полного выздоровления ее кормили исключительно молоком и хлебом.
При этом вес животного постоянно падал, дойдя через две недели до 24 кг. Тогда было начато кормление мясом. 27 2 февраля собака в первый раз после операции получила 100 г мяса. 28 и 29 февраля собаке к обычной пище прибавлялось по 200 г мяса. 29-го наступили первые симптомы отравления, проявившиеся в атактической походке животного. ночь с 29 февраля на 1 марта симптомы отравления усилились, появилась рвота, шатание при ходьбе, подергивание конечностей, неподвижный взгляд, сильное понижение, но не исчезновение болевой чувствительности. 1 марта в 1 час 30 минут дня у собаки взято 130 куб. см крови. Во время взятия крови у животного общие судороги, которые вскоре проходят. Два определения, сделанные с этой кровью, дали следующие числа. В 55 куб. см крови найдено 3.01 мг, на 100 куб. см крови NH3 - 5.4 мг. В 69 куб. см крови найдено 4.06 мг, на 100 куб. см крови NH3 -- 5.8 мг.
В промежуток от 1 до 5 марта собака получает лишь молоко и хлеб. 5 марта собаке вновь дано 400 г мяса. Уже на следующий день можно было заметить некоторое изменение в состоянии здоровья собаки: она была не так бодра, как накануне. Это состояние постепенно усиливалось, и 9-го наступили уже ясные симптомы отравления. Содержание аммиака во взятой в этот день порции крови равнялось 3.6 мг на 100 куб. см.
10 марта собака получает белый хлеб и молоко в неограниченном количестве. За все это время моча собаки сохраняет нормальные свойства и не содержит ни белка, ни желчного пигмента. В 100 куб. см собранной 17 марта мочи было найдено 32.4 мг NH3, resp. 26.7 мг аммиачного азота. Определение общего содержания азота в 100 куб. см мочи, произведенное по Кьелдалю, дало 0.8057 г. Таким образом аммиачный азот составляет 4% 4% общего содержания азота.
19 марта в 10 часов 30 минут утра собака получает 100 г мясного порошка, что соответствует почти фунту мяса. Кроме того, было дано 80 г свежего мяса и 800 куб. см молока. В 5 5 часов пополудни животное сонливо, ходит, сильно пошатываясь, и лишь слабо реагирует на уколы иглой. В 6 часов взята проба крови, в 100 куб. см которой найдено 2.8 мг NH3. Ночью собрано у собаки 100 куб. смочи, содержащей 80.6 мг NH3, resp. 66.3 мг аммиачного азота. Определение, по Кьелдалю, общего содержания азота дало 1.77 NH3 1.458 г азота. Таким образом аммиачный азот составляет 4.5% общего содержания азота. Собака оправилась от этого припадка, и до 26 марта ее не беспокоили. В этот день она получила 1200 г мяса, большую часть которого она вырвала той же ночью. На следующий день в 10 часов утра собака вновь получает 800 г мяса, из которого 300 она выбрасывает рвотой в 3 часа пополудни. Вскоре затем наступают симптомы отравления: сильное слюноотделение, атаксия. преимущественно задних конечностей, затем слепота; по временам - подергивание личных мышц. Ночью симптомы отравления настолько усилились, что можно было опасаться за жизнь; поэтому в 3 часа утра взято для исследования 130 куб. см крови. Животное оставалось в коматозном состоянии до самой смерти, которая последовала в 7 часов 30 минут утра. За несколько минут перед смертью было взято еще 90 куб. см крови.
Тотчас произведенное вскрытие показало, что фистула была наложена удачно и отверстие было настолько велико, что о застое крови не могло быть речи. Печень мала и желта; микроскопическое исследование обнаружило ее атрофию и жировое перерождение. В почках констатировано припухание мочевых канальцев и помутнение эпителия. В мочевом пузыре найдено 520 куб. см желтой прозрачной мочи удельного веса 1.026. Моча показала щелочную реакцию, не содержала белка и сильно мутилась при стоянии на холоду с выделением уратов. Прибавление азотной кислоты к охлажденной порции мочи давалосадок азотнокислой мочевины. Определение отдельных, азотсодержащих, составные частей мочи дало следующие числа (в процентах): общее количество азота 2.253 г, аммиака 0.2078 г, resp. 0.1711 аммиачного азота, что составляет 7.6% общего количества азота. Анализ взятой ночью пробы был произведен на следующий же день утром. В 45 куб. см крови было найдено 4.24 мг NH3, следовательно в 100 куб. см крови - 9.4 мг. Такое ненормальное содержание аммиака заставило нас ближайший день повторить определение. B 36 куб. см той же крови было найдено 2.89 мг, следовательно в 100 куб. см - 8.0 мг NH3. Беря среднее из двух определений, находим, что кровь содержала 8.7% аммиака. В крови, взятой за несколько минут перед смертью, в агонии, было найдено в 44 куб. см 2.146 мг NH3, resp. в 100 куб. см 4.87 мг. Определения аммиака в органах дали следующие числа.
Из своих опытов Магнамини выводит заключение, что количество мочевины после операции слегка уменьшается. Тем не менее, такого вывода из данных Магнамини сделать нельзя. В первом опыте уменьшение достигает 2.46 % (разница между 76.40 и 73.94, а не между 77.10 и 73.23, как полагает Магнамини). Опыт второй не входит в расчет, так как нет чисел состава мочи до операции. В опыте третьем уменьшение достигает 10 % (разница между 74.11 и 64.12). Напротив, в опыте четвертом мочевинный азот увеличивался на 4% после операции. В зависимости от этого нарушаются и отношения всех других азотистых составных частей мочи. Единственное заключение, которое можно вывести из данных Магнамини, это то, что собаки, оперированные по Квейроло, кроме белка, выделяют также аммиак в ненормально большом количестве.
В вышеописанных наших опытах содержание аммиака в крови после удаления печени лишь немного повышалось, так что было бы трудно считать причиной смерти животных лишь одно накопление в их организме карбаминовокислого аммиака. Однако мы не вполне согласны с замечаниями Либлейна, [41] что картины болезни, получающиеся после острого удаления печени и при наложении венной фистулы, совершенно различны. Имеющиеся особенности носят характер скорее количественной, чем качественной разницы. А известно, что тот или другой темп отравления оказывает большое влияние на картину отравления. Специально относительно аммиачных солей указываем на работу д-ра Юринского.2 2 [42] Точно так же Либлейн не прав, полагая, что симптомы интоксикации у собак с венной фистулой являются результатом скорее болезненных изменений важных и в особенности нервных аппаратов, чем последствием беспрерывно возо5новляющихся токсических инсультов. Ли блейн упускает из виду, что эти симптомы интоксикации при венной фистуле отсутствуют, как скоро перевязывается не воротная, а нижняя полая вена, 3 [43] и, кроме того, могут быть произвольно вызываемы при помощи кормления мясом или приема аммиачных солей в дозах, совершенно безвредных для неоперированных собак таких же размеров. Что сильные физические или психические раздражения могут способствовать наступлению явлений интоксикации, мы сами имели случай наблюдать, 4 [44] но, после того как было найдено, что кровь собак с венной фистулой в период интоксикации содержит более чем тройное количество аммиака по сравнению с кровью нормальной собаки, мы полагаем, что именно это накопление в крови карбаминовокислого аммиака и следует признать за причину интоксикации.
Наша собака, в крови которой было констатировано накопление карбаминовокислого аммиака, обнаруживала симптомы отравления после введения в желудок лимоннокислого аммиака. Тем не менее нам казалось интересным определить, как велико будет содержание аммиака в крови собаки, которая будет заболевать, так сказать, самопроизвольно, после обильного приема мясной пищи. С этой целью д-ром Лундбергом при нашем содействии был произведен следующий опыт.
Собака, 33.7 кг весом, 2 дня перед операцией получает ежедневно по 600 куб. см молока и по 800 г хлеба. За день перед операцией из небольшой артерии на бедре выпущено немного крови, в которой определено содержание аммиака. В 84 куб. см крови найдено 1.89 мг NH3 2.2 мг в 100 куб. см. В день операции, 8 февраля, утром собака получает только молоко. Около 12 часов произведена операция под хлороформным наркозом после того, как незадолго перед этим собаке было впрыснуто в вену 14 куб. см 1%-го раствора морфия. Венная фистула сделана возможно большей. Кровоизлияние празрезании ножницами было незначительное и притом тотчас остановлено. Операция, произведенная описанным выше способом, прошла гладко. На следующий день собака была в хорошем состоянии; до полного выздоровления ее кормили исключительно молоком и хлебом.
При этом вес животного постоянно падал, дойдя через две недели до 24 кг. Тогда было начато кормление мясом. 27 2 февраля собака в первый раз после операции получила 100 г мяса. 28 и 29 февраля собаке к обычной пище прибавлялось по 200 г мяса. 29-го наступили первые симптомы отравления, проявившиеся в атактической походке животного. ночь с 29 февраля на 1 марта симптомы отравления усилились, появилась рвота, шатание при ходьбе, подергивание конечностей, неподвижный взгляд, сильное понижение, но не исчезновение болевой чувствительности. 1 марта в 1 час 30 минут дня у собаки взято 130 куб. см крови. Во время взятия крови у животного общие судороги, которые вскоре проходят. Два определения, сделанные с этой кровью, дали следующие числа. В 55 куб. см крови найдено 3.01 мг, на 100 куб. см крови NH3 - 5.4 мг. В 69 куб. см крови найдено 4.06 мг, на 100 куб. см крови NH3 -- 5.8 мг.
В промежуток от 1 до 5 марта собака получает лишь молоко и хлеб. 5 марта собаке вновь дано 400 г мяса. Уже на следующий день можно было заметить некоторое изменение в состоянии здоровья собаки: она была не так бодра, как накануне. Это состояние постепенно усиливалось, и 9-го наступили уже ясные симптомы отравления. Содержание аммиака во взятой в этот день порции крови равнялось 3.6 мг на 100 куб. см.
10 марта собака получает белый хлеб и молоко в неограниченном количестве. За все это время моча собаки сохраняет нормальные свойства и не содержит ни белка, ни желчного пигмента. В 100 куб. см собранной 17 марта мочи было найдено 32.4 мг NH3, resp. 26.7 мг аммиачного азота. Определение общего содержания азота в 100 куб. см мочи, произведенное по Кьелдалю, дало 0.8057 г. Таким образом аммиачный азот составляет 4% 4% общего содержания азота.
19 марта в 10 часов 30 минут утра собака получает 100 г мясного порошка, что соответствует почти фунту мяса. Кроме того, было дано 80 г свежего мяса и 800 куб. см молока. В 5 5 часов пополудни животное сонливо, ходит, сильно пошатываясь, и лишь слабо реагирует на уколы иглой. В 6 часов взята проба крови, в 100 куб. см которой найдено 2.8 мг NH3. Ночью собрано у собаки 100 куб. смочи, содержащей 80.6 мг NH3, resp. 66.3 мг аммиачного азота. Определение, по Кьелдалю, общего содержания азота дало 1.77 NH3 1.458 г азота. Таким образом аммиачный азот составляет 4.5% общего содержания азота. Собака оправилась от этого припадка, и до 26 марта ее не беспокоили. В этот день она получила 1200 г мяса, большую часть которого она вырвала той же ночью. На следующий день в 10 часов утра собака вновь получает 800 г мяса, из которого 300 она выбрасывает рвотой в 3 часа пополудни. Вскоре затем наступают симптомы отравления: сильное слюноотделение, атаксия. преимущественно задних конечностей, затем слепота; по временам - подергивание личных мышц. Ночью симптомы отравления настолько усилились, что можно было опасаться за жизнь; поэтому в 3 часа утра взято для исследования 130 куб. см крови. Животное оставалось в коматозном состоянии до самой смерти, которая последовала в 7 часов 30 минут утра. За несколько минут перед смертью было взято еще 90 куб. см крови.
Тотчас произведенное вскрытие показало, что фистула была наложена удачно и отверстие было настолько велико, что о застое крови не могло быть речи. Печень мала и желта; микроскопическое исследование обнаружило ее атрофию и жировое перерождение. В почках констатировано припухание мочевых канальцев и помутнение эпителия. В мочевом пузыре найдено 520 куб. см желтой прозрачной мочи удельного веса 1.026. Моча показала щелочную реакцию, не содержала белка и сильно мутилась при стоянии на холоду с выделением уратов. Прибавление азотной кислоты к охлажденной порции мочи давалосадок азотнокислой мочевины. Определение отдельных, азотсодержащих, составные частей мочи дало следующие числа (в процентах): общее количество азота 2.253 г, аммиака 0.2078 г, resp. 0.1711 аммиачного азота, что составляет 7.6% общего количества азота. Анализ взятой ночью пробы был произведен на следующий же день утром. В 45 куб. см крови было найдено 4.24 мг NH3, следовательно в 100 куб. см крови - 9.4 мг. Такое ненормальное содержание аммиака заставило нас ближайший день повторить определение. B 36 куб. см той же крови было найдено 2.89 мг, следовательно в 100 куб. см - 8.0 мг NH3. Беря среднее из двух определений, находим, что кровь содержала 8.7% аммиака. В крови, взятой за несколько минут перед смертью, в агонии, было найдено в 44 куб. см 2.146 мг NH3, resp. в 100 куб. см 4.87 мг. Определения аммиака в органах дали следующие числа.
 Таким образом наша собака жила 48 дней после операции, и, как видно из описания опыта, всякий раз после приема мясной пищи у собаки наступали более или менее выраженные симсимптомы отравления. Чем больше мяса получала собака, тем больше было аммиака в ее моче и крови. Особенно богата аммиаком была кровь в ночь перед смертью, когда в артериальной крови собаки было столько аммиака, сколько в наших прежних опытах мы находили лишь в v. mesenterica и pancreatica после обильного мясного питания. И в органах, особенно в мозгу и в легких, найдено довольно высокое содержание аммиака. Этим последним обстоятельством объясняется, может быть, явление, наблюдаемое у людей при прогрессивном циррозе печени, на которое впервые обращено внимание Мюнцером." [45] Мюнцер именно показал, что при этой болезни количество азота, выделяемого мочой, значительно ниже того, которое принимается вместе с пищей. В приводимом Мюнцером и исследованном Фавицким случае атрофического цирроза печени в последней стадии Фавицкий нашел (в 3-м ряду, обнимающем 7-дневный период), что в среднем из 16. 16.1 г принятого в виде пищи азота только 9.83 г выделилось мочой и 2.52 г калом. Куда исчезли 4 г, т. е. четвертая часть введенного в организм азота, анализ не показывает. Высокое содержание аммиака в крови и легких наших собак с венной фистулой показывает, что часть аммиака в газообразном виде выделяется при дыхании. Мы предполагаем продолжить наши исследования в этом направлении и своевременно сообщим о достигнутых результатах.
В заключение считаем приятной обязанностью выразить нашу благодарность И. А. Залескому, ассистенту Химического отдела Института, за его содействие при выполнении этой работы.
Таким образом наша собака жила 48 дней после операции, и, как видно из описания опыта, всякий раз после приема мясной пищи у собаки наступали более или менее выраженные симсимптомы отравления. Чем больше мяса получала собака, тем больше было аммиака в ее моче и крови. Особенно богата аммиаком была кровь в ночь перед смертью, когда в артериальной крови собаки было столько аммиака, сколько в наших прежних опытах мы находили лишь в v. mesenterica и pancreatica после обильного мясного питания. И в органах, особенно в мозгу и в легких, найдено довольно высокое содержание аммиака. Этим последним обстоятельством объясняется, может быть, явление, наблюдаемое у людей при прогрессивном циррозе печени, на которое впервые обращено внимание Мюнцером." [45] Мюнцер именно показал, что при этой болезни количество азота, выделяемого мочой, значительно ниже того, которое принимается вместе с пищей. В приводимом Мюнцером и исследованном Фавицким случае атрофического цирроза печени в последней стадии Фавицкий нашел (в 3-м ряду, обнимающем 7-дневный период), что в среднем из 16. 16.1 г принятого в виде пищи азота только 9.83 г выделилось мочой и 2.52 г калом. Куда исчезли 4 г, т. е. четвертая часть введенного в организм азота, анализ не показывает. Высокое содержание аммиака в крови и легких наших собак с венной фистулой показывает, что часть аммиака в газообразном виде выделяется при дыхании. Мы предполагаем продолжить наши исследования в этом направлении и своевременно сообщим о достигнутых результатах.
В заключение считаем приятной обязанностью выразить нашу благодарность И. А. Залескому, ассистенту Химического отдела Института, за его содействие при выполнении этой работы.
 Еще неопределеннее, еще безотраднее, чем физиология желчи как пищеварительного агента, стояла до последнего времени физиология кишечного сока. В крайнем случае он или сам отрицался как объект, или у него совершенно отрицали всякое пищеварительное действие. Относительно него абсолютно истинно, что в физиологии его нет ни одного пункта, о котором бы не спорили или не спорят. Впрочем в одном только исследователи сходятся, что пищеварительное действие кишечного сока во всяком случае второстепенное и малозначительное (слабое сахарообразующее в инвертирующее). Нашей лаборатории в самос последнее время (опыты д-ра Н. П. Шеповальникова) посчастливилось сразу и значительно поднять репутацию этой столь давно известной, но оставшейся почти без дела жидкости. Мы вышли из той же точки зрения, которая оказалась плодотворной относительно желчи: не является ли и кишечный сок также помощником, другим помощником панкреатического сока? И это тем более, что при подробном знакомстве с ролью желчи как пособника поджелудочного сока бросалось в глаза очень сильное активирование жирового фермента сравнительно с двумя другими ферментами. Можно было ожидать, что кишечный сок будет по преимуществу активировать другой какой-то фермент поджелудочного сока, как желчь жировой. Действительность вполне оправдала наши расчеты. Кишечному соку бесспорно принадлежит вполне резкая способностьусиливать действие всех панкреатических ферментов и в особенности белкового, последнего часто в поражающей степени. Тот, кто хоть раз проделает эти опыты, ни на минуту не задумается признать, что в этом и заключается главнейшее физиологическое назначение кишечного сока. Ввиду новизны и важности находки я считаю уместным продемонстрировать факт перед вами. (На экран были отброшены тени от кусков фибрина, лежащих в двух сосудах: в одном - с чистым панкреатическим соком, а в другом - со смесью этого сока с кишечным соком. В то время как в последнем сосуде в глазах аудитории последовательно один за другим сполна переварились 3 куска фибрина, в первом же только началось переваривание первого куска). Обычные приемы: кипячение сока, действие очень малыми количествами его и т. д., убедили нас, что мы имеем дело в данном случае с ферментом. Таким образом перед нами оказался не фермент той или другой части пищи, а фермент ферментов. Я предложил бы назвать его энтерокиназой, от - привожу в движение, возбуждаю, и энтеро - кишечный, потому что возможны и другие ферменты в таком роде. Нужно заметить, что в то время как активирующее действие на жировой и крахмальный ферменты панкреатического сока одинаково принадлежит кишечному соку как из duodenum, так и из других отделов тонких кишек, усиливающее действие на белковый фермент главнейшим образом характеризует дуодена:ьный сок. Первые пробные опыты дают право с большим основанием надеяться, что при совместном химическом действии трех жидкостей панкреатического сока, желчи и кишечного сока, в случае различных объектов пищеварения, откроется широчайший простор для тончайших приспособлений.
И отделение кишечного сока также, кажется, вполне своеобразно, будучи чисто местным, т. e. отделение происходит только на том участке кишки, который подвергается непосредственно раздражению. Это получает свой полный смысл ввиду того, что пищевые массы и в особенности некоторые из них только медленно передвигаются по всему кишечному каналу, и было бы бесполезно появление сока в конце кишки, когда пища только через десятки минут и даже часы окажется там.
Итак, все химические агенты пищеварения составляют род ассоциации - то цепляясь, то сменяясь, то помогая друг другу. Этот реальный синтез пищеварения я и смею считать важнейшим общим результатом наших лабораторных усилий, Вместе с тем не могу не обратить внимания на то, что прием, лежащий в основе этих работ, должен считаться своевременных и плодотворным и для других отделов физиологии. Только имея в виду целое, нормальный ход работы в том или другом отделе организма, мы без труда отличаем случайное от существенного, искусственное от нормального, легко находим новые факты и часто быстро замечаем ошибки. Идея общей, совместной работы частей проливает яркий свет на всю исследуемую область.
При анализе кривой отделения различных соков все чаще и чаще восставал перед нами вопрос о передвижении пищевых масс вдоль пищеварительного канала. Чтобы понять то или другое колебание кривых, надо было знать: где, сколько и какой находится пищи в каждый данный момент. И что же нам давала современная физиология движений пищеварительного канала? Правда, эта физиология очень обширна: каких методов ни было употреблено для изучения предмета, какие раздражения ни были испытаны, каких нервов ни раздражали, и все же эта огромная работа не давала нам ни малейшего ответа на наш вопрос. Это был чрезвычайно богатый склад, но отдельных, разрозненных аналитических данных. Почему одна пища остается тут, другая движется дальше? Почему одна движется быстро, другая медленно? Не дробится ли при передвижении сложная пища на составные части? и т. д. Все это должно происходить и происходит потому, что на пищу, сложную смесь разных веществ, в различных пунктах пищеварительного канала льются различные соки в различных комбинациях, что касается количества и качества их. Но благодаря чему, каким частным элементарным условиям передвижение пищи совершается в действительности и, очевидно, целесообразным образом с точки зрения тонкой машины? Синтез, действительный ход и механизм движения пищеварительного канала также почти не был затронут до самого последнего времени, как и синтез отделительной работы того же канала. Честь почина этого синтеза принадлежит двум немецким исследователям - Гиршу и Мерингу, одновременно открывшим важный факт, что переход содержимого желудка в кишки с количественной стороны регулируется верхним отделом кишек, рефлекторно останавливающим на время выталкивательные движения желудка и закрывающим привратник после вступления в него из желудка каждой отдельной порции. Наше исследование пошло по тому же пути и дало уже нам несколько интересных результатов. Во-первых, оказалось (опыты д-ра Сердюкова), что слизистая оболочка 12-перстной кишки определяет поступление содержимого желудка в кишки помимо массы, в связи, в отношении с кислой реакцией этого содержимого, со степенью ее. Если постоянно и понемногу вливать раствора соляной кислоты или чистого желудочного сока 12-перстную кишку через Фистулу, то налитый предварительно в желудок раствор соды можно задерживать там неопределенно долгое время. Без указанной же процедуры над кишкой щелочное содержимое желудка обыкновенно уходит из него очень скоро. Механический рефлекс тут не причем, потому что вливание соды в кишку вместо кислоты нисколько не мешает в условиях нашего опыта уходу соды из желудка. С другой стороны, было замечено, что у собак c панкреатической фистулой переход кислота из желудка в кишки резко замедлен сравнительно с собаками, не имеющими этой фистулы. Таким образом после приема каждой кислой порции из желудка кишка рефлекторным актом как бы распоряжается относительно закрытия до поры до времени выхода желудка и прекращения дальнейших выжимательных движений его. Пропущенная кислая масса вызывает на себя усиленное выделение щелочного поджелудочного сока и таким образом постепенно мало-помалу нейтрализуется. И лишь теперь опять допускается выход новой кислой порции из желудка. Благодаря такой регуляции со стороны кишки делается возможной, вместо пищеварительного хаоса, правильная смена кислого желудочного пищеварения на щелочное кишечное. В самом деле, если бы кислое содержимое желудка беспорядочно следовало в кишки, то желчь, примешивающаяся к нему, прекращала бы или очень ослабляла действие пепсина, а недостаточная притупленность кислотности массы мешала бы обнаружиться действию поджелудочных ферментов. Дело обработки пищи могло бы часто сводиться на нет. Теперь этого не происходит. Действие опасного в кишках желудочного фермента прекращается, но вместе с тем, вследствие достаточной нейтрализации массы и появления на сцене помощников поджелудочного сока - желчи и кишечного сока, поджелудочным ферментам представляется полная возможность широко развернуть свою деятельности.
Хотя ясные прямые указания на различно быстрое прохождение кислых и щелочных жидкостей из желудка в кишки встречаются уже в работе доктора Гирша, но странным образом и сам автор и другие исследователи рефлекса с кишки на поступление пищи и кишек (Меринг и Марбэ) не только не оценили факта во всем его значении, а вскоре совершенно отодвинули его в тень, исключительно увлекшись механиефлексом с тонких кишек на переход пищи из желудка.
В других наших опытах (опыты д-ра П. О. Широких) относительно самого желудка было подмечено следующее. У голодного животного, вероятно, психическим путем возбуждаются по временам опоражнивающие движения желудка. Если в это время, незаметно для животного, через фистулу желудка вливать щелочные или нейтральные жидкости (физиологический раствор поваренной соли, % раствор соды, жидкий яичный белок, молоко), то все они очень быстро, через несколько минут, в больших массах переводятся в кишки. Если же желудок с самого начала покоен, то указанные жидкости, введенные в него с тою же предосторожностью, остаются в нем многие десятки минут, совершенно не трогаясь с места. Произвольно возникшие движения желудка могут быть прекращены или чисто психически, если особенно сильно раздражать собаку едой, или, еще верней, если собаке устроить мнимое кормление (на эзофаготомированной собаке). Следовательно, раз сейчас должна войти или действительно входит в желудок пища, существующие выталкивающие движения желудка прекращаются - и до тех пор, очевидно, пока пища в нем или не будет опробована, или не подвергнется надлежащей обработке. Соответственно этому съеденное собакой молоко, в противоположность незаметно налитому в желудок, не переливается сейчас же в кишки, хотя бы перед едой и существовали выталкивающие движения желудка. Произвольные выталкивающие движения желудка прекращаются также и в том случае, если в желудок через фистулу вливаются кислые жидкости (новое доказательство вышеописанного отношения). Вы видите, господа, что и в движениях желудка при первых шагах нашего исследования мы встретились с теми же двумя особенностями, что и в наших прежних работах - целесообразностью и обнаружением разнообразных психических влияний.
Я быстро обозрел наши новые физиологические опыты. Я вижу, как еще много осталось сделать; до полного обладания предметом еще не близко, но зато дальнейшие шаги ясны, и можно иметь основательную надежду, что предмет и дальше будет так же поддаваться усилиям изучения, как и до сих пор.
Наши лабораторные животные, перенесшие нужные нам предварительные операции и служившие затем для опытов в продолжение многих месяцев и годов, по временам подвергались заболеваниям и именно того органа, над которым мы работали. Сначала это повергало нас в немалую печать. Но это было, очевидно, недоразумение, и оно, наконец, рассеялось.
Почему же патологическое состояние пищеварения не наше дело? Что такое патологическое состояние? Это встреча, соприкосновение организма с каким-нибудь чрезвычайным условием или, вернее, с необычным размером ежедневных условий. Вы подвергаетесь механическому удару, теплу или холоду, атаке со стороны патогенных микроорганизмов и т. д. в такой мере, которая превышает обыкновенную меру этих условий. Естественно начинается и особенно серьезная борьба организма с этими условиями, т. е., во-первых, пускаются в ход оборонительные приборы тела. Эти приборы - часть тела, как и всякая другая; они живут в связи с остальным телом; они участвуют в общем, жизненном равновесии тела; они, очевидно, - предмет физиологии; и я, физиолог, могу познакомиться с ними только во время болезни, иначе я не вижу их работы. Дальше. Борьба кончается или отбитием врага и прекращением работы оборонительных приборов, или победой врага -- тогда наступает полом или разрушение той или другой части организма. Разрушен орган - выпадает его функция. Но это наш обыкновенный физиологический прием, употребляемый нами для выяснения роли органа, причем часто осуществляемый природой с такой точностью, о которой мы не можем и мечтать с нашими техническими средствами. Если раздражение ограничится одним известным органом, то начинается постепенное возмещение его деятельности, наступает новое уравновешивание организма, вступают деятельность другие замещающие органы. Мы узнаем, таким образом, новые и более тонкие связи органов, скрытые их силы. Если разрушение не остановилось на одном органе, а, цепляясь, распространяется дальше, мы опять еще раз, на новый лад изучаем функциональную связь органов и, наконец, определяем тот момент и механизм, когда истощается объединяющая сила организма как целого. . Разве это с начала и до конца не физиология, углубление в связи и значение частей организма? И только какойнибудь неисправимый схоласт мог бы сказать, что это не наше дело. Напротив, именно физиолог с его компетенцией в методических и логических приемах исследования жизни является здесь самым законным работником.
Могучая сила эксперимента скоро дала себя знать и в новой для нас области - экспериментальной патологии пищеварения.
Только в лице двух работников лаборатория прямо, нарочито обратилась к патологии пищеварения, и, однако, я в состоянии сообщить уже несколько данных, способных, как мне кажется. возбудить интерес и в клиническом мире.
Наши факты пока исключительно относятся только до патологии желудочных желез. Хорошо известный членам нашего собрания способ уединенного желудочка оказался и при патологических исследованиях неоцененным способом как раскрываюший во всей ясности подробности болезненного состояния, так и чрезвычайно способствующий анализу этого болезненного состояния. Если вы болезнетворные условия прикладываете к поверхности маленького желудочка (энергическое тепло, холод, разные химически сильно действующие вещества и т. д.), то вы наблюдаете отклонение желез от нормы в идеально чистом виде, от вас не ускользает ни одна капля болезненно измененного секрета стенки, для вас не затуманивается ни одна, самая незначительная черта болезненного ее состояния. Вы наблюдаете патологическое состояние, вами вызванное, часы и дни, с начала и до конца, в тс время как во всем остальном пищеварительном канале дела идут обычным порядком и не происходит никаких осложнений со стороны последовательных расстройств остального пищеварительного канала и общего питания. Последнее понятно, так как изолированный желудочек никогда не принимает участия в пищеварительном деле, т. е. он ничего не обрабатывает и от него не идет никаких постоянных раздражений на остальной желудок и кишки, потому что он всегда пуст. Исключение представляет только небольшой период времени, когда чрезвычайный раздражитель прикладывается в качестве болезнетворной причины к его стенке и может рефлекторно влиять и на остальной пищеварительный канал. Следовательно, при таких опытах дело почти исключительно идет о знакомстве с патологическим состоянием клеток пепсиновых желез. Если же вы десиствуете болезнетворными причинами на большой желудок, то на маленьком получается возможность или видеть различные рефлекторные влияния, или наблюдать следствие измененного хода пищеварения. Таким образом представляется, между прочим, возможность отличать болезненное состояние рефлекторной поверхности, с одной стороны, и слоя желез - с другой.
Вот наши факты. Если вы вводите в маленький желудочек на минуты сильнодействующие вещества, как: абсолютный винный спирт, раствор сулемы 1 на 500, 10% раствор ляписа, сильная эмульсия горчичного масла, вы вызываете тем более или менее сильное, иногда грандиозное отделение слизи (опыты д-ра Я. X. Завриева). Перед вами, повидимому, серьезное заболевание в виде острого слизистого катара. Но болезнь ли это в самом деле? В крайнем случае слизи льется с нашей поверхности чуть ли не в 100 раз больше нормы. Иногда за весь отделительный период вы получаете только слизь вместо сока. И, однако, спрашиваю еще раз, болезнь ли это? Иногда уже через час, через два это более или менее обильное отделение слизи, наступившее непосредственно после приложения раздражающего агента, уже совершенно изгладилось. В другой раз от чрезвычайного слизетечения, заменившего в день опыта весь отделительный нормальный продукт, на другой день сверх всякого вашего ожидания не остается почти и следа. Поражает контраст между чрезвычайностью явления и его мимолетностью. Напрашивается предположение, что в описанных случаях мы совсем не имели еще дела с болезнью - перед нами разыгралась только победоносная борьба с болезнетворную причиной. В самом деле, не сейчас ли только покровный эпителий желудка развернул перед нами свое настоящее физиологическое назначение, о котором при обыкновенном течении жизни мы не имели бы случая составить себе надлежащее понятие? Своей чрезвычайной работой, изливая массу слизистой жидкости, разбавляющей, связывающей и оттесняющей от стенки введенные вредные вещества, он предотврашает опасность, которою грозят эти вещества важнейшим частям слизистой оболочки. Что наше объяснение верно, с большой достоверностью следует из факта, что при чрезвычайной деятельности покровного эпителия в данном случае эпителий пепсиновых желез остается в совершенном покое. Значит, указанные выше химические вещества раздражают только один эпителий, не трогая другого. Это то же самое, когда мясо, находясь в пищеварительном канале, раздражает только эпителий пепсиновых желез и оставляет совершенно нетронутым покровный эпителий. Мы, как мне кажется, стоим перед весьма важным фактом, что чрезвычайные раздражители, являющиеся в качестве болезнетворных причин, представляют собою специфические раздражители тех защитительных приборов организма, которые назначены для борьбы с соответствующими болезнетворными причинами. Мы думаем, что это представление должно быть обобщено на все случаи болезни, и в этом кроется общий механизм приспособлений организма вообще при встрече с патогенными условиями, совершенно подобно тому, как нормальный, сочетанный и приспособленный ход жизни имеет в своем основании специфическое раздражение того или другого аппарата.
Само собою разумеется, что действие вышеупомянутых агентов, как ни энергично исполняет свою защитительную роль покровный эпителий, может при известном размере коснуться и глубоких слоев слизистой оболочки, и тогда перед нами проходит измененная деятельность пепсиновых желез с большим разнообразием в частностях, смотря по роду агента и большею частью с физическим характером. Это есть, конечно, различные болезненные состояния пепсиновых желез, но вместе с тем и материал к физиологической характеристике этих клеток. Из имеющегося уже значительного запаса относящихся сюда интересных данных я остановлю ваше внимание на одном из них. После приложения 10 % раствора ляписа мы вызывали в пепсиновых железах крайне отчетливое состояние астении - раздражительной слабости (опыты д-ра Я. X. Завриева).
Я представляю в двух столбцах цифр часовые количества сока, изливающегося из изолированного желудочка на одно и то же количество мяса (150 г) до и во время вызванного нами специального заболевания этого желудочка.
Еще неопределеннее, еще безотраднее, чем физиология желчи как пищеварительного агента, стояла до последнего времени физиология кишечного сока. В крайнем случае он или сам отрицался как объект, или у него совершенно отрицали всякое пищеварительное действие. Относительно него абсолютно истинно, что в физиологии его нет ни одного пункта, о котором бы не спорили или не спорят. Впрочем в одном только исследователи сходятся, что пищеварительное действие кишечного сока во всяком случае второстепенное и малозначительное (слабое сахарообразующее в инвертирующее). Нашей лаборатории в самос последнее время (опыты д-ра Н. П. Шеповальникова) посчастливилось сразу и значительно поднять репутацию этой столь давно известной, но оставшейся почти без дела жидкости. Мы вышли из той же точки зрения, которая оказалась плодотворной относительно желчи: не является ли и кишечный сок также помощником, другим помощником панкреатического сока? И это тем более, что при подробном знакомстве с ролью желчи как пособника поджелудочного сока бросалось в глаза очень сильное активирование жирового фермента сравнительно с двумя другими ферментами. Можно было ожидать, что кишечный сок будет по преимуществу активировать другой какой-то фермент поджелудочного сока, как желчь жировой. Действительность вполне оправдала наши расчеты. Кишечному соку бесспорно принадлежит вполне резкая способностьусиливать действие всех панкреатических ферментов и в особенности белкового, последнего часто в поражающей степени. Тот, кто хоть раз проделает эти опыты, ни на минуту не задумается признать, что в этом и заключается главнейшее физиологическое назначение кишечного сока. Ввиду новизны и важности находки я считаю уместным продемонстрировать факт перед вами. (На экран были отброшены тени от кусков фибрина, лежащих в двух сосудах: в одном - с чистым панкреатическим соком, а в другом - со смесью этого сока с кишечным соком. В то время как в последнем сосуде в глазах аудитории последовательно один за другим сполна переварились 3 куска фибрина, в первом же только началось переваривание первого куска). Обычные приемы: кипячение сока, действие очень малыми количествами его и т. д., убедили нас, что мы имеем дело в данном случае с ферментом. Таким образом перед нами оказался не фермент той или другой части пищи, а фермент ферментов. Я предложил бы назвать его энтерокиназой, от - привожу в движение, возбуждаю, и энтеро - кишечный, потому что возможны и другие ферменты в таком роде. Нужно заметить, что в то время как активирующее действие на жировой и крахмальный ферменты панкреатического сока одинаково принадлежит кишечному соку как из duodenum, так и из других отделов тонких кишек, усиливающее действие на белковый фермент главнейшим образом характеризует дуодена:ьный сок. Первые пробные опыты дают право с большим основанием надеяться, что при совместном химическом действии трех жидкостей панкреатического сока, желчи и кишечного сока, в случае различных объектов пищеварения, откроется широчайший простор для тончайших приспособлений.
И отделение кишечного сока также, кажется, вполне своеобразно, будучи чисто местным, т. e. отделение происходит только на том участке кишки, который подвергается непосредственно раздражению. Это получает свой полный смысл ввиду того, что пищевые массы и в особенности некоторые из них только медленно передвигаются по всему кишечному каналу, и было бы бесполезно появление сока в конце кишки, когда пища только через десятки минут и даже часы окажется там.
Итак, все химические агенты пищеварения составляют род ассоциации - то цепляясь, то сменяясь, то помогая друг другу. Этот реальный синтез пищеварения я и смею считать важнейшим общим результатом наших лабораторных усилий, Вместе с тем не могу не обратить внимания на то, что прием, лежащий в основе этих работ, должен считаться своевременных и плодотворным и для других отделов физиологии. Только имея в виду целое, нормальный ход работы в том или другом отделе организма, мы без труда отличаем случайное от существенного, искусственное от нормального, легко находим новые факты и часто быстро замечаем ошибки. Идея общей, совместной работы частей проливает яркий свет на всю исследуемую область.
При анализе кривой отделения различных соков все чаще и чаще восставал перед нами вопрос о передвижении пищевых масс вдоль пищеварительного канала. Чтобы понять то или другое колебание кривых, надо было знать: где, сколько и какой находится пищи в каждый данный момент. И что же нам давала современная физиология движений пищеварительного канала? Правда, эта физиология очень обширна: каких методов ни было употреблено для изучения предмета, какие раздражения ни были испытаны, каких нервов ни раздражали, и все же эта огромная работа не давала нам ни малейшего ответа на наш вопрос. Это был чрезвычайно богатый склад, но отдельных, разрозненных аналитических данных. Почему одна пища остается тут, другая движется дальше? Почему одна движется быстро, другая медленно? Не дробится ли при передвижении сложная пища на составные части? и т. д. Все это должно происходить и происходит потому, что на пищу, сложную смесь разных веществ, в различных пунктах пищеварительного канала льются различные соки в различных комбинациях, что касается количества и качества их. Но благодаря чему, каким частным элементарным условиям передвижение пищи совершается в действительности и, очевидно, целесообразным образом с точки зрения тонкой машины? Синтез, действительный ход и механизм движения пищеварительного канала также почти не был затронут до самого последнего времени, как и синтез отделительной работы того же канала. Честь почина этого синтеза принадлежит двум немецким исследователям - Гиршу и Мерингу, одновременно открывшим важный факт, что переход содержимого желудка в кишки с количественной стороны регулируется верхним отделом кишек, рефлекторно останавливающим на время выталкивательные движения желудка и закрывающим привратник после вступления в него из желудка каждой отдельной порции. Наше исследование пошло по тому же пути и дало уже нам несколько интересных результатов. Во-первых, оказалось (опыты д-ра Сердюкова), что слизистая оболочка 12-перстной кишки определяет поступление содержимого желудка в кишки помимо массы, в связи, в отношении с кислой реакцией этого содержимого, со степенью ее. Если постоянно и понемногу вливать раствора соляной кислоты или чистого желудочного сока 12-перстную кишку через Фистулу, то налитый предварительно в желудок раствор соды можно задерживать там неопределенно долгое время. Без указанной же процедуры над кишкой щелочное содержимое желудка обыкновенно уходит из него очень скоро. Механический рефлекс тут не причем, потому что вливание соды в кишку вместо кислоты нисколько не мешает в условиях нашего опыта уходу соды из желудка. С другой стороны, было замечено, что у собак c панкреатической фистулой переход кислота из желудка в кишки резко замедлен сравнительно с собаками, не имеющими этой фистулы. Таким образом после приема каждой кислой порции из желудка кишка рефлекторным актом как бы распоряжается относительно закрытия до поры до времени выхода желудка и прекращения дальнейших выжимательных движений его. Пропущенная кислая масса вызывает на себя усиленное выделение щелочного поджелудочного сока и таким образом постепенно мало-помалу нейтрализуется. И лишь теперь опять допускается выход новой кислой порции из желудка. Благодаря такой регуляции со стороны кишки делается возможной, вместо пищеварительного хаоса, правильная смена кислого желудочного пищеварения на щелочное кишечное. В самом деле, если бы кислое содержимое желудка беспорядочно следовало в кишки, то желчь, примешивающаяся к нему, прекращала бы или очень ослабляла действие пепсина, а недостаточная притупленность кислотности массы мешала бы обнаружиться действию поджелудочных ферментов. Дело обработки пищи могло бы часто сводиться на нет. Теперь этого не происходит. Действие опасного в кишках желудочного фермента прекращается, но вместе с тем, вследствие достаточной нейтрализации массы и появления на сцене помощников поджелудочного сока - желчи и кишечного сока, поджелудочным ферментам представляется полная возможность широко развернуть свою деятельности.
Хотя ясные прямые указания на различно быстрое прохождение кислых и щелочных жидкостей из желудка в кишки встречаются уже в работе доктора Гирша, но странным образом и сам автор и другие исследователи рефлекса с кишки на поступление пищи и кишек (Меринг и Марбэ) не только не оценили факта во всем его значении, а вскоре совершенно отодвинули его в тень, исключительно увлекшись механиефлексом с тонких кишек на переход пищи из желудка.
В других наших опытах (опыты д-ра П. О. Широких) относительно самого желудка было подмечено следующее. У голодного животного, вероятно, психическим путем возбуждаются по временам опоражнивающие движения желудка. Если в это время, незаметно для животного, через фистулу желудка вливать щелочные или нейтральные жидкости (физиологический раствор поваренной соли, % раствор соды, жидкий яичный белок, молоко), то все они очень быстро, через несколько минут, в больших массах переводятся в кишки. Если же желудок с самого начала покоен, то указанные жидкости, введенные в него с тою же предосторожностью, остаются в нем многие десятки минут, совершенно не трогаясь с места. Произвольно возникшие движения желудка могут быть прекращены или чисто психически, если особенно сильно раздражать собаку едой, или, еще верней, если собаке устроить мнимое кормление (на эзофаготомированной собаке). Следовательно, раз сейчас должна войти или действительно входит в желудок пища, существующие выталкивающие движения желудка прекращаются - и до тех пор, очевидно, пока пища в нем или не будет опробована, или не подвергнется надлежащей обработке. Соответственно этому съеденное собакой молоко, в противоположность незаметно налитому в желудок, не переливается сейчас же в кишки, хотя бы перед едой и существовали выталкивающие движения желудка. Произвольные выталкивающие движения желудка прекращаются также и в том случае, если в желудок через фистулу вливаются кислые жидкости (новое доказательство вышеописанного отношения). Вы видите, господа, что и в движениях желудка при первых шагах нашего исследования мы встретились с теми же двумя особенностями, что и в наших прежних работах - целесообразностью и обнаружением разнообразных психических влияний.
Я быстро обозрел наши новые физиологические опыты. Я вижу, как еще много осталось сделать; до полного обладания предметом еще не близко, но зато дальнейшие шаги ясны, и можно иметь основательную надежду, что предмет и дальше будет так же поддаваться усилиям изучения, как и до сих пор.
Наши лабораторные животные, перенесшие нужные нам предварительные операции и служившие затем для опытов в продолжение многих месяцев и годов, по временам подвергались заболеваниям и именно того органа, над которым мы работали. Сначала это повергало нас в немалую печать. Но это было, очевидно, недоразумение, и оно, наконец, рассеялось.
Почему же патологическое состояние пищеварения не наше дело? Что такое патологическое состояние? Это встреча, соприкосновение организма с каким-нибудь чрезвычайным условием или, вернее, с необычным размером ежедневных условий. Вы подвергаетесь механическому удару, теплу или холоду, атаке со стороны патогенных микроорганизмов и т. д. в такой мере, которая превышает обыкновенную меру этих условий. Естественно начинается и особенно серьезная борьба организма с этими условиями, т. е., во-первых, пускаются в ход оборонительные приборы тела. Эти приборы - часть тела, как и всякая другая; они живут в связи с остальным телом; они участвуют в общем, жизненном равновесии тела; они, очевидно, - предмет физиологии; и я, физиолог, могу познакомиться с ними только во время болезни, иначе я не вижу их работы. Дальше. Борьба кончается или отбитием врага и прекращением работы оборонительных приборов, или победой врага -- тогда наступает полом или разрушение той или другой части организма. Разрушен орган - выпадает его функция. Но это наш обыкновенный физиологический прием, употребляемый нами для выяснения роли органа, причем часто осуществляемый природой с такой точностью, о которой мы не можем и мечтать с нашими техническими средствами. Если раздражение ограничится одним известным органом, то начинается постепенное возмещение его деятельности, наступает новое уравновешивание организма, вступают деятельность другие замещающие органы. Мы узнаем, таким образом, новые и более тонкие связи органов, скрытые их силы. Если разрушение не остановилось на одном органе, а, цепляясь, распространяется дальше, мы опять еще раз, на новый лад изучаем функциональную связь органов и, наконец, определяем тот момент и механизм, когда истощается объединяющая сила организма как целого. . Разве это с начала и до конца не физиология, углубление в связи и значение частей организма? И только какойнибудь неисправимый схоласт мог бы сказать, что это не наше дело. Напротив, именно физиолог с его компетенцией в методических и логических приемах исследования жизни является здесь самым законным работником.
Могучая сила эксперимента скоро дала себя знать и в новой для нас области - экспериментальной патологии пищеварения.
Только в лице двух работников лаборатория прямо, нарочито обратилась к патологии пищеварения, и, однако, я в состоянии сообщить уже несколько данных, способных, как мне кажется. возбудить интерес и в клиническом мире.
Наши факты пока исключительно относятся только до патологии желудочных желез. Хорошо известный членам нашего собрания способ уединенного желудочка оказался и при патологических исследованиях неоцененным способом как раскрываюший во всей ясности подробности болезненного состояния, так и чрезвычайно способствующий анализу этого болезненного состояния. Если вы болезнетворные условия прикладываете к поверхности маленького желудочка (энергическое тепло, холод, разные химически сильно действующие вещества и т. д.), то вы наблюдаете отклонение желез от нормы в идеально чистом виде, от вас не ускользает ни одна капля болезненно измененного секрета стенки, для вас не затуманивается ни одна, самая незначительная черта болезненного ее состояния. Вы наблюдаете патологическое состояние, вами вызванное, часы и дни, с начала и до конца, в тс время как во всем остальном пищеварительном канале дела идут обычным порядком и не происходит никаких осложнений со стороны последовательных расстройств остального пищеварительного канала и общего питания. Последнее понятно, так как изолированный желудочек никогда не принимает участия в пищеварительном деле, т. е. он ничего не обрабатывает и от него не идет никаких постоянных раздражений на остальной желудок и кишки, потому что он всегда пуст. Исключение представляет только небольшой период времени, когда чрезвычайный раздражитель прикладывается в качестве болезнетворной причины к его стенке и может рефлекторно влиять и на остальной пищеварительный канал. Следовательно, при таких опытах дело почти исключительно идет о знакомстве с патологическим состоянием клеток пепсиновых желез. Если же вы десиствуете болезнетворными причинами на большой желудок, то на маленьком получается возможность или видеть различные рефлекторные влияния, или наблюдать следствие измененного хода пищеварения. Таким образом представляется, между прочим, возможность отличать болезненное состояние рефлекторной поверхности, с одной стороны, и слоя желез - с другой.
Вот наши факты. Если вы вводите в маленький желудочек на минуты сильнодействующие вещества, как: абсолютный винный спирт, раствор сулемы 1 на 500, 10% раствор ляписа, сильная эмульсия горчичного масла, вы вызываете тем более или менее сильное, иногда грандиозное отделение слизи (опыты д-ра Я. X. Завриева). Перед вами, повидимому, серьезное заболевание в виде острого слизистого катара. Но болезнь ли это в самом деле? В крайнем случае слизи льется с нашей поверхности чуть ли не в 100 раз больше нормы. Иногда за весь отделительный период вы получаете только слизь вместо сока. И, однако, спрашиваю еще раз, болезнь ли это? Иногда уже через час, через два это более или менее обильное отделение слизи, наступившее непосредственно после приложения раздражающего агента, уже совершенно изгладилось. В другой раз от чрезвычайного слизетечения, заменившего в день опыта весь отделительный нормальный продукт, на другой день сверх всякого вашего ожидания не остается почти и следа. Поражает контраст между чрезвычайностью явления и его мимолетностью. Напрашивается предположение, что в описанных случаях мы совсем не имели еще дела с болезнью - перед нами разыгралась только победоносная борьба с болезнетворную причиной. В самом деле, не сейчас ли только покровный эпителий желудка развернул перед нами свое настоящее физиологическое назначение, о котором при обыкновенном течении жизни мы не имели бы случая составить себе надлежащее понятие? Своей чрезвычайной работой, изливая массу слизистой жидкости, разбавляющей, связывающей и оттесняющей от стенки введенные вредные вещества, он предотврашает опасность, которою грозят эти вещества важнейшим частям слизистой оболочки. Что наше объяснение верно, с большой достоверностью следует из факта, что при чрезвычайной деятельности покровного эпителия в данном случае эпителий пепсиновых желез остается в совершенном покое. Значит, указанные выше химические вещества раздражают только один эпителий, не трогая другого. Это то же самое, когда мясо, находясь в пищеварительном канале, раздражает только эпителий пепсиновых желез и оставляет совершенно нетронутым покровный эпителий. Мы, как мне кажется, стоим перед весьма важным фактом, что чрезвычайные раздражители, являющиеся в качестве болезнетворных причин, представляют собою специфические раздражители тех защитительных приборов организма, которые назначены для борьбы с соответствующими болезнетворными причинами. Мы думаем, что это представление должно быть обобщено на все случаи болезни, и в этом кроется общий механизм приспособлений организма вообще при встрече с патогенными условиями, совершенно подобно тому, как нормальный, сочетанный и приспособленный ход жизни имеет в своем основании специфическое раздражение того или другого аппарата.
Само собою разумеется, что действие вышеупомянутых агентов, как ни энергично исполняет свою защитительную роль покровный эпителий, может при известном размере коснуться и глубоких слоев слизистой оболочки, и тогда перед нами проходит измененная деятельность пепсиновых желез с большим разнообразием в частностях, смотря по роду агента и большею частью с физическим характером. Это есть, конечно, различные болезненные состояния пепсиновых желез, но вместе с тем и материал к физиологической характеристике этих клеток. Из имеющегося уже значительного запаса относящихся сюда интересных данных я остановлю ваше внимание на одном из них. После приложения 10 % раствора ляписа мы вызывали в пепсиновых железах крайне отчетливое состояние астении - раздражительной слабости (опыты д-ра Я. X. Завриева).
Я представляю в двух столбцах цифр часовые количества сока, изливающегося из изолированного желудочка на одно и то же количество мяса (150 г) до и во время вызванного нами специального заболевания этого желудочка.
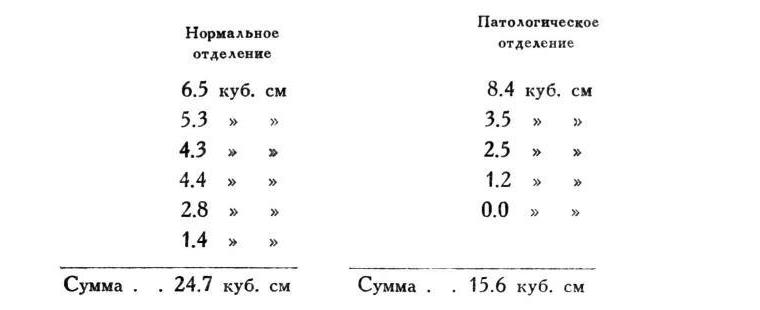 Как вы видите, ход отделения при заболевании приобрел совершенно необычный, своеобразный характер. Величина первого часа отделения отчетливо превосходит нормальный размер, но зато во второй час наблюдается необычно крутое и низкое падение, ниже нормальной величины; то же еще раз повторяется и в третий час -- и отделение обрывается в целом раньше времени, дав в итоге сока гораздо меньше против нормы. Клетка сделалась раздражительней обыкновенного и вместе с тем чрезвычайно утомляемой. Без дальнего ясна важность описанного состояния клетки. Очевидно, состояние это не есть особенность влияния именно ляписа, а должно встречаться и при других условиях, представляя собою одну из типических форм угнетенной деятельности клеток. Можно наверное сказать, что знание этой формы повлияет как на клиническую методику исследования желудка, так и на терапию заболевания. И, сколько я знаю, этот интересный факт впервые открылся лабораторному эксперименту, как ни безмерно, по всей вероятности, велико было число случаев у клиники раньше наблюсти его. Какое разительное доказательство чрезвычайной трудности клинического наблюдения сравнительно с лабораторным!
У У одной из наших собак в изолированном желудочке оказалась круглая язва; она постепенно ширилась, временами давала сильные кровотечения и кончила прободением стенки с последовательным перитонитом и смертью животного (наблюдения и опыты д-ра А. Н. Волковича). Вместе с развитием язвы наблюдалась постепенно развивающаяся гиперсекреция, которая к концу дошла до превышения нормы в 3-4 раза. Но еще больший интерес, чем эта гиперсекреция, возбудил резко измененный ход отделения, особенно характерный при еде хлеба. Как впервые показал доктор П. П. Хижин, хлебное отделение характеризуется тем, что после обильного отделения первого часа во втором наступает сильное падение секреции - чаще всего в 2 раза. Как показал дальнейший анализ, этими двумя часами у хлеба резко разграничиваются обильный психический, центральный период отделения от слабого химического, рефлекторного периода. У нашего животного с круглой язвой первый час отделения ничем не разнился в размере от нормального, но второй теперь оставался равным первому, вместо того чтобы быть вдвое менее, как в норме; также и дальнейшие часы стояли гораздо выше нормы.
Представляю ряды соответствующих цифр.
Как вы видите, ход отделения при заболевании приобрел совершенно необычный, своеобразный характер. Величина первого часа отделения отчетливо превосходит нормальный размер, но зато во второй час наблюдается необычно крутое и низкое падение, ниже нормальной величины; то же еще раз повторяется и в третий час -- и отделение обрывается в целом раньше времени, дав в итоге сока гораздо меньше против нормы. Клетка сделалась раздражительней обыкновенного и вместе с тем чрезвычайно утомляемой. Без дальнего ясна важность описанного состояния клетки. Очевидно, состояние это не есть особенность влияния именно ляписа, а должно встречаться и при других условиях, представляя собою одну из типических форм угнетенной деятельности клеток. Можно наверное сказать, что знание этой формы повлияет как на клиническую методику исследования желудка, так и на терапию заболевания. И, сколько я знаю, этот интересный факт впервые открылся лабораторному эксперименту, как ни безмерно, по всей вероятности, велико было число случаев у клиники раньше наблюсти его. Какое разительное доказательство чрезвычайной трудности клинического наблюдения сравнительно с лабораторным!
У У одной из наших собак в изолированном желудочке оказалась круглая язва; она постепенно ширилась, временами давала сильные кровотечения и кончила прободением стенки с последовательным перитонитом и смертью животного (наблюдения и опыты д-ра А. Н. Волковича). Вместе с развитием язвы наблюдалась постепенно развивающаяся гиперсекреция, которая к концу дошла до превышения нормы в 3-4 раза. Но еще больший интерес, чем эта гиперсекреция, возбудил резко измененный ход отделения, особенно характерный при еде хлеба. Как впервые показал доктор П. П. Хижин, хлебное отделение характеризуется тем, что после обильного отделения первого часа во втором наступает сильное падение секреции - чаще всего в 2 раза. Как показал дальнейший анализ, этими двумя часами у хлеба резко разграничиваются обильный психический, центральный период отделения от слабого химического, рефлекторного периода. У нашего животного с круглой язвой первый час отделения ничем не разнился в размере от нормального, но второй теперь оставался равным первому, вместо того чтобы быть вдвое менее, как в норме; также и дальнейшие часы стояли гораздо выше нормы.
Представляю ряды соответствующих цифр.
 Как понимать это извращение? Мне кажется, наиболее правильное толкование таково: так как отделение первого часа, как центральное, нормально, то это свидетельствует о нормальности желез, центробежных нервов и их центров. Если во второй час мы наблюдаем усиление отделения выше нормы и знаем, что это отделение есть рефлекторное, то приходится признать, что раздраженное состояние в данном случае секреторного аппарата исходит от центростремительных нервов или от их концов. Мы, может быть, знакомимся таким образом с особенным пунктом заболевания, который, сколько я знаю, до сих пор не обособлялся при клиническом исследовании болезни.
При ранних, случайных заболеваниях наших животных приходилось многократно видеть то усиление, то ослабление работы желез против нормы. Много раз казалось, что эти противоположные состояния должны быть фазами одного и того же заболевания. Но что первое, что второе? Наши опыты над нарочитыми заболеваниями как большого, так и маленького желудочка с большим постоянством устанавливали, что первая реакция пепсиновых желез на сильный, чрезвычайный агент есть угнетение их деятельности на несколько часов или даже дней. Это угнетение рефлекторного характера, дело задерживающей нервной системы, приводимой в деятельность чрезвычайным раздражителем. Если влить, например, в большой желудок ледяную воду, раствор ляписа и т. д. (опыты д-ра И. К. Соборова), то отделение затем при нормальной еде окажется задержанным, особенно в первые часы, и в маленьком желудочке, оставшемся совершенно в стороне от болезнетворных воздействий. Можно было бы думать, что желудок при встрече с чрезвычайными раздражителями нарочитым рефлексом всячески тормозит деятельность пепсиновых желез, так сказать, в расчете вернее предохранить от вредного влияния глубоко успокоенную клетку. Единственное исключение из приведенного отношения представляет концентрированный алкоголь. Если алкоголь вливается в большой желудок, то в маленьком наступает обильнейшее выделение сока, то же и наоборот (опыты д-ра Я. Х. Завриева).
Вызывая неоднократно заболевания в большом желудке, мы получили возможность с поразительной ясностью наблюдать компенсаторную деятельность в уединенном желудочке как здоровом участке желудка (опыты д-ра И. К. Соборова). Лишь только в большом происходило уменьшение секреции против нормы, в маленьком сейчас же наблюдалось усиление. Когда мы прекратили совершенно деятельность большого желудка посредством ожога горячей водой, и на много дней, маленький постепенно начал развивать колоссальную деятельность и в конце концов, по крайней мере для некоторых сортов вишни, возместил сполна большой желудок, работая в 10 раз больше против нормы, так как маленький желудочек в норме по размерам секреции составлял 0.1 большого. Обратно, при усилении работы желез в большом желудке, маленький заметно сокращал свою работу. Таким образом при патологических состояниях большого желудка маленький воспроизводит деятельнольшого в обратном виде. Отношение между желудками как в норме, так и при патологических состояниях того или другого устанавливалось на несколько ладов. У голодного животного при открытой фистуле большого желудка поддразниванием вызывалось и измерялось отделение в обоих желудках. Или также голодному животному при открытой фистуле большого желудка в продолжение некоторого времени давали есть мясо или кусками, или, что лучше, в виде жидкого пюре. Затем большой желудок отмывали от остатков пищи и опять из обоих желудков собирали выделяющийся сок. Иногда через 3-4 часа нормального пищеварения открывали фистулу большого желудка, выпускали содержимое, желудок мыли и опять собирали продолжавший вытекать еще некоторое время сок и сравнивали. Наконец часто в конце пищеварения, открывая желудочную фистулу, большой желудок находили уже свободным от пищи, а отделение из него, как и из маленького желудка, еще продолжалось. Pезкий факт викарирующей способности в желудке, очевидно, настойчиво выдвигает чисто физиологическую задачу анализировать механизм этой способности.
Только что перечисленным экспериментально-патологическим фактам я осмеливаюсь приписывать некоторую важность. Мне кажется, что благодаря им болезненное состояние характеризуется полнее, в нем точнее отграничивается защитительнофизиологическое от чисто патологического; само патологическое расчленяется на фазы и, наконец, определеннее локализируется. Я проникнут горячим убеждением, что дальнейшие старания на этом пути увенчаются еще более совершенным успехом и мы будем в состоянии так же полно и осмысленно представить себе болезненные состояния пищеварительного канала, как в последнее время в поражающей красоте развертывается перед нами нормальный ход пищеварительного дела.
Но должны ли мы как экспериментаторы удовлетворяться этим? Не думаю. Не натурально ли, видя отклонение от нормы и глубоко вникнув в их механизм, желать повернуть их к норме? Только это и есть последняя проба полноты вашего физиологического знания и размеров вашей власти над предметом. Следовательно, мы естественно пришли к экспериментальной терапии. Отбросьте практическую цель экспериментальной терапии, останется новый и плодотворный способ изучения жизни, потому что вы будете подходить к изучаемой вами жизни с новой стороны и во всяком случае постоянно будете освещать те прорехи, которые представляет современное физиологическое учение.
Поясню мою мысль примером: механик кончает свое изучение той или другой машины тем, что подвергается экзамену, состоящему в сборе разобранной и спутанной машины. То же должно быть и с физиологом. Только тот может сказать, что он изучил жизнь, кто сумеет вернуть нарушенный ход ее к норме. Еще раз экспериментальная терапия в своей сущности есть проверка физиологии.
Пусть не будет недоразумений. Этими словами об экспериментальной терапии, как и вышеприведенными по поводу патологии, я не говорю, конечно, что-нибудь новое, я - только голос современности. Нет спора, что великая честь действительного объединения всей медицины в эксперименте принадлежит современной бактериологии. Она одновременно и физиология, и патология, и терапия, с начала и до конца экспериментальная. Как молодая и потому сильная научная отрасль, бактериология развернулась во всю натуральную, законную ширь своей идеи, не считаясь с традиционными рамками и шаблонами, которые искусственно разграничивают разработку других, более старых вопросов.
Обращаюсь к нашим экспериментально-терапевтическим опытам. Конечно, это пока еще мелочь, но мы имеем, как мне кажется, законную надежду в ближайшем будущем настоять на этом способе исследования в мере, соответствующей результатам физиологического и экспериментально-патологического исследования. Понятно, на первых порах нами часто руководят клиникотерапевтические указания, однако мы убеждены, что вскоре наша терапия все чаще и чаще будет являться выводом из физиологического и экспериментально-патологического знания, и тогда экспериментальная, лабораторная терапия сама будет указывать клинике, и притом деловито, с полною компетенциею, на целисообразный образ действия.
Как о первом примере нашей терапии позвольте напомнить. об уходе за собаками с перерезанными на шее блуждающими нервами. При внезапном исключении главного секреторного и двигательного нерва желудка у таких животных на первых порах почти совершенно отсутствует пищеварительная работа желудка, и пища поэтому подвергается очень быстрому загниванию, что в свою очередь отягчает положение дела еще больше. Предварительное возбуждение желез всякий раз посредством химического возбудителя (мясной бульон), вместо теперь отсутствующего нормального психического, и систематическое промывание дочиста желудка от остатков предшествующей порции пищи дают возможность скоро справиться с затруднением и установить удовлетворительный порядок дела. Не могу при этом не выйти на минуту из рамок моей темы и еще раз повторить перед вами, 1 [49] что так долго остававшийся темным вопрос о жизни безвагусных собак, наконец, решен физиологиею успешно лишь благодаря завершившейся полноте физиологического анализа механизма нарушений, наступающих после перерезки блуждающих нервов. Вот яркий пример вполне лабораторной, рациональной терапии против тяжелого, смертельного, правда, также лабораторного, нарушения организма. Сделайте желудочную фистулу и через нее, как указано выше, регулируйте пищеварение в желудке, разъедините операциею эзофаготомии полость рта от полости желудка, чтобы и при рвоте содержимое желудка не попадало в рот и легкие, и перерезка блуждающих нервов на шее перестает быть смертельной и вполне совместима с продолжительным и цветущим существованием животного (опыты проф. И. П. Павлова и д-ра П. Э. Качковского). 1 [50]
Возвращаюсь снова к пищеварению. На ряде собак с развитой гиперсекрецией, то наступавшей вследствие самопроизвольного заболевания животных, то вызванной нами нарочно, была применена нами щелочь % раствор соды) как лекарство. И мы имели удовлетворение видеть полное оправдание того представления о лечебном действии щелочей, которое я развивал 5 лет тому назад перед почтенным собранием и которое так разнилось от ходячего и до сих пор еще упорствующего клинического мнения о щелочах. Все наши гиперсекреции (опыты проф. И. П. Павлова и д-ра И. К. Соборова) сейчас же поддавались действию щелочи и резко сокращались, а систематическое применение щелочи навсегда и совершенно изглаживало чрезмерную раздражительность желез.
Нельзя не обратить внимания на то, что с экспериментальной установкой астении желудочных желез для употребления щелочей рациональное показание расширяется еще больше. Ясно, что при раздражительной слабости клетки, т. е. при чрезвычайной раздражительности, быстро ведущей к истощению, щелочь с своим тормозящим влиянием окажется здесь очень кстати. Конечно, предстоит задача физиологически анализировать механизм действия щелочей.
Мы пробовали уже (опыты д-ра Соборова) и сейчас исследуем тщательно влияние отдельных сортов еды на гиперсекрецию. Судя по всему имеющемуся, гиперсекреторное состояние желудочных желез легко поддается лечебным мерам. Главнейшая трудность, очевидно, будет заключаться в победе над различными видами гиперсекреции.
Мы пытались также помогать и ослабленной деятельности желудочных желез, создавая благоприятные условия для выработки сока. Одно из этих условий мы видели в введении большого количества воды в организм (опыты д-ра Завриева). Последнее основывалось на фактах резкой зависимости количества сока от количества воды в теле (опыты проф. Павлова). Ясно, что изготовление сока железами прежде всего есть притяжение отделительными клетками воды из крови. Этому притяжению кровь с своей стороны в известных случаях представляет значительное препятствие. Раз воды в теле недостаточно, клетки или мало, или совсем не могут забирать нужную им для соков воду. Следовательно, обратно - я могу помочь слабо действующей секреторной клетке, с трудом притягивающей воду из крови, тем, что нарочно разжижу кровь, придам ей лишнюю воду, которую она не только не удерживает, а, наоборот, старается отдать от себя. Наши опыты дали известное подтверждение этим соображениям, но не могут еще считаться законченными.
Теперь обращаюсь к медицине. Как ни исключительно сложны биологические явления среди других явлений природы, как ни трудно поэтому установление истинной причинности между ними, а следовательно и достижение власти над ними, и, однако, непреодолимым велением жизни медицина еще в отдаленные времена призвана была управлять этими явлениями призвана, стало быть, раньше всякого естественно-научного их изучения. И она достигала, по крайней мере до некоторой степени, того, что от нее требовалось. Задача, казалось, так бесконечно велика и безнадежно тяжела вместе с тем все же хоть отчасти решена. Среди бесчисленного множества возможных решений все же захвачены некоторые счастливые разгадки. Этот невероятный успех был возможен только благодаря двум чрезвычайным условиям: непрерывному, со дня первого человека, и страстному стремлению людей к здоровью и жизни и - второе участию в этой работе массы людей, почти всех людей. Но если достигнутое сейчас медициной поражает ум, зато, - кто будет спорить с этим? - оно очень, очень мало сравнительно с тем, чем будет медицина в конце концов. Произойдет это, однако, не через то только, что медицина, можно сказать, ежеминутно пользуется и будет пользоваться, чем дальше, тем больше, для своих распознавательных и лечебных действий всем тем, что дают общие успехи естествознания. Оставаясь только практической, медицина едва ли могла бы достигнуть полного триумфа, потому что по существу дела обречена в большей части своей деятельности на применение лишь одного орудия естествознания - наблюдения, другое - опыт - употребляется ею только с крайнею осторожностью, в относительно узких пределах. Но наблюдение - метод вполне достаточный для изучения только более простых явлений. Чем сложнее явление, - а что сложнее жизни? - тем неизбежнее опыт. Только опыт, ничем, кроме естественных размеров изобретательности ума человеческого, не ограниченный опыт завершит, увенчает дело медицины. Наблюдение видит в животном организме массу явлений, существующих рядом и связанных друг с другом то существенно, то косвенно, то случайно. Ум должен догадаться насчет действительного характера связи и это при множестве возможных предположений. Опыт как бы берет явления в свои руки и пускает в ход то одно, то другое и таким образом в искусственных, упрощенных комбинациях определяет истинную связь между явлениями. Иначе сказать, наблюдение собирает то, что ему предлагает природа, опыт же берет у природы то, что он хочет. И сила биологического опыта поистине колоссальна. Этот опыт в какие-нибудь 70-80 лет создал чуть не всю современную и огромную физиологию органов сложного животного. Всякий образованный человек, если он незнаком еще с биологией, повидав обыкновенный, только несколько старательней обставленный курс демонстративной физиологии животных для медиков, наверное, будет повергнут в крайнее изумление тою властию, которая обнаружится перед ним в руках современного физиолога над сложным организмом животного. И это изумление его еще больше возрастет, когда он заметит, что эта власть - дело не тысячелетий или столетий, а только десятков лет.
Этот победоносный опыт на наших глазах сейчас распространяет свою силу как на патологию, так и на терапию. Нельзя представить себе, почему бы он не сохранил этой силы в новых областях. Мне кажется, капитальнейший успех современной медицины в том и заключается, что она получила возможность в настоящее время вся, во всех ее главнейших сторонах, разрабатываться экспериментально. Чрезвычайное движение этому перевороту сообщила опять же бактериология. Хотя еще за несколько времени перед нею патология перешла в лабораторию, но огромным тормозом при экспериментально-патологическом исследовании являлось влияние неведомой, но огромной болезнетворной причины в виде мира микроорганизмов. У нас имелись под руками все мертвые условия болезни, как механическая сила, тепло и т. д., но у нас не было огромной живой причины в виде микроорганизмов. Лишь с открытием болезнетворных организмов развернулась перед экспериментатором вся область патологической физиологии, и в настоящее время уже ничто не мешает иметь в лаборатории почти весь патологический мир для исследования.
Хотя клиника своими тысячелетними трудами тонко уловила образы различных болезней, дала почти полную морфологию патологических состояний, хотя патологическая анатомия, грубая, как и микроскопические и клинические исследования последнего времени, собрала и ежеминутно собирает огромный материал относительно внутренних подробностей болезненного процесса, однако полный анализ, полное значение механизма болезненного процесса с начала и до конца получатся только из рук эксперимента. Одна патологическая анатомия для этого - еще слишком грубый прием, а одна клиника без опыта бессильна вполне проникнуть в сложность явлений. Лишь лабораторный эксперимент способен в общей картине болезни точно отличить то, что составляет защитительные приемы организма и всякое возмещение утраченного от поломов собственно, только он укажет точно сцепление поломов, т. е. первичную порчу и дальнейшие, ею вызванные. А лишь при этом знании и возможна целесообразная и плодотворная помощь болеющему организму и исключается возможность посторонним вмешательством принести иногда вред вместо пользы. Это - с одной стороны. С другой - только эксперимент переберет и оценит все истинные причины болезненного состояния, потому что он начинает с причины, которую нарочито заставляет действовать. Медицина на этом пункте как раз наиболее бессильна; общеизвестно, что этиология - самый слабый отдел медицины. И в самом деле, разве обыкновенно причины болезни не закрадываются и не начинают действовать в организме раньше, чем больной делается объектом медицинского внимания? А знание причин, конечно, существеннейшее дело медицины. Во-первых, только зная причину, можно метко устремляться против нее, а во-вторых, и это еще важнее, - можно не допустить ее до действия, до вторжения организм. Только познав все причины болезней, настоящая медицина превращается в медицину будущего, т. е. в гигиену в широком смысле слова. Ввиду очевидной бесспорности и важности всего этого нельзя не жалеть, что патология как исключительно экспериментальная наука, как патологическая физиология, все еще не заняла всюду подобающего ей места, то являясь в виде прибавки к патологической анатомии, то теряясь в программе общей патологии. Методы патологической анатомии и экспериментальной патологии слишком далеки друг от друга, чтобы практически, при условиях университетской учебной обстановки, могли ужиться в одном лице, в одном помещении на равных, законных правах. Мне кажется, с другой стороны, что в так называемой сейчас общей патологии центр тяжести должен быть положен именно в экспериментальной патологии, в анализе болезненных процессов путем эксперимента, а не в выводах или отвлечениях из фактов частной патологии, что часто представляет собою только перечень на иной лад частнопатологического материала. Едва ли может быть очень большой научный расчет на такую словесную обработку общепатологического материала, когда настоящее время делается так заманчиво плодотворной экспериментальная обработка нахлынувшего в лабораторию мира патологических явлений.
Легко представить себе всю трудность положения врача, когда он, действуя против той или другой болезни, против того или другого симптома известным лечебным приемом, часто совершенно не знает, что этот прием делает в организме и как помогает в данном случае. Какая неверность, неопределенность в действии, какой широкий простор для случайностей! Понятно поэтому стремление клиницистов уяснить себе механизм действия их лечебных орудий, и уже насчитывается несколько десятков лет, как на помощь терапии призван эксперимент, терапевтические приемы переданы в лабораторию и их действия на здоровых животных подвергаются там анализу. Экспериментирование занялось преимущественно химическими медикаментами - отсюда экспериментальная фармакология.
Однако фармаколог мало-помалу отошел от поставленной ему сначала цели, мало или совсем не озабочиваясь, не интересуясь лечебным действием данного вещества. Фармакология естественно превратилась в часть физиологии, изучающую действие химических агентов на животное тело и преследующую свои чисто теоретические цели. Против этого, конечно, по существу дела ничего возражать нельзя. Но благодаря указанному обстоятельству связь современного фармакологического материала с практической медициной, лежавшая, так сказать, в первоначальном проекте фармакологического эксперимента и напоминающая о себе до сих пор в названии науки как учения о лекарствах, стала, по крайней мере для данного момента, во многих случаях слабой, а иногда даже и чисто схоластической. Например, во многих учебниках после изложения физиологического действия тех или других лекарств следуют показания и противопоказания относительно их терапевтического употребления, сплошь и рядом вне всякой связи с изложенным физиологическим действием. Отсюда иногда жалобы на современную фармакологию со стороны врачей. В обоюдных интересах экспериментаторов, как и врачей, фармакология должна пополниться элементами экспериментальной терапии. Имея дело не только со здоровым, но и с больным животным, применяя те или другие лекарства и не только отмечая их действия вообще, но и преследуя как цель излечение больного животного, фармаколог путем анализа для себя расширит и углубит изучение реакций организма на данное химическое соединение, а также и вообще изучение организма, а для врача уяснит настоящее значение и истинный механизм действия терапевтического агента. Требование это, что касается по крайней мере изучения действия медикаментов на больных животных, сознавалось и заявлялось уже давно; препятствием к его осуществлению являлась только трудность иметь в лаборатории соответствующих больных животных - трудность, в настоящее время в значительной степени побежденная благодаря успехам экспериментальной патологии. Лишь при указанном выше слиянии фармакологии с экспериментгльной терапией по всей справедливости рассеются многие терапевтические миражи; с другой стороны, исключится печальная возможность неправильного забраковывания многих средств только потому, что фармакологический анализ в своих опытах над здоровыми животными или не коснулся еще надлежащих пунктов исследования, или совсем не мог с ними встретиться, имея дело только со здоровыми животными. В программе экспериментальной терапии найдет себенатуральное место экспериментальная обработка других терапевтических приемов, кроме химических агентов, которые сейчас остаются поистине без пристанища в обширном курсе медицинского академического преподавания.
Можно с достаточным основанием надеяться, что мы будем свидетелями огромного взрыва интереса среди исследователей, когда болезненные процессы, и помимо бактерийных, подвергнутся в лаборатории смелому, ничем не стесняемому и постоянно всячески контролируемому лечению. А еще более можно быть убежденному в том, что экспериментатор и вне области бактериологии вправе рассчитывать на не меньшие победы, когда в деле лечения с роли объяснителя, как это дело стояло посейчас, перейдет на роль инициатора. Некоторые надеялись сблизить фармакологию с медициной тем, что рекомендовали устраивать и устраивали при фармакологических лабораториях клинические отделения. Но, мне кажется, больше научной логики и больше шансов на практический успех имеют экспериментально-терапевтические лаборатории, чем специально фармакологические клиники. Ведь, все равно, как ни называйте клинику, больной человек и в новой клинике не может сделаться больше экспериментальным объектом, чем во всякой другой. Компетентность же и систематичность в лечении обязательны и для каждого клинического учителя, так как это только и отличает его от обыкновенного практического врача. Каким образом без особенной выгоды для дела будет принесен в жертву или экспериментатор клиницисту, или клиницист экспериментатору, так как беспрерывное и равномерное слияние этих двух родов деятельности едва ли в правиле практически осуществимо.
Заключаю. Только пройдя через огонь эксперимента, вся медицина станет тем, чем быть должна, т. е. сознательной, а следовательно всегда и вполне целесообразно действующей. Доказательство последнему у всех на глазах в современной хирургии. На чем основаны ее блестящие успехи? На полной сознательности действия. Опираясь на пластическую способность организма, обеспеченная в настоящее время антисептикой и асептикой против своего главного врага - микроорганизмов, она третирует свои предмет с чисто механической точки зрения, строго выводя свой образ действия из знания анатомического строения и физиологического значения той или другой части тела.
Я волнуюсь при мысли, насколько мне удалось оживить в вас, сделать деятельным убеждение в чрезвычайной роли эксперимента для достижения практической медициной ее целей. Но если вы имеете такое убеждение, то ваш долг всячески способствовать делу биологического эксперимента не только возможным личным участием, но и живой помощью экспериментаторам в их деятельности. Для успеха биологического, медицинского тоже, эксперимента нужны соответственные люди, место и средства.
Заметьте, господа, существенную разницу между кадрами клиницистов и экспериментаторов. Медицинские научные деятели происходят из всей массы медицинского сословия, и всякий практический врач, раз он обладает умом, энергией и талантом, может проявить свое участие в общей медицинской науке и сложиться в постоянного и крупного медицинского научного деятеля. Экспериментаторы составляют ничтожную кучку любителей: вне специальной обстановки лаборатории почти невозможно появление экспериментатора. Отсюда возникает для вас обязанность возможно благоприятствовать в стенах коллегии, как и в жизни, начинающим приверженцам лаборатории, лабораторной специализациею, до поры до времени, рискованно суживающим свои жиз
ненные шансы.
Известно, что и многие из клиницистов, как терапевтов, так и хирургов, обращаются к лабораторному эксперименту: то ради анализа болезненного процесса, то для уяснения действия терапевтических агентов, то как к пробе проектированного хирургического приема и т. д. И, конечно, это можно только приветствовать. Клиницисты сейчас ближе, чем физиологи, чувствуют потребность в лабораторном анализе тех или других клинических явлений патологического или терапевтического характера, и поэтому они в огромной массе случаев являются в настоящее время инициаторами экспериментально-патологического и экспериментально-терапевтического исследования. Это их большая заслуга есть и будет. Но дело в том, что для них эта деятельность все же стоит на втором плане, наполняя досуг, остающийся от их первой обязанности - лечить больных людей. Лабораторная же работа требует полной преданности, полного посвящения ей сил работника. И я стою на том, что нашим специальностям (экспериментальная патология и экспериментальная терапия) должно быть дано возможно благоприятное и вполне самостоятельное положение, так как они при широком понимании дела являются по методу и идее все тою же физиологией. Всюду в курсе медицинских наук должны быть три экспериментальных кафедры физиологии: нормальной, патологической и терапевтической.
Теперь - место. Всякий гуманный человек должен приветствовать появление всевозможных усовершенствованных лечебных заведений - будь то по частной или правительственной инициативе. Эти учреждения, во-первых, дома милосердия, и в них находят свой приют больные, т. е. потерпевшие в борьбе за жизнь больший или меньший урон, жертвы жизни. Во-вторых, это арена для деятельности лиц, призванных нести непосильную жизненную ношу - решать часто неразрешенные пока еще задачи. Я не злоупотребляю, господа, словами в последнем случае, я представляю жизнь c ee великим свойством приспособления и меру среднего человека. Но это так в лучшем случае, который только и должен привлекать наше внимание. Я представляю себе человека, умом измеряющего недосягаемую глубину задачи, а сердцем часто чувствующего горечь бессилия. Дайте ему все, что только можете, и все же это будет очень мало.
И однако, господа, роскошное больничное учреждение - все же дань горю и часто бессилию человеческому. Но тогда достоинство человека, гордость человека требует дворцов для приложения силы и обнаружения власти ума человеческого. И сильные передовые нации строят эти дворцы. Например, в Германии происходит как бы турнир роскоши между научными лабораториями и специально физиологическими. К. великому сожалению, далеко не то у нас, кроме одного выдающегося примера, одолженного своим осуществлением высокой мысли и просвещенной благотворительности творца Института эксперимонтальной медицины. Во многих других наших медицинских учреждениях чувствуется значительная недостаточность помещения, стоящая в противоречии с чрезвычайно разрастающимися задачами биологического эксперимента. Помимо огромного числа отдельных специальных комнат для производства различных опытов, в настоящее время являются неизбежной необходимостью обширные и разнообразные помещения для разных опытных животных. Я содержу сейчас в лаборатории Института экспериментальной медицины до тридцати оперированных собак, на которых изучается пищеварение и которые должны так содержаться, чтобы их здоровье не оставляло ничего лучшего желать. Едва ли кто возьмет на себя смелость утверждать, что эти животные не нужны или лишни в таком количестве. Это именно обстоятельство, с одной стороны, придает прочность нашим результатам, потому что при малейшем сомнении или подозрении лаборатория во всякое время может повторить, проверить еще и еще старые опыты. С другой - оно чрезвычайно способствует возбуждению и решению дальнейших, новых вопросов. А мои животные были нужны мне только для физиологических опытов. Сколько же может потребоваться животных для экспериментально-патологических и терапевтических опытов, когда процессы будут длиться не только месяцами, но и годами? Какое же плодотворное поле откроется при этих продолжительных наблюдениях животных, убеждают меня и мои случайные наблюдения последних лет. Я сначала отнюдь не имел намерения производить болезни, я только оперировал животных для своих физиологических целей и держал их месяцы и годы, и, однако, сколько болезненных процессов, и часто глубоких, развивалось на моих глазах. Я видел огромную брюшную водянку, восходящий паралич нервной системы, общую разрываемость кровеносных сосудов и т. д.
Итак, для биологического эксперимента требуются сооружения, стоящие многих сотен тысяч рублей, а у нас сплошь и рядом относятся к этому эксперименту с резкой враждебностью. На постройку новой больницы или клиники охотно идут и частные и правительственные лица; потребности и желания экспериментатора встречают по большей части отпор; симпатии его делу ему не видать и не ждать. Вульгарное, часто повторяемое отождествление эксперимента с резаньем собак, глубокой идеи дела - с печальной, но, к сожалению, необходимой обстановкой его достаточно красноречиво свидетельствует об этом. Самим экспериментаторам как проводящим жизнь в лаборатории и не состоящим в постоянном сношении с обществом не представляется возможности повлиять на неблагоприятное мнение общества об эксперименте и экспериментаторах. Это, господа, ваш долг (обращаюсь к врачебной части моих слушателей) помочь нам. Вы ежедневно среди общества с нижних до самых верхних слоев его! Вы связаны с ним интимными связями, присутствуя, участвуя активно в величайших радостях и горестях людей. Ваши слова, что касается науки о здоровье и жизни людей, не могут быть не услышаны. Вам надлежит распространять среди публики мысль о неизбежной необходимости и первостепенной важности в медицине животного эксперимента. Вы должны объяснить окружающим вас, что чем полнее будет продлен опыт на животных, тем менее часто больным придется быть в положении опытных объектов, со всеми печальными последствиями этого. Приведите им хоть такой пример: если бы в свое время было больше сделано опытов с вырезанием щитовидной железы у животных, то не было несчастных опытов над людьми, которым вырезали ради операции зоба дочиста щитовидную железу и которые вследствие этого впадали в непоправимый кретинизм. Пусть узнает от вас публика, что современная медицина вышла из жестоких опытов над людьми. Как известно, наша академическая медицина, что касается до терапевтических средств, широко черпала из народной медицины. А какими людскими жертвами добыта эта последняя, можно судить по теперешним, вовсе не так редким примерам, когда в какой-нибудь деревенской глуши (да и всегда ли только глуши!) от невежественного опыта с тем или другим лечением знахаря или знахарки в жестоких мучениях кончает смертью тот или другой их пациент. Не говорят ли и закон природы и религия о том, что животные созданы на службу человеку, лишь бы не было ненужного и бесполезного мучительства их.
Если для эксперимента нужны большие и специально приспособленные здания, то требуются, понятно, и соответственные средства для ведения дела, как научноео, так и учебного. И в этом отношении мы чрезвычайно отстали от наших западных соседей. Единственный в своем роде в России бюджет физиологической лаборатории Института экспериментальной медицины, в три с половиной раза превосходящий бюджет физиологической лаборатории такого колоссального медицинского учреждения, как Петербургская военно-медицинская академия, сам только что приближается к средним бюджетам германских университетов. Может ли сколько-нибудь широко и вольно развернуться научная и учебная деятельность экспериментальной кафедры при теперешних требованиях на какую-нибудь тысячу рублей? Та же скудость и относительно лабораторного персонала. Например, физиологическая лаборатория Военно-медицинской академии имеет только одного помощника! Какие практические занятия со студентами по физиологии, столь необходимые для непосредственного знакомства с фактами физиологии и для физиологической дисциплины ума будущего врача, возможны при таком составе кафедры? А в то же время, например, в Англии эти занятия поставлены уже широко; довольно сказать, что в тамошних лабораториях имеется по нескольку десятков вращающихся барабанов для студенческих занятий. Наша экспериментальная работа и наше преподавание экспериментальных наук свои лучшие упования возлагают еще на будущее.
Повторяю еще раз: окончательная победа медицины придет только через лабораторный эксперимент. И поэтому я осмеливаюсь предсказывать, что прогресс медицины в той или другой стране, в том или другом ученом или учебном медицинском учреждении будет измеряться тем вниманием, той заботливостью, которыми окружается там экспериментальный отдел медицины.
Я поставил сегодня труды нашей лаборатории, их общую идею, мои мысли об отношении эксперимента к медицине под покровительство высокого имени чествуемого сегодня клинициста. Имею ли я на то право? Я не сделал бы этого, если бы не был убежден в моем праве. Я имел честь в продолжение десяти лет стоять близко к деятельности покойного клинициста в ее лабораторной отрасли. Прошло только 10 лет со дня кончины Сергея Петровича, образ его свеж еще в памяти всех. Это ли не был клиницист, поражавший способностью разгадывать болезни и находить против них наилучшие средства! Его обаяние среди больных, поистине, носило волшебный характер: лечило часто одно его слово, одно посещение больного. Сколько раз приходилось слышать от его учеников-клиницистов печальное признание, что те же рецепты и, повидимому, при подобных же случаях оказывались недействительными у них, делая чудеса в руках учителя. Казалось бы, полное удовлетворение - как внутреннее, так и внешнее - должно было наполнять душу знаменитого клинициста, и, однако, глубокий ум его, не обольщаясь ближайшим успехом, искал ключа к великой загадке: что такое больной человек и как помочь ему в лаборатории, в животном эксперименте. На моих глазах десятки его учеников направлялись им в лабораторию. И эта высокая оценка эксперимента клиницистом составляет, по моему убеждению, не меньшую славу Сергея Петровича, чем его клиническая, известная всей России деятельность.
Как понимать это извращение? Мне кажется, наиболее правильное толкование таково: так как отделение первого часа, как центральное, нормально, то это свидетельствует о нормальности желез, центробежных нервов и их центров. Если во второй час мы наблюдаем усиление отделения выше нормы и знаем, что это отделение есть рефлекторное, то приходится признать, что раздраженное состояние в данном случае секреторного аппарата исходит от центростремительных нервов или от их концов. Мы, может быть, знакомимся таким образом с особенным пунктом заболевания, который, сколько я знаю, до сих пор не обособлялся при клиническом исследовании болезни.
При ранних, случайных заболеваниях наших животных приходилось многократно видеть то усиление, то ослабление работы желез против нормы. Много раз казалось, что эти противоположные состояния должны быть фазами одного и того же заболевания. Но что первое, что второе? Наши опыты над нарочитыми заболеваниями как большого, так и маленького желудочка с большим постоянством устанавливали, что первая реакция пепсиновых желез на сильный, чрезвычайный агент есть угнетение их деятельности на несколько часов или даже дней. Это угнетение рефлекторного характера, дело задерживающей нервной системы, приводимой в деятельность чрезвычайным раздражителем. Если влить, например, в большой желудок ледяную воду, раствор ляписа и т. д. (опыты д-ра И. К. Соборова), то отделение затем при нормальной еде окажется задержанным, особенно в первые часы, и в маленьком желудочке, оставшемся совершенно в стороне от болезнетворных воздействий. Можно было бы думать, что желудок при встрече с чрезвычайными раздражителями нарочитым рефлексом всячески тормозит деятельность пепсиновых желез, так сказать, в расчете вернее предохранить от вредного влияния глубоко успокоенную клетку. Единственное исключение из приведенного отношения представляет концентрированный алкоголь. Если алкоголь вливается в большой желудок, то в маленьком наступает обильнейшее выделение сока, то же и наоборот (опыты д-ра Я. Х. Завриева).
Вызывая неоднократно заболевания в большом желудке, мы получили возможность с поразительной ясностью наблюдать компенсаторную деятельность в уединенном желудочке как здоровом участке желудка (опыты д-ра И. К. Соборова). Лишь только в большом происходило уменьшение секреции против нормы, в маленьком сейчас же наблюдалось усиление. Когда мы прекратили совершенно деятельность большого желудка посредством ожога горячей водой, и на много дней, маленький постепенно начал развивать колоссальную деятельность и в конце концов, по крайней мере для некоторых сортов вишни, возместил сполна большой желудок, работая в 10 раз больше против нормы, так как маленький желудочек в норме по размерам секреции составлял 0.1 большого. Обратно, при усилении работы желез в большом желудке, маленький заметно сокращал свою работу. Таким образом при патологических состояниях большого желудка маленький воспроизводит деятельнольшого в обратном виде. Отношение между желудками как в норме, так и при патологических состояниях того или другого устанавливалось на несколько ладов. У голодного животного при открытой фистуле большого желудка поддразниванием вызывалось и измерялось отделение в обоих желудках. Или также голодному животному при открытой фистуле большого желудка в продолжение некоторого времени давали есть мясо или кусками, или, что лучше, в виде жидкого пюре. Затем большой желудок отмывали от остатков пищи и опять из обоих желудков собирали выделяющийся сок. Иногда через 3-4 часа нормального пищеварения открывали фистулу большого желудка, выпускали содержимое, желудок мыли и опять собирали продолжавший вытекать еще некоторое время сок и сравнивали. Наконец часто в конце пищеварения, открывая желудочную фистулу, большой желудок находили уже свободным от пищи, а отделение из него, как и из маленького желудка, еще продолжалось. Pезкий факт викарирующей способности в желудке, очевидно, настойчиво выдвигает чисто физиологическую задачу анализировать механизм этой способности.
Только что перечисленным экспериментально-патологическим фактам я осмеливаюсь приписывать некоторую важность. Мне кажется, что благодаря им болезненное состояние характеризуется полнее, в нем точнее отграничивается защитительнофизиологическое от чисто патологического; само патологическое расчленяется на фазы и, наконец, определеннее локализируется. Я проникнут горячим убеждением, что дальнейшие старания на этом пути увенчаются еще более совершенным успехом и мы будем в состоянии так же полно и осмысленно представить себе болезненные состояния пищеварительного канала, как в последнее время в поражающей красоте развертывается перед нами нормальный ход пищеварительного дела.
Но должны ли мы как экспериментаторы удовлетворяться этим? Не думаю. Не натурально ли, видя отклонение от нормы и глубоко вникнув в их механизм, желать повернуть их к норме? Только это и есть последняя проба полноты вашего физиологического знания и размеров вашей власти над предметом. Следовательно, мы естественно пришли к экспериментальной терапии. Отбросьте практическую цель экспериментальной терапии, останется новый и плодотворный способ изучения жизни, потому что вы будете подходить к изучаемой вами жизни с новой стороны и во всяком случае постоянно будете освещать те прорехи, которые представляет современное физиологическое учение.
Поясню мою мысль примером: механик кончает свое изучение той или другой машины тем, что подвергается экзамену, состоящему в сборе разобранной и спутанной машины. То же должно быть и с физиологом. Только тот может сказать, что он изучил жизнь, кто сумеет вернуть нарушенный ход ее к норме. Еще раз экспериментальная терапия в своей сущности есть проверка физиологии.
Пусть не будет недоразумений. Этими словами об экспериментальной терапии, как и вышеприведенными по поводу патологии, я не говорю, конечно, что-нибудь новое, я - только голос современности. Нет спора, что великая честь действительного объединения всей медицины в эксперименте принадлежит современной бактериологии. Она одновременно и физиология, и патология, и терапия, с начала и до конца экспериментальная. Как молодая и потому сильная научная отрасль, бактериология развернулась во всю натуральную, законную ширь своей идеи, не считаясь с традиционными рамками и шаблонами, которые искусственно разграничивают разработку других, более старых вопросов.
Обращаюсь к нашим экспериментально-терапевтическим опытам. Конечно, это пока еще мелочь, но мы имеем, как мне кажется, законную надежду в ближайшем будущем настоять на этом способе исследования в мере, соответствующей результатам физиологического и экспериментально-патологического исследования. Понятно, на первых порах нами часто руководят клиникотерапевтические указания, однако мы убеждены, что вскоре наша терапия все чаще и чаще будет являться выводом из физиологического и экспериментально-патологического знания, и тогда экспериментальная, лабораторная терапия сама будет указывать клинике, и притом деловито, с полною компетенциею, на целисообразный образ действия.
Как о первом примере нашей терапии позвольте напомнить. об уходе за собаками с перерезанными на шее блуждающими нервами. При внезапном исключении главного секреторного и двигательного нерва желудка у таких животных на первых порах почти совершенно отсутствует пищеварительная работа желудка, и пища поэтому подвергается очень быстрому загниванию, что в свою очередь отягчает положение дела еще больше. Предварительное возбуждение желез всякий раз посредством химического возбудителя (мясной бульон), вместо теперь отсутствующего нормального психического, и систематическое промывание дочиста желудка от остатков предшествующей порции пищи дают возможность скоро справиться с затруднением и установить удовлетворительный порядок дела. Не могу при этом не выйти на минуту из рамок моей темы и еще раз повторить перед вами, 1 [49] что так долго остававшийся темным вопрос о жизни безвагусных собак, наконец, решен физиологиею успешно лишь благодаря завершившейся полноте физиологического анализа механизма нарушений, наступающих после перерезки блуждающих нервов. Вот яркий пример вполне лабораторной, рациональной терапии против тяжелого, смертельного, правда, также лабораторного, нарушения организма. Сделайте желудочную фистулу и через нее, как указано выше, регулируйте пищеварение в желудке, разъедините операциею эзофаготомии полость рта от полости желудка, чтобы и при рвоте содержимое желудка не попадало в рот и легкие, и перерезка блуждающих нервов на шее перестает быть смертельной и вполне совместима с продолжительным и цветущим существованием животного (опыты проф. И. П. Павлова и д-ра П. Э. Качковского). 1 [50]
Возвращаюсь снова к пищеварению. На ряде собак с развитой гиперсекрецией, то наступавшей вследствие самопроизвольного заболевания животных, то вызванной нами нарочно, была применена нами щелочь % раствор соды) как лекарство. И мы имели удовлетворение видеть полное оправдание того представления о лечебном действии щелочей, которое я развивал 5 лет тому назад перед почтенным собранием и которое так разнилось от ходячего и до сих пор еще упорствующего клинического мнения о щелочах. Все наши гиперсекреции (опыты проф. И. П. Павлова и д-ра И. К. Соборова) сейчас же поддавались действию щелочи и резко сокращались, а систематическое применение щелочи навсегда и совершенно изглаживало чрезмерную раздражительность желез.
Нельзя не обратить внимания на то, что с экспериментальной установкой астении желудочных желез для употребления щелочей рациональное показание расширяется еще больше. Ясно, что при раздражительной слабости клетки, т. е. при чрезвычайной раздражительности, быстро ведущей к истощению, щелочь с своим тормозящим влиянием окажется здесь очень кстати. Конечно, предстоит задача физиологически анализировать механизм действия щелочей.
Мы пробовали уже (опыты д-ра Соборова) и сейчас исследуем тщательно влияние отдельных сортов еды на гиперсекрецию. Судя по всему имеющемуся, гиперсекреторное состояние желудочных желез легко поддается лечебным мерам. Главнейшая трудность, очевидно, будет заключаться в победе над различными видами гиперсекреции.
Мы пытались также помогать и ослабленной деятельности желудочных желез, создавая благоприятные условия для выработки сока. Одно из этих условий мы видели в введении большого количества воды в организм (опыты д-ра Завриева). Последнее основывалось на фактах резкой зависимости количества сока от количества воды в теле (опыты проф. Павлова). Ясно, что изготовление сока железами прежде всего есть притяжение отделительными клетками воды из крови. Этому притяжению кровь с своей стороны в известных случаях представляет значительное препятствие. Раз воды в теле недостаточно, клетки или мало, или совсем не могут забирать нужную им для соков воду. Следовательно, обратно - я могу помочь слабо действующей секреторной клетке, с трудом притягивающей воду из крови, тем, что нарочно разжижу кровь, придам ей лишнюю воду, которую она не только не удерживает, а, наоборот, старается отдать от себя. Наши опыты дали известное подтверждение этим соображениям, но не могут еще считаться законченными.
Теперь обращаюсь к медицине. Как ни исключительно сложны биологические явления среди других явлений природы, как ни трудно поэтому установление истинной причинности между ними, а следовательно и достижение власти над ними, и, однако, непреодолимым велением жизни медицина еще в отдаленные времена призвана была управлять этими явлениями призвана, стало быть, раньше всякого естественно-научного их изучения. И она достигала, по крайней мере до некоторой степени, того, что от нее требовалось. Задача, казалось, так бесконечно велика и безнадежно тяжела вместе с тем все же хоть отчасти решена. Среди бесчисленного множества возможных решений все же захвачены некоторые счастливые разгадки. Этот невероятный успех был возможен только благодаря двум чрезвычайным условиям: непрерывному, со дня первого человека, и страстному стремлению людей к здоровью и жизни и - второе участию в этой работе массы людей, почти всех людей. Но если достигнутое сейчас медициной поражает ум, зато, - кто будет спорить с этим? - оно очень, очень мало сравнительно с тем, чем будет медицина в конце концов. Произойдет это, однако, не через то только, что медицина, можно сказать, ежеминутно пользуется и будет пользоваться, чем дальше, тем больше, для своих распознавательных и лечебных действий всем тем, что дают общие успехи естествознания. Оставаясь только практической, медицина едва ли могла бы достигнуть полного триумфа, потому что по существу дела обречена в большей части своей деятельности на применение лишь одного орудия естествознания - наблюдения, другое - опыт - употребляется ею только с крайнею осторожностью, в относительно узких пределах. Но наблюдение - метод вполне достаточный для изучения только более простых явлений. Чем сложнее явление, - а что сложнее жизни? - тем неизбежнее опыт. Только опыт, ничем, кроме естественных размеров изобретательности ума человеческого, не ограниченный опыт завершит, увенчает дело медицины. Наблюдение видит в животном организме массу явлений, существующих рядом и связанных друг с другом то существенно, то косвенно, то случайно. Ум должен догадаться насчет действительного характера связи и это при множестве возможных предположений. Опыт как бы берет явления в свои руки и пускает в ход то одно, то другое и таким образом в искусственных, упрощенных комбинациях определяет истинную связь между явлениями. Иначе сказать, наблюдение собирает то, что ему предлагает природа, опыт же берет у природы то, что он хочет. И сила биологического опыта поистине колоссальна. Этот опыт в какие-нибудь 70-80 лет создал чуть не всю современную и огромную физиологию органов сложного животного. Всякий образованный человек, если он незнаком еще с биологией, повидав обыкновенный, только несколько старательней обставленный курс демонстративной физиологии животных для медиков, наверное, будет повергнут в крайнее изумление тою властию, которая обнаружится перед ним в руках современного физиолога над сложным организмом животного. И это изумление его еще больше возрастет, когда он заметит, что эта власть - дело не тысячелетий или столетий, а только десятков лет.
Этот победоносный опыт на наших глазах сейчас распространяет свою силу как на патологию, так и на терапию. Нельзя представить себе, почему бы он не сохранил этой силы в новых областях. Мне кажется, капитальнейший успех современной медицины в том и заключается, что она получила возможность в настоящее время вся, во всех ее главнейших сторонах, разрабатываться экспериментально. Чрезвычайное движение этому перевороту сообщила опять же бактериология. Хотя еще за несколько времени перед нею патология перешла в лабораторию, но огромным тормозом при экспериментально-патологическом исследовании являлось влияние неведомой, но огромной болезнетворной причины в виде мира микроорганизмов. У нас имелись под руками все мертвые условия болезни, как механическая сила, тепло и т. д., но у нас не было огромной живой причины в виде микроорганизмов. Лишь с открытием болезнетворных организмов развернулась перед экспериментатором вся область патологической физиологии, и в настоящее время уже ничто не мешает иметь в лаборатории почти весь патологический мир для исследования.
Хотя клиника своими тысячелетними трудами тонко уловила образы различных болезней, дала почти полную морфологию патологических состояний, хотя патологическая анатомия, грубая, как и микроскопические и клинические исследования последнего времени, собрала и ежеминутно собирает огромный материал относительно внутренних подробностей болезненного процесса, однако полный анализ, полное значение механизма болезненного процесса с начала и до конца получатся только из рук эксперимента. Одна патологическая анатомия для этого - еще слишком грубый прием, а одна клиника без опыта бессильна вполне проникнуть в сложность явлений. Лишь лабораторный эксперимент способен в общей картине болезни точно отличить то, что составляет защитительные приемы организма и всякое возмещение утраченного от поломов собственно, только он укажет точно сцепление поломов, т. е. первичную порчу и дальнейшие, ею вызванные. А лишь при этом знании и возможна целесообразная и плодотворная помощь болеющему организму и исключается возможность посторонним вмешательством принести иногда вред вместо пользы. Это - с одной стороны. С другой - только эксперимент переберет и оценит все истинные причины болезненного состояния, потому что он начинает с причины, которую нарочито заставляет действовать. Медицина на этом пункте как раз наиболее бессильна; общеизвестно, что этиология - самый слабый отдел медицины. И в самом деле, разве обыкновенно причины болезни не закрадываются и не начинают действовать в организме раньше, чем больной делается объектом медицинского внимания? А знание причин, конечно, существеннейшее дело медицины. Во-первых, только зная причину, можно метко устремляться против нее, а во-вторых, и это еще важнее, - можно не допустить ее до действия, до вторжения организм. Только познав все причины болезней, настоящая медицина превращается в медицину будущего, т. е. в гигиену в широком смысле слова. Ввиду очевидной бесспорности и важности всего этого нельзя не жалеть, что патология как исключительно экспериментальная наука, как патологическая физиология, все еще не заняла всюду подобающего ей места, то являясь в виде прибавки к патологической анатомии, то теряясь в программе общей патологии. Методы патологической анатомии и экспериментальной патологии слишком далеки друг от друга, чтобы практически, при условиях университетской учебной обстановки, могли ужиться в одном лице, в одном помещении на равных, законных правах. Мне кажется, с другой стороны, что в так называемой сейчас общей патологии центр тяжести должен быть положен именно в экспериментальной патологии, в анализе болезненных процессов путем эксперимента, а не в выводах или отвлечениях из фактов частной патологии, что часто представляет собою только перечень на иной лад частнопатологического материала. Едва ли может быть очень большой научный расчет на такую словесную обработку общепатологического материала, когда настоящее время делается так заманчиво плодотворной экспериментальная обработка нахлынувшего в лабораторию мира патологических явлений.
Легко представить себе всю трудность положения врача, когда он, действуя против той или другой болезни, против того или другого симптома известным лечебным приемом, часто совершенно не знает, что этот прием делает в организме и как помогает в данном случае. Какая неверность, неопределенность в действии, какой широкий простор для случайностей! Понятно поэтому стремление клиницистов уяснить себе механизм действия их лечебных орудий, и уже насчитывается несколько десятков лет, как на помощь терапии призван эксперимент, терапевтические приемы переданы в лабораторию и их действия на здоровых животных подвергаются там анализу. Экспериментирование занялось преимущественно химическими медикаментами - отсюда экспериментальная фармакология.
Однако фармаколог мало-помалу отошел от поставленной ему сначала цели, мало или совсем не озабочиваясь, не интересуясь лечебным действием данного вещества. Фармакология естественно превратилась в часть физиологии, изучающую действие химических агентов на животное тело и преследующую свои чисто теоретические цели. Против этого, конечно, по существу дела ничего возражать нельзя. Но благодаря указанному обстоятельству связь современного фармакологического материала с практической медициной, лежавшая, так сказать, в первоначальном проекте фармакологического эксперимента и напоминающая о себе до сих пор в названии науки как учения о лекарствах, стала, по крайней мере для данного момента, во многих случаях слабой, а иногда даже и чисто схоластической. Например, во многих учебниках после изложения физиологического действия тех или других лекарств следуют показания и противопоказания относительно их терапевтического употребления, сплошь и рядом вне всякой связи с изложенным физиологическим действием. Отсюда иногда жалобы на современную фармакологию со стороны врачей. В обоюдных интересах экспериментаторов, как и врачей, фармакология должна пополниться элементами экспериментальной терапии. Имея дело не только со здоровым, но и с больным животным, применяя те или другие лекарства и не только отмечая их действия вообще, но и преследуя как цель излечение больного животного, фармаколог путем анализа для себя расширит и углубит изучение реакций организма на данное химическое соединение, а также и вообще изучение организма, а для врача уяснит настоящее значение и истинный механизм действия терапевтического агента. Требование это, что касается по крайней мере изучения действия медикаментов на больных животных, сознавалось и заявлялось уже давно; препятствием к его осуществлению являлась только трудность иметь в лаборатории соответствующих больных животных - трудность, в настоящее время в значительной степени побежденная благодаря успехам экспериментальной патологии. Лишь при указанном выше слиянии фармакологии с экспериментгльной терапией по всей справедливости рассеются многие терапевтические миражи; с другой стороны, исключится печальная возможность неправильного забраковывания многих средств только потому, что фармакологический анализ в своих опытах над здоровыми животными или не коснулся еще надлежащих пунктов исследования, или совсем не мог с ними встретиться, имея дело только со здоровыми животными. В программе экспериментальной терапии найдет себенатуральное место экспериментальная обработка других терапевтических приемов, кроме химических агентов, которые сейчас остаются поистине без пристанища в обширном курсе медицинского академического преподавания.
Можно с достаточным основанием надеяться, что мы будем свидетелями огромного взрыва интереса среди исследователей, когда болезненные процессы, и помимо бактерийных, подвергнутся в лаборатории смелому, ничем не стесняемому и постоянно всячески контролируемому лечению. А еще более можно быть убежденному в том, что экспериментатор и вне области бактериологии вправе рассчитывать на не меньшие победы, когда в деле лечения с роли объяснителя, как это дело стояло посейчас, перейдет на роль инициатора. Некоторые надеялись сблизить фармакологию с медициной тем, что рекомендовали устраивать и устраивали при фармакологических лабораториях клинические отделения. Но, мне кажется, больше научной логики и больше шансов на практический успех имеют экспериментально-терапевтические лаборатории, чем специально фармакологические клиники. Ведь, все равно, как ни называйте клинику, больной человек и в новой клинике не может сделаться больше экспериментальным объектом, чем во всякой другой. Компетентность же и систематичность в лечении обязательны и для каждого клинического учителя, так как это только и отличает его от обыкновенного практического врача. Каким образом без особенной выгоды для дела будет принесен в жертву или экспериментатор клиницисту, или клиницист экспериментатору, так как беспрерывное и равномерное слияние этих двух родов деятельности едва ли в правиле практически осуществимо.
Заключаю. Только пройдя через огонь эксперимента, вся медицина станет тем, чем быть должна, т. е. сознательной, а следовательно всегда и вполне целесообразно действующей. Доказательство последнему у всех на глазах в современной хирургии. На чем основаны ее блестящие успехи? На полной сознательности действия. Опираясь на пластическую способность организма, обеспеченная в настоящее время антисептикой и асептикой против своего главного врага - микроорганизмов, она третирует свои предмет с чисто механической точки зрения, строго выводя свой образ действия из знания анатомического строения и физиологического значения той или другой части тела.
Я волнуюсь при мысли, насколько мне удалось оживить в вас, сделать деятельным убеждение в чрезвычайной роли эксперимента для достижения практической медициной ее целей. Но если вы имеете такое убеждение, то ваш долг всячески способствовать делу биологического эксперимента не только возможным личным участием, но и живой помощью экспериментаторам в их деятельности. Для успеха биологического, медицинского тоже, эксперимента нужны соответственные люди, место и средства.
Заметьте, господа, существенную разницу между кадрами клиницистов и экспериментаторов. Медицинские научные деятели происходят из всей массы медицинского сословия, и всякий практический врач, раз он обладает умом, энергией и талантом, может проявить свое участие в общей медицинской науке и сложиться в постоянного и крупного медицинского научного деятеля. Экспериментаторы составляют ничтожную кучку любителей: вне специальной обстановки лаборатории почти невозможно появление экспериментатора. Отсюда возникает для вас обязанность возможно благоприятствовать в стенах коллегии, как и в жизни, начинающим приверженцам лаборатории, лабораторной специализациею, до поры до времени, рискованно суживающим свои жиз
ненные шансы.
Известно, что и многие из клиницистов, как терапевтов, так и хирургов, обращаются к лабораторному эксперименту: то ради анализа болезненного процесса, то для уяснения действия терапевтических агентов, то как к пробе проектированного хирургического приема и т. д. И, конечно, это можно только приветствовать. Клиницисты сейчас ближе, чем физиологи, чувствуют потребность в лабораторном анализе тех или других клинических явлений патологического или терапевтического характера, и поэтому они в огромной массе случаев являются в настоящее время инициаторами экспериментально-патологического и экспериментально-терапевтического исследования. Это их большая заслуга есть и будет. Но дело в том, что для них эта деятельность все же стоит на втором плане, наполняя досуг, остающийся от их первой обязанности - лечить больных людей. Лабораторная же работа требует полной преданности, полного посвящения ей сил работника. И я стою на том, что нашим специальностям (экспериментальная патология и экспериментальная терапия) должно быть дано возможно благоприятное и вполне самостоятельное положение, так как они при широком понимании дела являются по методу и идее все тою же физиологией. Всюду в курсе медицинских наук должны быть три экспериментальных кафедры физиологии: нормальной, патологической и терапевтической.
Теперь - место. Всякий гуманный человек должен приветствовать появление всевозможных усовершенствованных лечебных заведений - будь то по частной или правительственной инициативе. Эти учреждения, во-первых, дома милосердия, и в них находят свой приют больные, т. е. потерпевшие в борьбе за жизнь больший или меньший урон, жертвы жизни. Во-вторых, это арена для деятельности лиц, призванных нести непосильную жизненную ношу - решать часто неразрешенные пока еще задачи. Я не злоупотребляю, господа, словами в последнем случае, я представляю жизнь c ee великим свойством приспособления и меру среднего человека. Но это так в лучшем случае, который только и должен привлекать наше внимание. Я представляю себе человека, умом измеряющего недосягаемую глубину задачи, а сердцем часто чувствующего горечь бессилия. Дайте ему все, что только можете, и все же это будет очень мало.
И однако, господа, роскошное больничное учреждение - все же дань горю и часто бессилию человеческому. Но тогда достоинство человека, гордость человека требует дворцов для приложения силы и обнаружения власти ума человеческого. И сильные передовые нации строят эти дворцы. Например, в Германии происходит как бы турнир роскоши между научными лабораториями и специально физиологическими. К. великому сожалению, далеко не то у нас, кроме одного выдающегося примера, одолженного своим осуществлением высокой мысли и просвещенной благотворительности творца Института эксперимонтальной медицины. Во многих других наших медицинских учреждениях чувствуется значительная недостаточность помещения, стоящая в противоречии с чрезвычайно разрастающимися задачами биологического эксперимента. Помимо огромного числа отдельных специальных комнат для производства различных опытов, в настоящее время являются неизбежной необходимостью обширные и разнообразные помещения для разных опытных животных. Я содержу сейчас в лаборатории Института экспериментальной медицины до тридцати оперированных собак, на которых изучается пищеварение и которые должны так содержаться, чтобы их здоровье не оставляло ничего лучшего желать. Едва ли кто возьмет на себя смелость утверждать, что эти животные не нужны или лишни в таком количестве. Это именно обстоятельство, с одной стороны, придает прочность нашим результатам, потому что при малейшем сомнении или подозрении лаборатория во всякое время может повторить, проверить еще и еще старые опыты. С другой - оно чрезвычайно способствует возбуждению и решению дальнейших, новых вопросов. А мои животные были нужны мне только для физиологических опытов. Сколько же может потребоваться животных для экспериментально-патологических и терапевтических опытов, когда процессы будут длиться не только месяцами, но и годами? Какое же плодотворное поле откроется при этих продолжительных наблюдениях животных, убеждают меня и мои случайные наблюдения последних лет. Я сначала отнюдь не имел намерения производить болезни, я только оперировал животных для своих физиологических целей и держал их месяцы и годы, и, однако, сколько болезненных процессов, и часто глубоких, развивалось на моих глазах. Я видел огромную брюшную водянку, восходящий паралич нервной системы, общую разрываемость кровеносных сосудов и т. д.
Итак, для биологического эксперимента требуются сооружения, стоящие многих сотен тысяч рублей, а у нас сплошь и рядом относятся к этому эксперименту с резкой враждебностью. На постройку новой больницы или клиники охотно идут и частные и правительственные лица; потребности и желания экспериментатора встречают по большей части отпор; симпатии его делу ему не видать и не ждать. Вульгарное, часто повторяемое отождествление эксперимента с резаньем собак, глубокой идеи дела - с печальной, но, к сожалению, необходимой обстановкой его достаточно красноречиво свидетельствует об этом. Самим экспериментаторам как проводящим жизнь в лаборатории и не состоящим в постоянном сношении с обществом не представляется возможности повлиять на неблагоприятное мнение общества об эксперименте и экспериментаторах. Это, господа, ваш долг (обращаюсь к врачебной части моих слушателей) помочь нам. Вы ежедневно среди общества с нижних до самых верхних слоев его! Вы связаны с ним интимными связями, присутствуя, участвуя активно в величайших радостях и горестях людей. Ваши слова, что касается науки о здоровье и жизни людей, не могут быть не услышаны. Вам надлежит распространять среди публики мысль о неизбежной необходимости и первостепенной важности в медицине животного эксперимента. Вы должны объяснить окружающим вас, что чем полнее будет продлен опыт на животных, тем менее часто больным придется быть в положении опытных объектов, со всеми печальными последствиями этого. Приведите им хоть такой пример: если бы в свое время было больше сделано опытов с вырезанием щитовидной железы у животных, то не было несчастных опытов над людьми, которым вырезали ради операции зоба дочиста щитовидную железу и которые вследствие этого впадали в непоправимый кретинизм. Пусть узнает от вас публика, что современная медицина вышла из жестоких опытов над людьми. Как известно, наша академическая медицина, что касается до терапевтических средств, широко черпала из народной медицины. А какими людскими жертвами добыта эта последняя, можно судить по теперешним, вовсе не так редким примерам, когда в какой-нибудь деревенской глуши (да и всегда ли только глуши!) от невежественного опыта с тем или другим лечением знахаря или знахарки в жестоких мучениях кончает смертью тот или другой их пациент. Не говорят ли и закон природы и религия о том, что животные созданы на службу человеку, лишь бы не было ненужного и бесполезного мучительства их.
Если для эксперимента нужны большие и специально приспособленные здания, то требуются, понятно, и соответственные средства для ведения дела, как научноео, так и учебного. И в этом отношении мы чрезвычайно отстали от наших западных соседей. Единственный в своем роде в России бюджет физиологической лаборатории Института экспериментальной медицины, в три с половиной раза превосходящий бюджет физиологической лаборатории такого колоссального медицинского учреждения, как Петербургская военно-медицинская академия, сам только что приближается к средним бюджетам германских университетов. Может ли сколько-нибудь широко и вольно развернуться научная и учебная деятельность экспериментальной кафедры при теперешних требованиях на какую-нибудь тысячу рублей? Та же скудость и относительно лабораторного персонала. Например, физиологическая лаборатория Военно-медицинской академии имеет только одного помощника! Какие практические занятия со студентами по физиологии, столь необходимые для непосредственного знакомства с фактами физиологии и для физиологической дисциплины ума будущего врача, возможны при таком составе кафедры? А в то же время, например, в Англии эти занятия поставлены уже широко; довольно сказать, что в тамошних лабораториях имеется по нескольку десятков вращающихся барабанов для студенческих занятий. Наша экспериментальная работа и наше преподавание экспериментальных наук свои лучшие упования возлагают еще на будущее.
Повторяю еще раз: окончательная победа медицины придет только через лабораторный эксперимент. И поэтому я осмеливаюсь предсказывать, что прогресс медицины в той или другой стране, в том или другом ученом или учебном медицинском учреждении будет измеряться тем вниманием, той заботливостью, которыми окружается там экспериментальный отдел медицины.
Я поставил сегодня труды нашей лаборатории, их общую идею, мои мысли об отношении эксперимента к медицине под покровительство высокого имени чествуемого сегодня клинициста. Имею ли я на то право? Я не сделал бы этого, если бы не был убежден в моем праве. Я имел честь в продолжение десяти лет стоять близко к деятельности покойного клинициста в ее лабораторной отрасли. Прошло только 10 лет со дня кончины Сергея Петровича, образ его свеж еще в памяти всех. Это ли не был клиницист, поражавший способностью разгадывать болезни и находить против них наилучшие средства! Его обаяние среди больных, поистине, носило волшебный характер: лечило часто одно его слово, одно посещение больного. Сколько раз приходилось слышать от его учеников-клиницистов печальное признание, что те же рецепты и, повидимому, при подобных же случаях оказывались недействительными у них, делая чудеса в руках учителя. Казалось бы, полное удовлетворение - как внутреннее, так и внешнее - должно было наполнять душу знаменитого клинициста, и, однако, глубокий ум его, не обольщаясь ближайшим успехом, искал ключа к великой загадке: что такое больной человек и как помочь ему в лаборатории, в животном эксперименте. На моих глазах десятки его учеников направлялись им в лабораторию. И эта высокая оценка эксперимента клиницистом составляет, по моему убеждению, не меньшую славу Сергея Петровича, чем его клиническая, известная всей России деятельность.
 Физиологическая хирургия пищеварительного канала должна давать экспериментатору возможность в любое время получать точные сведения о работе пищеварительное железы, т. е. устанавливать количественные и качественные колебания ее химического продукта - пищеварительного сока; в то же время хирургический метод должен гарантировать сохранение оперированного животного нормальным или лишь очень небольшое отклонение его от нормы. Задачу получения пищеварительных соков для химического исследования в чистом виде и даже в большом количестве в настоящее время следует считать разрешенной окончательно и удачно для всех пищеварительных желез. Напротив, точное изучение всех нормальных условий, при которых соки поступают из желез в пищеварительный канал, наталкивается на три существенные трудности, которые далеко еще не везде и не в полной мере преодолены. Во-первых, нужно иметь возможность непрерывно и с полной точностью следить за отделением данного сока и собирать его для химического исследования, но в то же время общий ход пищеварения не должен нарушаться, несмотря на обеднение всей системы одним из реактивов. Во-вторых, при железах микроскопической величины для добывания чистого сока необходимо изолировать часть выделяющей сок поверхности от остального пищеварительного канала, но эта поверхность в то же время обычно рефлекторно раздражается нормальными возбудителями. В-третьих, при разрезании на части больших желез (такое разрезание часто требуется методикой) или при изоляции частей выделяющей сок поверхности часто перерезаются и нервы желез, сохранение которых необходимо для нормальной их функции. В преодолении этих трудностей методика подвигается вперед лишь медленно; они вызванатомическими условиями и поэтому различны по своей величине в отношении отдельных желез. У слюнных желез, благодаря множественности и попарному расположению этих органов и благодаря простой физиологической роли слюны, отведение секрета одной из двух парных желез легко компенсируется. В желудке можно легко изолировать часть выделяющей поверхности искусственно; притом имеется возможность пользоваться для физиологического исследования только маленькой частью общего количества сока; отнятие этой части лишь мало повлияет на общий ход пищеварения. В отношении поджелудочной железы трудность уже больше, но и здесь открываются различные возможности для ее преодоления; так, например, благодаря присутствию двух выводных протоков, можно разрезать поджелудочную железу на две части. Что касается желчи и, в частности, исследования ее перехода в пищеварительный канал, то задача отведения части тока желчи для экспериментальных целей еще совершенно не затронута.
Сходно обстоит дело со второй трудностью. В отношении больших желез, расположенных вне пищеварительной трубки и связанных с нею выводным протоком, эта трудность совершенно отпадает, так как здесь выделяющая поверхность полностью отделена от поверхности, принимающей раздражение. В желудке обе поверхности совпадают, и все же изолированная для исследования часть желудка может служить для наших целей, потому что секреторный рефлекс имеет в желудке диффузный характер, т. е. раздражение определенного места внутренней поверхности желудка распространяет свое действие и на железы, находящиеся во всех остальных местах. В кишечнике дело обстоит большею частью не так: здесь господствует местное раздражение, местный рефлекс, а потому тут налицо вся трудность задачи наблюдения нормальной работы кишечных желез на изолированном участке кишки.
Что касается последней трудности, то секреторные нервы там, где они известны и проходят в определенных частях желез или в определенных слоях пищеварительной трубки, легко могут быть сохранены, если при отделении избегать повреждения этих частей или слоев. В отношении многих желез, однако, отсутствуют необходимые для этого знания физиологии или же по анатомическим причинам невозможно произвести рассечение желез, не перерезав одновременно нервы.
Физиологическая хирургия пищеварительного канала должна давать экспериментатору возможность в любое время получать точные сведения о работе пищеварительное железы, т. е. устанавливать количественные и качественные колебания ее химического продукта - пищеварительного сока; в то же время хирургический метод должен гарантировать сохранение оперированного животного нормальным или лишь очень небольшое отклонение его от нормы. Задачу получения пищеварительных соков для химического исследования в чистом виде и даже в большом количестве в настоящее время следует считать разрешенной окончательно и удачно для всех пищеварительных желез. Напротив, точное изучение всех нормальных условий, при которых соки поступают из желез в пищеварительный канал, наталкивается на три существенные трудности, которые далеко еще не везде и не в полной мере преодолены. Во-первых, нужно иметь возможность непрерывно и с полной точностью следить за отделением данного сока и собирать его для химического исследования, но в то же время общий ход пищеварения не должен нарушаться, несмотря на обеднение всей системы одним из реактивов. Во-вторых, при железах микроскопической величины для добывания чистого сока необходимо изолировать часть выделяющей сок поверхности от остального пищеварительного канала, но эта поверхность в то же время обычно рефлекторно раздражается нормальными возбудителями. В-третьих, при разрезании на части больших желез (такое разрезание часто требуется методикой) или при изоляции частей выделяющей сок поверхности часто перерезаются и нервы желез, сохранение которых необходимо для нормальной их функции. В преодолении этих трудностей методика подвигается вперед лишь медленно; они вызванатомическими условиями и поэтому различны по своей величине в отношении отдельных желез. У слюнных желез, благодаря множественности и попарному расположению этих органов и благодаря простой физиологической роли слюны, отведение секрета одной из двух парных желез легко компенсируется. В желудке можно легко изолировать часть выделяющей поверхности искусственно; притом имеется возможность пользоваться для физиологического исследования только маленькой частью общего количества сока; отнятие этой части лишь мало повлияет на общий ход пищеварения. В отношении поджелудочной железы трудность уже больше, но и здесь открываются различные возможности для ее преодоления; так, например, благодаря присутствию двух выводных протоков, можно разрезать поджелудочную железу на две части. Что касается желчи и, в частности, исследования ее перехода в пищеварительный канал, то задача отведения части тока желчи для экспериментальных целей еще совершенно не затронута.
Сходно обстоит дело со второй трудностью. В отношении больших желез, расположенных вне пищеварительной трубки и связанных с нею выводным протоком, эта трудность совершенно отпадает, так как здесь выделяющая поверхность полностью отделена от поверхности, принимающей раздражение. В желудке обе поверхности совпадают, и все же изолированная для исследования часть желудка может служить для наших целей, потому что секреторный рефлекс имеет в желудке диффузный характер, т. е. раздражение определенного места внутренней поверхности желудка распространяет свое действие и на железы, находящиеся во всех остальных местах. В кишечнике дело обстоит большею частью не так: здесь господствует местное раздражение, местный рефлекс, а потому тут налицо вся трудность задачи наблюдения нормальной работы кишечных желез на изолированном участке кишки.
Что касается последней трудности, то секреторные нервы там, где они известны и проходят в определенных частях желез или в определенных слоях пищеварительной трубки, легко могут быть сохранены, если при отделении избегать повреждения этих частей или слоев. В отношении многих желез, однако, отсутствуют необходимые для этого знания физиологии или же по анатомическим причинам невозможно произвести рассечение желез, не перерезав одновременно нервы.
 Как уже упомянуто, ввиду того, что имеется несколько слюнных желез и слюну легко заменить жидкой пищей, отведение одной или двух слюнных желез из ротовой полости не вызовет такого значительного изменения в последнее, которое могло бы привести к достойным упоминания количественным или качественным отклонениям от нормы в работе исследуемой железы.
Как уже упомянуто, ввиду того, что имеется несколько слюнных желез и слюну легко заменить жидкой пищей, отведение одной или двух слюнных желез из ротовой полости не вызовет такого значительного изменения в последнее, которое могло бы привести к достойным упоминания количественным или качественным отклонениям от нормы в работе исследуемой железы.


 В техническом отношении при операции следует иметь в виду следующие пункты. Брюшной разрез производится по белой линии и простирается от мечевидного отростка на 7-9 см книзу. До вскрытия полости желудка рекомендуется перетянуть привратник и кардию двумя, конечно стерилизованными, резиновыми трубками. Разрез стенки желудка сначала делается лишь через серозный и мышечный слои. Особенно нужно следить за тем, чтобы разрезы по обе стороны желудка проходили приблизительно параллельно к линии прикрепления брыжейки, так как только при этом условии удается из обрезанного вокруг лоскута получить хорошо сформированный свод. Теперь наружная поверхность слизистой по всей линии разреза свободна, и на. ней ясно видны проходящие в подслизистойв поперечном направлении сосуды. Эти сосуды обкалываются иглой и перевязываются двумя лигатурами, соответственно обоим разошедшимся краям раны. Лишь теперь разрезают слизистую, притом без всякого кровотечения. Сначала полость желудка вскрывают лишь на маленьком пространстве, а затем производят дезинфекцию внутренности желудка, лучше всего наливая в него 0.5 % раствор соляной кислоты. Кажущаяся очень трудной часть операции разрезание слизистой у основания лоскута и отпрепаровка ее по обе стороны - в действительности производится очень легко. Сначала, быстро и легко работая ножом, проводят поверхностный разрез вдоль всего основания лоскута, причем помощник натягивает двумя пинцетами слизистую и расправляет оперирующему путь для разреза. На 2-3 минуты на линию разреза накладывается продолговатый марлевый тампон для остановки кровотечения. Затем еще раз проводят ножом по линии разреза, причем помощник снова поднимает пинцетами края разреза, чтобы натянуть подслизистую; последнюю тогда легко рассечь вплоть до мышечного слоя. Таким образом мы быстро получаем достаточно широкий и свободно отпрепарованный лоскут слизистой с каждой стороны разреза. Опять вкладывается марлевый тампон для остановки кровотечения, а затем, постепенно снимая тампон, охватывают по отдельности и перевязывают более крупные кровоточащие сосуды. Каждому лоскута слизистой (принадлежащему к большому желудку и к будущему маленькому желудку) теперь с помощью швов придают форму куполообразного свода, своей вогнутостью направленного к соответствующей части желудка. Швы накладываются так, чтобы они нигде не прободали слизистую, а захватывали бы у раны желудка серозный и мышечный слои, а у лоскута слизистой - подслизистую. Отмечают концы и середину лоскутов слизистой с помощью временно наложенных пеанов; соединяют каждую сторону лоскутов слизистой с соответствующим краем раны желудка 4-5 швами, причем линию шва начинают у конечной точки лоскута и идут к середине. Затем рана желудка на всем своем протяжении на большом и малом желудке закрывается с помощью швов. При этом оба куполообразные углубления, состоящие из слизистой, образуют плотную перегородку между обеими частями желудка. Отверстие маленького желудка нужно значительно сузить, приблизительно до диаметра карандаша, для предупреждения выпадения, которое в ином случае обязательно произойдет. Операция при некоторой сноровке заканчивается в течение 11/2-2 часов и хорошо переносится собакой.
В первое время после операции часто наблюдаются патологические симптомы, которые могут вызвать, конечно необоснованное, беспокойство: ускорение пульса, отказ от пищи, рвотные движения, иногда явления пареза. Вероятно, эти явления обусловлены рефлекторно натяжением нервов, проходящих в мосте, который соединяет большой желудок с малым. Они особенно ясно проявляются после первых более обильных кормлений и отличаются от сходных явлении, наблюдающихся, за исключением пареза, при перитоните, своим очень изменчивым характером. Они иногда могут становиться очень сильными. Это неприятное осложнение лучше всего предупредить, прикрепив маленький желудок несколькими швами к большому, а также прикрепляя последний к брюшной стенке.
При дальнейшем уходе за собаками много приходится бороться с разъеданием раны желудочным соком, изливающимся в свободное от опытов время наружу. Поэтому хорошо по возможности долго оставлять собаку стоять в станке и вне времени наблюдения и вводить в маленький желудок трубку для оттока. К станку приделывают соответствующим образом разные подпорки для головы и ног, которыми собака пользуется для принятия удобного положения, когда она усталет стоять на ногах. В отношении разъедания раны способ крайне нуждается в усовершенствовании. Может быть, его можно достигнуть, вставив в маленький желудок соответствующую фистульную трубку. Если таковой не имеется, то удобнее всего собирать сок из фистулы, вводя в желудочек резиновую трубки с многочисленными дырочками у одного конца. В другой конец резиновой трубки вставляется точно подходящая к нему трубка, а на уровне брюшной стенки к резиновой трубочке приделывается круг из крепкой резины, немного вдающийся в отверстие фистулы и хорошо закупоривающий ее. Все вместе удерживается в своем положении с помощью маленького градуированного цилиндра, который привязывается под трубочкой с помощью резиновой трубки, обведенной вокруг туловища животного (рис. 5). Без трубочки не следует собирать сок, иначе он быстро начинает переваривать окружность отверстия фистулы. Особенно благоприятным обстоятельством для техники собирания сока является то, что резиновая трубочка сама по себе не действует как раздражитель и не вызывает ни малейшего отделения желудочного сока. Несмотря на все меры предосторожности, у некоторых собак может произойти выпадение желудочка, которое становится все значительнее и в конце концов при случайном повышении внутрибрюшного давления может привести к полному пролапсу. В этом случае необходимо сузить отверстие желудочка посредством резекции клиновидного куска или, в упорных случаях, вскрыв брюшную полость, пришить желудочек к большому желудку или к брюшной стенки; наконец можно также переместить отверстие фистулы желудочка в новое место брюшной стенки.
В техническом отношении при операции следует иметь в виду следующие пункты. Брюшной разрез производится по белой линии и простирается от мечевидного отростка на 7-9 см книзу. До вскрытия полости желудка рекомендуется перетянуть привратник и кардию двумя, конечно стерилизованными, резиновыми трубками. Разрез стенки желудка сначала делается лишь через серозный и мышечный слои. Особенно нужно следить за тем, чтобы разрезы по обе стороны желудка проходили приблизительно параллельно к линии прикрепления брыжейки, так как только при этом условии удается из обрезанного вокруг лоскута получить хорошо сформированный свод. Теперь наружная поверхность слизистой по всей линии разреза свободна, и на. ней ясно видны проходящие в подслизистойв поперечном направлении сосуды. Эти сосуды обкалываются иглой и перевязываются двумя лигатурами, соответственно обоим разошедшимся краям раны. Лишь теперь разрезают слизистую, притом без всякого кровотечения. Сначала полость желудка вскрывают лишь на маленьком пространстве, а затем производят дезинфекцию внутренности желудка, лучше всего наливая в него 0.5 % раствор соляной кислоты. Кажущаяся очень трудной часть операции разрезание слизистой у основания лоскута и отпрепаровка ее по обе стороны - в действительности производится очень легко. Сначала, быстро и легко работая ножом, проводят поверхностный разрез вдоль всего основания лоскута, причем помощник натягивает двумя пинцетами слизистую и расправляет оперирующему путь для разреза. На 2-3 минуты на линию разреза накладывается продолговатый марлевый тампон для остановки кровотечения. Затем еще раз проводят ножом по линии разреза, причем помощник снова поднимает пинцетами края разреза, чтобы натянуть подслизистую; последнюю тогда легко рассечь вплоть до мышечного слоя. Таким образом мы быстро получаем достаточно широкий и свободно отпрепарованный лоскут слизистой с каждой стороны разреза. Опять вкладывается марлевый тампон для остановки кровотечения, а затем, постепенно снимая тампон, охватывают по отдельности и перевязывают более крупные кровоточащие сосуды. Каждому лоскута слизистой (принадлежащему к большому желудку и к будущему маленькому желудку) теперь с помощью швов придают форму куполообразного свода, своей вогнутостью направленного к соответствующей части желудка. Швы накладываются так, чтобы они нигде не прободали слизистую, а захватывали бы у раны желудка серозный и мышечный слои, а у лоскута слизистой - подслизистую. Отмечают концы и середину лоскутов слизистой с помощью временно наложенных пеанов; соединяют каждую сторону лоскутов слизистой с соответствующим краем раны желудка 4-5 швами, причем линию шва начинают у конечной точки лоскута и идут к середине. Затем рана желудка на всем своем протяжении на большом и малом желудке закрывается с помощью швов. При этом оба куполообразные углубления, состоящие из слизистой, образуют плотную перегородку между обеими частями желудка. Отверстие маленького желудка нужно значительно сузить, приблизительно до диаметра карандаша, для предупреждения выпадения, которое в ином случае обязательно произойдет. Операция при некоторой сноровке заканчивается в течение 11/2-2 часов и хорошо переносится собакой.
В первое время после операции часто наблюдаются патологические симптомы, которые могут вызвать, конечно необоснованное, беспокойство: ускорение пульса, отказ от пищи, рвотные движения, иногда явления пареза. Вероятно, эти явления обусловлены рефлекторно натяжением нервов, проходящих в мосте, который соединяет большой желудок с малым. Они особенно ясно проявляются после первых более обильных кормлений и отличаются от сходных явлении, наблюдающихся, за исключением пареза, при перитоните, своим очень изменчивым характером. Они иногда могут становиться очень сильными. Это неприятное осложнение лучше всего предупредить, прикрепив маленький желудок несколькими швами к большому, а также прикрепляя последний к брюшной стенке.
При дальнейшем уходе за собаками много приходится бороться с разъеданием раны желудочным соком, изливающимся в свободное от опытов время наружу. Поэтому хорошо по возможности долго оставлять собаку стоять в станке и вне времени наблюдения и вводить в маленький желудок трубку для оттока. К станку приделывают соответствующим образом разные подпорки для головы и ног, которыми собака пользуется для принятия удобного положения, когда она усталет стоять на ногах. В отношении разъедания раны способ крайне нуждается в усовершенствовании. Может быть, его можно достигнуть, вставив в маленький желудок соответствующую фистульную трубку. Если таковой не имеется, то удобнее всего собирать сок из фистулы, вводя в желудочек резиновую трубки с многочисленными дырочками у одного конца. В другой конец резиновой трубки вставляется точно подходящая к нему трубка, а на уровне брюшной стенки к резиновой трубочке приделывается круг из крепкой резины, немного вдающийся в отверстие фистулы и хорошо закупоривающий ее. Все вместе удерживается в своем положении с помощью маленького градуированного цилиндра, который привязывается под трубочкой с помощью резиновой трубки, обведенной вокруг туловища животного (рис. 5). Без трубочки не следует собирать сок, иначе он быстро начинает переваривать окружность отверстия фистулы. Особенно благоприятным обстоятельством для техники собирания сока является то, что резиновая трубочка сама по себе не действует как раздражитель и не вызывает ни малейшего отделения желудочного сока. Несмотря на все меры предосторожности, у некоторых собак может произойти выпадение желудочка, которое становится все значительнее и в конце концов при случайном повышении внутрибрюшного давления может привести к полному пролапсу. В этом случае необходимо сузить отверстие желудочка посредством резекции клиновидного куска или, в упорных случаях, вскрыв брюшную полость, пришить желудочек к большому желудку или к брюшной стенки; наконец можно также переместить отверстие фистулы желудочка в новое место брюшной стенки.
 Прежде чем пользоваться для наблюдения собаками с желудочком, надо убедиться в том, что желудочек пригоден, т. е. что он в своей работе действительно точно копирует большой желудок. Простой гарантией того, что операция удалась, что сохранилось достаточное количество волокон блуждающего нерва, является установление типичного течения секреции при различной пище: хлебе, мясе и молоке. У некоторых, очень жадных собак, однако, при всякой пище выделяется столько аппетитного сока, что это извращает нормальные типы секреции; у таких собак нужно сравнивать работу большого желудка и желудочка непосредственно. Уже по одной этой причине собаки с желудочком должны всегда иметь фистулу большого желудка. Безупречное состояние желудочка является доказанным лишь в том случае, если в числе различных условий взаимоотношение между работоспособностью большого желудка и желудочка, т. е. взаимоотношение выработанных ими количеств сока, остается постоянным. Эти условия вкратце следующие.
Прежде всего - психическое раздражение собаки. Здесь опыт очень прост. При пустом желудке, как только обе части желудка придут в состояние покоя, т. е выделяют больше кислого сока, собаку дразнят кормом и определяют количество вытекающего из каждой фистулы сока. Хорошо собирать сок в течение долгого времени, так как в течение более короткого периода проявляются неравномерности в опорожнении обеих частей желудка, как и при примешивании слюны к содержимому большого желудка. Во-вторых, возбуждение желудочных желез со стороны желудка мясом или растворами либиховского мясного экстракта. Возбудитель вводят в большой желудок, ждут, когда секреция желудочка приблизится кму максимуму, затем опорожняют большой желудок, промывают его, если нужно, водою и собирают сок из обеих фистул. В-третьих, проверка тормозящего действия жира. Собаке дают мясо с добавлением большого количества масла (25% и более), или жирное мясо (свинина, гусиное мясо), или, наконец, яичный желток; через 1-2 часа, пока еще имеется ясное торможение секреции, открывают желудок, опорожняют и очищают его и затем сравнивают полученные из обеих фистул количества сока. Если во всех этих случаях полученные из обеих частей желудка количества сока остаются постоянном соотношении, то желудочек является идеальной копией большого желудка.
Хирургическая методика дает нам возможность производить многие исследования и наблюдения на совершенно здоровых животных. Но поэтому и при постановке опытов нужно постоянно и серьезно считаться с влиянием психических явлений y собаки. Полное исключение психического влияния является существенным условием каждого серьезного исследования на собаках с желудочными фистулами, так как именно в работе желудочных желез чрезвычайно сильно проявляется психический мо момент. Поэтому нужно вместе с подопытным животным удалиться в хорошо изолированную комнату и тщательно избегать всего, что могло бы вызвать у собаки мысли о пище. Каждый опыт должен отвечать основному требованию - его можно начинать только после того, как секреция долгое время равнялась нулю. Если не соблюдать этого правила, то мы получим неопределенные и даже совершенно искаженные результаты; сам наблюдатель отдает свою работу на произвол случайности. Самым решительным образом следует бороться с утверждением, будто не нужно или невозможно ждать остановки секреции. Если имеется секреция, то налицо и возбуждающая причина, которая когда-нибудь ведь должна перестать действовать. Если не выждать этого момента, то остается нерешенным вопрос: когда он наступает во время опыта? У всех собак, если только у них закончилось пищеварение и если нет никаких других возбуждающих условий, раньше или позже всегда наступает длительная остановка секреции.
Эту же осторожность, конечно, следует соблюдать и при работе на собаках со слюнной Фистулой. Здесь влияние психического возбуждения проявляется еще яснее, так как выделение слюны - это гораздо более подвижный процесс, чем отделение желудочного сока, а потому здесь большею частью гораздо сильнее бросается в глаза непосредственное взаимоотношение между раздражением и работой железы.
Прежде чем пользоваться для наблюдения собаками с желудочком, надо убедиться в том, что желудочек пригоден, т. е. что он в своей работе действительно точно копирует большой желудок. Простой гарантией того, что операция удалась, что сохранилось достаточное количество волокон блуждающего нерва, является установление типичного течения секреции при различной пище: хлебе, мясе и молоке. У некоторых, очень жадных собак, однако, при всякой пище выделяется столько аппетитного сока, что это извращает нормальные типы секреции; у таких собак нужно сравнивать работу большого желудка и желудочка непосредственно. Уже по одной этой причине собаки с желудочком должны всегда иметь фистулу большого желудка. Безупречное состояние желудочка является доказанным лишь в том случае, если в числе различных условий взаимоотношение между работоспособностью большого желудка и желудочка, т. е. взаимоотношение выработанных ими количеств сока, остается постоянным. Эти условия вкратце следующие.
Прежде всего - психическое раздражение собаки. Здесь опыт очень прост. При пустом желудке, как только обе части желудка придут в состояние покоя, т. е выделяют больше кислого сока, собаку дразнят кормом и определяют количество вытекающего из каждой фистулы сока. Хорошо собирать сок в течение долгого времени, так как в течение более короткого периода проявляются неравномерности в опорожнении обеих частей желудка, как и при примешивании слюны к содержимому большого желудка. Во-вторых, возбуждение желудочных желез со стороны желудка мясом или растворами либиховского мясного экстракта. Возбудитель вводят в большой желудок, ждут, когда секреция желудочка приблизится кму максимуму, затем опорожняют большой желудок, промывают его, если нужно, водою и собирают сок из обеих фистул. В-третьих, проверка тормозящего действия жира. Собаке дают мясо с добавлением большого количества масла (25% и более), или жирное мясо (свинина, гусиное мясо), или, наконец, яичный желток; через 1-2 часа, пока еще имеется ясное торможение секреции, открывают желудок, опорожняют и очищают его и затем сравнивают полученные из обеих фистул количества сока. Если во всех этих случаях полученные из обеих частей желудка количества сока остаются постоянном соотношении, то желудочек является идеальной копией большого желудка.
Хирургическая методика дает нам возможность производить многие исследования и наблюдения на совершенно здоровых животных. Но поэтому и при постановке опытов нужно постоянно и серьезно считаться с влиянием психических явлений y собаки. Полное исключение психического влияния является существенным условием каждого серьезного исследования на собаках с желудочными фистулами, так как именно в работе желудочных желез чрезвычайно сильно проявляется психический мо момент. Поэтому нужно вместе с подопытным животным удалиться в хорошо изолированную комнату и тщательно избегать всего, что могло бы вызвать у собаки мысли о пище. Каждый опыт должен отвечать основному требованию - его можно начинать только после того, как секреция долгое время равнялась нулю. Если не соблюдать этого правила, то мы получим неопределенные и даже совершенно искаженные результаты; сам наблюдатель отдает свою работу на произвол случайности. Самым решительным образом следует бороться с утверждением, будто не нужно или невозможно ждать остановки секреции. Если имеется секреция, то налицо и возбуждающая причина, которая когда-нибудь ведь должна перестать действовать. Если не выждать этого момента, то остается нерешенным вопрос: когда он наступает во время опыта? У всех собак, если только у них закончилось пищеварение и если нет никаких других возбуждающих условий, раньше или позже всегда наступает длительная остановка секреции.
Эту же осторожность, конечно, следует соблюдать и при работе на собаках со слюнной Фистулой. Здесь влияние психического возбуждения проявляется еще яснее, так как выделение слюны - это гораздо более подвижный процесс, чем отделение желудочного сока, а потому здесь большею частью гораздо сильнее бросается в глаза непосредственное взаимоотношение между раздражением и работой железы.
 Разрез длиною в 4 -5 см обыкновенно проводится по белой линии и начинается непосредственно у мечевидного отростка; однако разрез с таким же успехом можно делать у наружного края m. rectus abd., причем он начинается у реберной дуги. Обычно бывает совершенно нетрудно прощупать пальцем в глубине правой стороны брюшной полости двенадцатиперстную кишку и поджелудочную железу. Однако иногда, вероятно вследствие вызванной морфием сильной перистальтики, двенадцатиперстная кишка покидает свое обычное положение, и тогда бывает довольно трудно найти ее вслепую. Разумнее всего в таком случае не ощупывать долго брюшную полость, a расширить брюшной разрез настолько, чтобы можно было искать глазами, тем более, что двенадцатиперстная кишка теперь находится совсем впереди, часто в непосредственном соседстве с брюшной раной. Больший выводной проток поджелудочной железы ищут не на той стороне двенадцатиперстной кишки, где доступен желчный проток, а на противоположной стороне, для чего откидывают вытянутую из брюшной полости двенадцатиперстную кишку направо. Выводной проток находят на 1/2 2 см выше того места, где вертикальная часть поджелудочной железы отступает от кишечника. Часто выводной проток скрыт за сосудами, которые нужно перевязать. Так как зачастую найти выводной проток довольно трудно, то Манковский рекомендует вскрыть кишечник с противоположной стороны посредством маленького продольного разреза и отыскать устье выводного протока на слизистой оболочке. После того как проток найден, в его ближайшей окружности отпрепаровывают поджелудочную железу тупым путем от двенадцатиперстной кишки, вводят через образовавшиеся таким образом промежутки между кишечником и железой два полых зонда, перекрещивают их и вырезают ножницами из кишечной стенки продолговато-ромбовидный кусок, в середине которого находится устье протока (рис. 6). Кишечная рана зашивается, а вырезанный ромб пересаживается в брюшную рану. Предварительно, однака, сантиметра на 1'/2--2 выше и ниже выводного протока проводятся две толстых нити под двенадцатиперстной кишкой. При зашивании раны эти нити проводятся через брюшную стенку и служат для того, чтобы фиксировать двенадцатиперстную кишку на брюшной стенке для предупреждения отрыва выводного протока; их удаляют уже на следующий день после операции. Пересаженное таким образом наружу нормальное устье протока поджелудочной железы не зарастает и превосходно функционирует годами.
Однако при этом способе приходилось и приходится еще в теперь бороться со значительными трудностями. Сюда относится: 1. Переваривающее действие сока на кожу, которое, с одной стороны, приводит к острым заболеваниям и кровотечениям, а с другой стороны - к хроническому утолщению кожи и папилломатозным разрастаниям. С тем и другим борются, возможно долго оставляя собаку подвязанной воронкой в станке и заставляя ее вне опыта лежать на подстилке из материала, всасывающего воду (опилки, песок и т. п.), а в дальнейшем область живота часто обмывают и тщательно обсушивают. 2. Дальнейшим недостатком является постоянная потеря сока, рано или поздно влияющая на здоровье животного и даже могущая вызвать егорть. Мивотные иногда заболевают уже через несколько недель после операции, иногда, однако, позже - через несколько месяцев. Заболевание проявляется в отсутствии аппетита и различных нервных явлениях. В более редких случаях наступает острая общая слабость; гораздо чаще наблюдаются явления раздражения двигательной сферы: контрактуры, фибриллярные подергивания, тонические и клонические судороги, при которых может наступить смерть. Хорошим профилактическим средством является режим, ограниченный небольшим количеством молока и хлеба. При первых признаках заболевания или даже при очень повышенном отделении сока, обычно ведущем к заболеванию, очень хорошо действуют 2 -5.0 соды, ежедневно прибавляемой к пище. Еще не выяснено, в чем заключается причина заболевания; имеются указания на органическое заболевание центральной нервной системы. Также не ясно, на чем основывается хорошее действие приведенных мероприятий. Правда, не подлежит сомнению, что на работу желез можно сильно влиять количественно и качественно с помощью известного режима питания и что сода определенным образом тормозит отделение панкреатического сока; но еще требуется точное экспериментальное исследование для того, чтобы вполне понять действие этих средств. Все же можно рассчитывать на то, что, частью благодаря благоприятному индивидуальному предрасположению, частью благодаря применению описанных мероприятий, мы получим собак, которые в продолжение многих месяцев и даже лет будут жить в лаборатории и смогут служить для самых различных исследований на панкреатической железе.
Было бы, однако, неправильно считать такое разрешение задачи идеальным, так как, во-первых, жизни животного подвергается новой специальной опасности, проявляющейся, как только уход становится не вполне внимательным, после чего животное погибает от последствий фистулы. Затем приходится часто довольно долго ждать, пока работа поджелудочной железы примет более или менее постоянный тип. Наконец - и это важнее всего - нельзя считать пищеварение, а значит и работу поджелудочной железы, по крайней мере во вслучаях, нормальной, так как панкреатический сок вряд ли принимает какое-либо участие в пищеварении, потому что через маленький выводной проток в кишечник попадают только незначительные количества сока. Ухудшение пищеварения у собак с панкреатической фистулой почти не проявляется, пока они получают соответствующую пищу, но оно тотчас же обнаруживается, как только собаки получат пищу, очень богатую жиром; наступает расстройство пищеварения, от которого собаки оправляются лишь с трудом. Надо, следовательно, стремиться к тому, чтобы отвести, с целью наблюдения, наружу небольшую часть сока; большая часть его, как при изолированном желудке, должна участвовать в акте пищезарения. Еще лучше было бы меньший ответвленный проток отводить наружу лишь на время наблюдения, на время отдыха вводить его также в кишечник. Это было бы идеальным разрешением задачи.
В 1894 г. Фодера описал способ, который, казалось, подавал некоторую надежду на такое разрешение вопроса. Автор вводит в открытый сбоку большой выводной проток металлическую трубочку особого устройства, котором проживает и проходит наружу через брюшную рану. Трубочка удерживается в выводном протоке посредством двух согнутых под прямым углом крючкообразных отростков, находящихся в просвете протока. Введение трубочки возможно потому, что она состоит из двух половинок, свинчивающихся друг с другом лишь in situ. Хотя автор сообщает о семи собаках, на которых удалась эта операция, - некоторые собаки были убиты через несколько недель после операции с целью исследования фистулы, - все же неясно, оттекал ли панкреатический сок только наружу или также и в кишечник. Автор также не приводит ни в этой, ни в позднейших работах данных, по которым можно было бы составить себе представление о размерах и течении отделения панкреатического сока при пищеварении. Затем отсутствуют какие-либо данные о трудностях при применении рекомендуемого способа и о перспективах успеха операции. В общем работа производит впечатление чего-то законченного скорее в теоретическом, чем в практическом отношении. Если, однако, существенная сторона способа - приживление боковой фистульной трубки в выводном протоке поджелудочной железы и долгое сохранение его проходимости в нормальном направлении - окажется систематически выполнимой, то этим был бы достигнут самый совершенный метод панкреатической фистулы. Опыты, произведенные в моей лаборатории, однако, еще не привели к желаемому результату.
Поэтому мы испробовали другие способы.
Прежде всего по вышеизложенным причинам нам казалось рациональным выбрать для наложения фистулы не большой выводной проток, как обычно, а маленький. Новая операция ничем существенным не отличается от прежней. Из того, что желчный проток и маленький проток поджелудочной железы открываются вместе в кишечник, не проистекает трудностей. После вырезания куска кишечной стенки с сосочком устья - при этом, понятно, желчный проток перерезается поперек -одится рассечь лишь d. choledochus и покрывающую его слизистую кишечника вдоль на протяжении */4--1 см, т. е. создать для него новое устье. При этом осуществляется и разделение обоих выводных протоков. Наложение швов на кишечник и пересадка вырезанного куска кишки производятся обычным путем. Течение этой операции обычно также хорошее. Трудность способа проявляется, однако, в том, что из маленького выводного протока вытекают лишь несколько капель сока или ничего не вытекает: Поэтому пришлось произвести расширение маленького выводного протока. Большой выводной проток перевязывался, и весь сок проходил теперь, при обширном внутреннем анастомозе обоих протоков, через маленький выводной проток. Действительно, количество сока увеличилось до 100 куб. см (2-й час после кормления хлебом). Можно было далее рассчитывать на то, что большой выводной проток, как всегда после перевязки, будет еще регенерировать недели две, тогда как временное расширение маленького выводного протока сохранится. Действительно, через 3-4 недели выделение из маленького протока поджелудочной железы понизилось до маленьких цифр, которые, однако, были слишком малы. Правда, теперь выделялось вообще больше сока, чем прежде, так что при определенном кормлении, например хлебом, можно было собрать несколько кубических сантиметров сока, но этого количества было слишком мало и выделялось оно очень нерегулярно. Лишь после введения 0.5% соляной кислоты почти всегда удавалось получать значительные количества сока. Во всяком случае этот способ заслуживает дальнейшей разработки; возможно, что дальнейшие опыты дадут более благоприятный результат. Так как оперированные таким образом животные совершенно здоровы, не теряют сока и не страдают от разъедания кожи, при введении же соляной кислоты дают достаточную секрецию, то этот способ уже теперь можно рекомендовать для добывания панкреатического сока (не для исследования работы желез).
Другое видоизменение панкреатической фистулы в личной беседе мне предложил проф. Саноцкий; оно, с его согласия, было произведено в лаборатории. Так как поджелудочная железа имеет два выводных протока, то казалось совершенно естественным использовать только часть железы с соответствующим протоком для фистулы. Для этого нужно только наложить обычную фистулу большого протока и перерезать железу выше в поперечном направлении; таким путем становится возможным отвести наружу секрет третьей или четвертой части всей железы. Операция почти не отличается от наложения типичной панкреатической фистулы; она длится только на 5-10 минут дольше того, сколько требуется для перерезки поджелудочной железы на 1 см выше большого выводного протока и обшивания культи железы притянутой складкой сальника; последнее делается для того, чтобы достигнуть скорейшего заживления и избежать зарастания вновь железы. Крупные сосуды железы проходят в продольном направлении и расположены почти вне железы - между поджелудочной железой и двенадцатиперстной кишкой; поэтому-то и легко изолировать и сохранить их. С ними вместе, очевидно, проходят и крупные нервные стволы, откуда вероятно, что условия работы для изолированной части железы останутся нормальными. Первые опыты, поставленные на оперированной таким образом собаке, повидимому, оправдают эти ожидания. Хотя все эти новые видоизменения панкреатической фистулы еще не окончательно выработаны, но все же можно надеяться, что с их помощью скоро удастся достигнуть существенного улучшения современного способа. Собирают панкреатический сок в стеклянную или металлическую воронку, широкое отверстие которой прилежит к брюшной стенке и удерживается на месте с помощью тонких резиновых трубок, обвязанных вокруг туловища животного. Прикрепить воронку с помощью замазки к коже, как при собирании слюны, не удалось. Пока нет пищеварения и вызванного психическим путем отделения желудочного сока, не происходит и выделения панкреатического сока; однако периодически у разных собак в различные сроки - можно наблюдать незначительное самопроизвольное выделение панкреатического сока, продолжающееся около получаса и не зависящее от вышеназванных двух моме. тов (неопубликованные еще опыт д-ра Болдырева из моей лаборатории). Поэтому лучше всего начинать опыт через короткое время после такой волны самостоятельного выделения.
Разрез длиною в 4 -5 см обыкновенно проводится по белой линии и начинается непосредственно у мечевидного отростка; однако разрез с таким же успехом можно делать у наружного края m. rectus abd., причем он начинается у реберной дуги. Обычно бывает совершенно нетрудно прощупать пальцем в глубине правой стороны брюшной полости двенадцатиперстную кишку и поджелудочную железу. Однако иногда, вероятно вследствие вызванной морфием сильной перистальтики, двенадцатиперстная кишка покидает свое обычное положение, и тогда бывает довольно трудно найти ее вслепую. Разумнее всего в таком случае не ощупывать долго брюшную полость, a расширить брюшной разрез настолько, чтобы можно было искать глазами, тем более, что двенадцатиперстная кишка теперь находится совсем впереди, часто в непосредственном соседстве с брюшной раной. Больший выводной проток поджелудочной железы ищут не на той стороне двенадцатиперстной кишки, где доступен желчный проток, а на противоположной стороне, для чего откидывают вытянутую из брюшной полости двенадцатиперстную кишку направо. Выводной проток находят на 1/2 2 см выше того места, где вертикальная часть поджелудочной железы отступает от кишечника. Часто выводной проток скрыт за сосудами, которые нужно перевязать. Так как зачастую найти выводной проток довольно трудно, то Манковский рекомендует вскрыть кишечник с противоположной стороны посредством маленького продольного разреза и отыскать устье выводного протока на слизистой оболочке. После того как проток найден, в его ближайшей окружности отпрепаровывают поджелудочную железу тупым путем от двенадцатиперстной кишки, вводят через образовавшиеся таким образом промежутки между кишечником и железой два полых зонда, перекрещивают их и вырезают ножницами из кишечной стенки продолговато-ромбовидный кусок, в середине которого находится устье протока (рис. 6). Кишечная рана зашивается, а вырезанный ромб пересаживается в брюшную рану. Предварительно, однака, сантиметра на 1'/2--2 выше и ниже выводного протока проводятся две толстых нити под двенадцатиперстной кишкой. При зашивании раны эти нити проводятся через брюшную стенку и служат для того, чтобы фиксировать двенадцатиперстную кишку на брюшной стенке для предупреждения отрыва выводного протока; их удаляют уже на следующий день после операции. Пересаженное таким образом наружу нормальное устье протока поджелудочной железы не зарастает и превосходно функционирует годами.
Однако при этом способе приходилось и приходится еще в теперь бороться со значительными трудностями. Сюда относится: 1. Переваривающее действие сока на кожу, которое, с одной стороны, приводит к острым заболеваниям и кровотечениям, а с другой стороны - к хроническому утолщению кожи и папилломатозным разрастаниям. С тем и другим борются, возможно долго оставляя собаку подвязанной воронкой в станке и заставляя ее вне опыта лежать на подстилке из материала, всасывающего воду (опилки, песок и т. п.), а в дальнейшем область живота часто обмывают и тщательно обсушивают. 2. Дальнейшим недостатком является постоянная потеря сока, рано или поздно влияющая на здоровье животного и даже могущая вызвать егорть. Мивотные иногда заболевают уже через несколько недель после операции, иногда, однако, позже - через несколько месяцев. Заболевание проявляется в отсутствии аппетита и различных нервных явлениях. В более редких случаях наступает острая общая слабость; гораздо чаще наблюдаются явления раздражения двигательной сферы: контрактуры, фибриллярные подергивания, тонические и клонические судороги, при которых может наступить смерть. Хорошим профилактическим средством является режим, ограниченный небольшим количеством молока и хлеба. При первых признаках заболевания или даже при очень повышенном отделении сока, обычно ведущем к заболеванию, очень хорошо действуют 2 -5.0 соды, ежедневно прибавляемой к пище. Еще не выяснено, в чем заключается причина заболевания; имеются указания на органическое заболевание центральной нервной системы. Также не ясно, на чем основывается хорошее действие приведенных мероприятий. Правда, не подлежит сомнению, что на работу желез можно сильно влиять количественно и качественно с помощью известного режима питания и что сода определенным образом тормозит отделение панкреатического сока; но еще требуется точное экспериментальное исследование для того, чтобы вполне понять действие этих средств. Все же можно рассчитывать на то, что, частью благодаря благоприятному индивидуальному предрасположению, частью благодаря применению описанных мероприятий, мы получим собак, которые в продолжение многих месяцев и даже лет будут жить в лаборатории и смогут служить для самых различных исследований на панкреатической железе.
Было бы, однако, неправильно считать такое разрешение задачи идеальным, так как, во-первых, жизни животного подвергается новой специальной опасности, проявляющейся, как только уход становится не вполне внимательным, после чего животное погибает от последствий фистулы. Затем приходится часто довольно долго ждать, пока работа поджелудочной железы примет более или менее постоянный тип. Наконец - и это важнее всего - нельзя считать пищеварение, а значит и работу поджелудочной железы, по крайней мере во вслучаях, нормальной, так как панкреатический сок вряд ли принимает какое-либо участие в пищеварении, потому что через маленький выводной проток в кишечник попадают только незначительные количества сока. Ухудшение пищеварения у собак с панкреатической фистулой почти не проявляется, пока они получают соответствующую пищу, но оно тотчас же обнаруживается, как только собаки получат пищу, очень богатую жиром; наступает расстройство пищеварения, от которого собаки оправляются лишь с трудом. Надо, следовательно, стремиться к тому, чтобы отвести, с целью наблюдения, наружу небольшую часть сока; большая часть его, как при изолированном желудке, должна участвовать в акте пищезарения. Еще лучше было бы меньший ответвленный проток отводить наружу лишь на время наблюдения, на время отдыха вводить его также в кишечник. Это было бы идеальным разрешением задачи.
В 1894 г. Фодера описал способ, который, казалось, подавал некоторую надежду на такое разрешение вопроса. Автор вводит в открытый сбоку большой выводной проток металлическую трубочку особого устройства, котором проживает и проходит наружу через брюшную рану. Трубочка удерживается в выводном протоке посредством двух согнутых под прямым углом крючкообразных отростков, находящихся в просвете протока. Введение трубочки возможно потому, что она состоит из двух половинок, свинчивающихся друг с другом лишь in situ. Хотя автор сообщает о семи собаках, на которых удалась эта операция, - некоторые собаки были убиты через несколько недель после операции с целью исследования фистулы, - все же неясно, оттекал ли панкреатический сок только наружу или также и в кишечник. Автор также не приводит ни в этой, ни в позднейших работах данных, по которым можно было бы составить себе представление о размерах и течении отделения панкреатического сока при пищеварении. Затем отсутствуют какие-либо данные о трудностях при применении рекомендуемого способа и о перспективах успеха операции. В общем работа производит впечатление чего-то законченного скорее в теоретическом, чем в практическом отношении. Если, однако, существенная сторона способа - приживление боковой фистульной трубки в выводном протоке поджелудочной железы и долгое сохранение его проходимости в нормальном направлении - окажется систематически выполнимой, то этим был бы достигнут самый совершенный метод панкреатической фистулы. Опыты, произведенные в моей лаборатории, однако, еще не привели к желаемому результату.
Поэтому мы испробовали другие способы.
Прежде всего по вышеизложенным причинам нам казалось рациональным выбрать для наложения фистулы не большой выводной проток, как обычно, а маленький. Новая операция ничем существенным не отличается от прежней. Из того, что желчный проток и маленький проток поджелудочной железы открываются вместе в кишечник, не проистекает трудностей. После вырезания куска кишечной стенки с сосочком устья - при этом, понятно, желчный проток перерезается поперек -одится рассечь лишь d. choledochus и покрывающую его слизистую кишечника вдоль на протяжении */4--1 см, т. е. создать для него новое устье. При этом осуществляется и разделение обоих выводных протоков. Наложение швов на кишечник и пересадка вырезанного куска кишки производятся обычным путем. Течение этой операции обычно также хорошее. Трудность способа проявляется, однако, в том, что из маленького выводного протока вытекают лишь несколько капель сока или ничего не вытекает: Поэтому пришлось произвести расширение маленького выводного протока. Большой выводной проток перевязывался, и весь сок проходил теперь, при обширном внутреннем анастомозе обоих протоков, через маленький выводной проток. Действительно, количество сока увеличилось до 100 куб. см (2-й час после кормления хлебом). Можно было далее рассчитывать на то, что большой выводной проток, как всегда после перевязки, будет еще регенерировать недели две, тогда как временное расширение маленького выводного протока сохранится. Действительно, через 3-4 недели выделение из маленького протока поджелудочной железы понизилось до маленьких цифр, которые, однако, были слишком малы. Правда, теперь выделялось вообще больше сока, чем прежде, так что при определенном кормлении, например хлебом, можно было собрать несколько кубических сантиметров сока, но этого количества было слишком мало и выделялось оно очень нерегулярно. Лишь после введения 0.5% соляной кислоты почти всегда удавалось получать значительные количества сока. Во всяком случае этот способ заслуживает дальнейшей разработки; возможно, что дальнейшие опыты дадут более благоприятный результат. Так как оперированные таким образом животные совершенно здоровы, не теряют сока и не страдают от разъедания кожи, при введении же соляной кислоты дают достаточную секрецию, то этот способ уже теперь можно рекомендовать для добывания панкреатического сока (не для исследования работы желез).
Другое видоизменение панкреатической фистулы в личной беседе мне предложил проф. Саноцкий; оно, с его согласия, было произведено в лаборатории. Так как поджелудочная железа имеет два выводных протока, то казалось совершенно естественным использовать только часть железы с соответствующим протоком для фистулы. Для этого нужно только наложить обычную фистулу большого протока и перерезать железу выше в поперечном направлении; таким путем становится возможным отвести наружу секрет третьей или четвертой части всей железы. Операция почти не отличается от наложения типичной панкреатической фистулы; она длится только на 5-10 минут дольше того, сколько требуется для перерезки поджелудочной железы на 1 см выше большого выводного протока и обшивания культи железы притянутой складкой сальника; последнее делается для того, чтобы достигнуть скорейшего заживления и избежать зарастания вновь железы. Крупные сосуды железы проходят в продольном направлении и расположены почти вне железы - между поджелудочной железой и двенадцатиперстной кишкой; поэтому-то и легко изолировать и сохранить их. С ними вместе, очевидно, проходят и крупные нервные стволы, откуда вероятно, что условия работы для изолированной части железы останутся нормальными. Первые опыты, поставленные на оперированной таким образом собаке, повидимому, оправдают эти ожидания. Хотя все эти новые видоизменения панкреатической фистулы еще не окончательно выработаны, но все же можно надеяться, что с их помощью скоро удастся достигнуть существенного улучшения современного способа. Собирают панкреатический сок в стеклянную или металлическую воронку, широкое отверстие которой прилежит к брюшной стенке и удерживается на месте с помощью тонких резиновых трубок, обвязанных вокруг туловища животного. Прикрепить воронку с помощью замазки к коже, как при собирании слюны, не удалось. Пока нет пищеварения и вызванного психическим путем отделения желудочного сока, не происходит и выделения панкреатического сока; однако периодически у разных собак в различные сроки - можно наблюдать незначительное самопроизвольное выделение панкреатического сока, продолжающееся около получаса и не зависящее от вышеназванных двух моме. тов (неопубликованные еще опыт д-ра Болдырева из моей лаборатории). Поэтому лучше всего начинать опыт через короткое время после такой волны самостоятельного выделения.
 Но и теперь еще у фистулы желчного пузыря есть один существенный недостаток, состоящий в том, что вся желчь вытекает наружу, тогда как в норме имеется своего рода кругооборот желчи и сама желчь становится возбудителем своего отделения; кроме того, при этих условиях желчь не принимает участия в пищеварении. Поэтому нужно было отвести наружу только часть, и притом постоянную, желчи. А. Чермак предложил соответствующий способ, однако не разработав его достаточно. Этот способ основан на своеобразном расположении желчных путей у собаки, как это видно на прилагаемом грубо схематическое рисунке (рис. 7). Если перевязать d. choledochus в обозначенном на рисунке месте, то часть желчи будет оттекать в пузырь и оттуда наружу; остальная часть будет переходить в кишечник, не собираясь предварительно в резервуаре. Важно установить, что происходит с желчью в этом отделе желчных путей: переводится ли она тотчас же после своего отделения в кишечник; застаивается ли она в желчных путях, чтобы лишь во время пищеварения перейти в кишечник, или же она, может быть, при более значительной задержке все-таки попадает через какие-либо выше расположенные анастомозы систему отведенных в фистулу протоков. На трупе А. Чермак проектировал фистулу, которая еще более отвечала бы идеальным требованиям, предъявляемым к фистуле желчного пузыря. Перевязав одну из ветвей d. choledochus, он делает анастомоз между ним и желчным пузырем. Самый пузырь он разделяет на две половины и пользуется для фистулы только той половиной, в которую открывается пересаженная ветвь d. choledochus. В интересах дела чрезвычайно желательно удачное осуществление автором его проекта.
Но и теперь еще у фистулы желчного пузыря есть один существенный недостаток, состоящий в том, что вся желчь вытекает наружу, тогда как в норме имеется своего рода кругооборот желчи и сама желчь становится возбудителем своего отделения; кроме того, при этих условиях желчь не принимает участия в пищеварении. Поэтому нужно было отвести наружу только часть, и притом постоянную, желчи. А. Чермак предложил соответствующий способ, однако не разработав его достаточно. Этот способ основан на своеобразном расположении желчных путей у собаки, как это видно на прилагаемом грубо схематическое рисунке (рис. 7). Если перевязать d. choledochus в обозначенном на рисунке месте, то часть желчи будет оттекать в пузырь и оттуда наружу; остальная часть будет переходить в кишечник, не собираясь предварительно в резервуаре. Важно установить, что происходит с желчью в этом отделе желчных путей: переводится ли она тотчас же после своего отделения в кишечник; застаивается ли она в желчных путях, чтобы лишь во время пищеварения перейти в кишечник, или же она, может быть, при более значительной задержке все-таки попадает через какие-либо выше расположенные анастомозы систему отведенных в фистулу протоков. На трупе А. Чермак проектировал фистулу, которая еще более отвечала бы идеальным требованиям, предъявляемым к фистуле желчного пузыря. Перевязав одну из ветвей d. choledochus, он делает анастомоз между ним и желчным пузырем. Самый пузырь он разделяет на две половины и пользуется для фистулы только той половиной, в которую открывается пересаженная ветвь d. choledochus. В интересах дела чрезвычайно желательно удачное осуществление автором его проекта.

 Второй тип кишечной фистулы - операция Тири - уже давно известен и так часто применялся, что нет необходимости в точном его описании. Прежде всего назовем здесь также уже давно применявшуюся фистулу по Тири-Велла, при которой в брюшную стенку вшивается не один конец изолированного кишечного цилиндра, как при операции пори, а оба его конца. Для предупреждения выпадения хорошо все только сузить концы кишки, но и скрепить их другом несколькими швами, проводимыми сквозь оба колена кишечной петли.
Дальнейшее видоизменение концевой кишечной (дуоденальной) фистулы, произведенное в моей лаборатории, состоит в том, что верхнее отграничение изолированного отрезка кишки достигается лишь с помощью перегородки из слизистой, так что непрерывность серозного и мышечного слоя не нарушается. Нижний конец изолированного кишечного цилиндра, конечно, образуется путем перерезки всех слоев кишки; он, как обычно, суживается и вшивается в брюшную рану. Для восстановления непрерывности кишечного тракта нижний отдел двенадцатиперстной кишки зашивается наглухо и с помощью бокового анастомоза соединяется над перегородкой из слизистой с двенадцатиперстной кишкой. Операция возникла из предположения, что, может быть, имеются нервни, непосредственно спускающиеся с желудка на двенадцатиперстную кишку. В предпринятых до сих пор опытах не выявилось, однако, различия между отрезком двенадцатиперстной кишки, изолированным по этому способу и по Тири.
Наконец в моей лаборатории применялась еще следующая модификация кишечной фистулы. Длинная изолированная кишечная петля сшивалась своими концами, так что образовалось кишечное кольцо по Герману; в одном месте в кольцо вводилась канюля, которая выводилась наружу при помощи троакара.
Второй тип кишечной фистулы - операция Тири - уже давно известен и так часто применялся, что нет необходимости в точном его описании. Прежде всего назовем здесь также уже давно применявшуюся фистулу по Тири-Велла, при которой в брюшную стенку вшивается не один конец изолированного кишечного цилиндра, как при операции пори, а оба его конца. Для предупреждения выпадения хорошо все только сузить концы кишки, но и скрепить их другом несколькими швами, проводимыми сквозь оба колена кишечной петли.
Дальнейшее видоизменение концевой кишечной (дуоденальной) фистулы, произведенное в моей лаборатории, состоит в том, что верхнее отграничение изолированного отрезка кишки достигается лишь с помощью перегородки из слизистой, так что непрерывность серозного и мышечного слоя не нарушается. Нижний конец изолированного кишечного цилиндра, конечно, образуется путем перерезки всех слоев кишки; он, как обычно, суживается и вшивается в брюшную рану. Для восстановления непрерывности кишечного тракта нижний отдел двенадцатиперстной кишки зашивается наглухо и с помощью бокового анастомоза соединяется над перегородкой из слизистой с двенадцатиперстной кишкой. Операция возникла из предположения, что, может быть, имеются нервни, непосредственно спускающиеся с желудка на двенадцатиперстную кишку. В предпринятых до сих пор опытах не выявилось, однако, различия между отрезком двенадцатиперстной кишки, изолированным по этому способу и по Тири.
Наконец в моей лаборатории применялась еще следующая модификация кишечной фистулы. Длинная изолированная кишечная петля сшивалась своими концами, так что образовалось кишечное кольцо по Герману; в одном месте в кольцо вводилась канюля, которая выводилась наружу при помощи троакара.
 Из этой таблицы (1) вы можете убедиться в полном параллелизме растворяющего и свертывающего действий. Но, может быть, это получилось оттого, что вообще слишком велика разница в ферментном содержании разных проб? Может быть, параллелизм нарушится при меньших разницах? Проследим для этого действие растворяющего и свертывающего белки фермента и по часам отделительного периода (табл. 2).
И в этой таблице (2) мы с отчетливостью видим, что переваривающее и свертывающее действие желудочного сока никогда не расходятся, и в таком случае мысль о тождестве пепсина и химозина получает уже серьезное значение. Ведь, если бы переваривающее и свертывающее действие желудочного сока было обязано двум различным ферментам, несомненно, что эти два действия когда-нибудь разошлись бы.
Из этой таблицы (1) вы можете убедиться в полном параллелизме растворяющего и свертывающего действий. Но, может быть, это получилось оттого, что вообще слишком велика разница в ферментном содержании разных проб? Может быть, параллелизм нарушится при меньших разницах? Проследим для этого действие растворяющего и свертывающего белки фермента и по часам отделительного периода (табл. 2).
И в этой таблице (2) мы с отчетливостью видим, что переваривающее и свертывающее действие желудочного сока никогда не расходятся, и в таком случае мысль о тождестве пепсина и химозина получает уже серьезное значение. Ведь, если бы переваривающее и свертывающее действие желудочного сока было обязано двум различным ферментам, несомненно, что эти два действия когда-нибудь разошлись бы.
 Затем мы обратились к тем научным данным, на которых основано учение о двойственности белковых ферментов. Если действительно мы имеем дело с двумя ферментами, то их надо разделить. Гамарстен, который является одним из видных защитников этой двойственности, находит, что получение чистого пепсина очень легко. Для этого стоит только подкислить настой из слизистой оболочки желудка и поставить в термостат. Через несколько времени весь химозин разрушится и останется только пепсин. Нам необходимо было проделать этот опыт, и мы убедились, что это совсем не так. Мы берем желудочный сок (более чистый, чем настой прежних авторов), определяем его переваривающую и свертывающую силу и следим, что делается с этим соком в термостате. Здесь, однако, для нас возникал технический вопрос: как следить за свертывающим действием желудочного сока изо дня в день? Если получится разница, может быть, эта разница обусловлена не силой фермента, а различной способностью молока свертываться? С этою целью мы простерилизовали сразу много молока и брали отдельные порции его, причем способность его свертываться не меняется. За этой неизменяемостью мы следили все время при помощи другой порции желудочного сока, поставленного на холод, следовательно остающегося без изменения. Этот опыт вполне убедил нас, что в термостате нет никакого преимущественного разрушения свертывающегося фермента, напротив, мы видели вполне параллельное падение как свертывающей, так и растворяющей силы желудочного сока (табл. 3).
Вы видите, что в контрольном опыте свертывающая и переваривающая силы остаются все время одинаковыми, а в соке, находящемся в термостате, и та и другая медленно, но постепенно падают, причем в этом падении наблюдается самый строгий параллелизм. Почему у Гамарстена вышло иначе, судить не берусь.
Второй способ разделения ферментов, как говорит и сам Гамарстен, более труден. Мы не будем передавать вам его в подробности, скажем только, что он основан на расчете, что нерастворимая углемагнезиальная соль увлечет с собою пепсин. С помощью повторного прибавления этой соли и фильтрования в конце концов удается получить такой фильтрат, который не имеет переваривающей силы, в котором остается только химозин. Проверяя этот опыт, мы, однако, не могли получить такого фильтрата, в котором не было бы пепсина. Последний всегда был налицо, но для обнаружения его требовались некоторые приемы. Если взять фильтрат (после повторного фильтрования) и прямо исследовать его ферментативное действие, то мы, действительно, находим только химозин, пепсина же не будет. Но стоит только соответственно разбавить фильтрат подкисленной водой, и пепсинное действие сейчас открывается. Как же объяснить, что при прямом определении открыть его не удается? Вы знаете, что щелочи действуют разрушающим образом на действие пепсина, а углекислая магнезия есть щелочь. Кроме того, она как соль также может задержать пепсинное действие. Очевидно, часть ее растворяется и мешает проявиться этому пепсинному действию. Как только концентрация щелочи уменьшится, это действие выступает с ясностью. Следовательно, и здесь нет того разделения, о котором говорили прежние авторы. Однако мы знаем даже в продаже препараты, которые претендуют на такое разделение ферментного действие, и нам оставалось испытать эти препараты. Мы занялись их исследованием, и я должен сознаться, что первые опыты в этом направлении чрезвычайно поражали. Особенно в одном препарате - рижском сычужке, казалось, существует полное разделение двух ферментов, так как, при сильнейшем свертывающем действии этого препарата на молоко, растворяющее его действие совершенно отсутствовало не только на куриный белок, но и на фибрин. Но на самом деле и это было не так. Стоило нам разбавить этот сычужок в 100 раз 0.2% HCI, и переваривание белков выразилось цифрой 1.3 мм меттовского столбика. Рассчитывая величину по правилу Шюца и Борисова, мы видим, что переваривающая сила пепсина в этом сычужке также максимыльная.
Затем мы обратились к тем научным данным, на которых основано учение о двойственности белковых ферментов. Если действительно мы имеем дело с двумя ферментами, то их надо разделить. Гамарстен, который является одним из видных защитников этой двойственности, находит, что получение чистого пепсина очень легко. Для этого стоит только подкислить настой из слизистой оболочки желудка и поставить в термостат. Через несколько времени весь химозин разрушится и останется только пепсин. Нам необходимо было проделать этот опыт, и мы убедились, что это совсем не так. Мы берем желудочный сок (более чистый, чем настой прежних авторов), определяем его переваривающую и свертывающую силу и следим, что делается с этим соком в термостате. Здесь, однако, для нас возникал технический вопрос: как следить за свертывающим действием желудочного сока изо дня в день? Если получится разница, может быть, эта разница обусловлена не силой фермента, а различной способностью молока свертываться? С этою целью мы простерилизовали сразу много молока и брали отдельные порции его, причем способность его свертываться не меняется. За этой неизменяемостью мы следили все время при помощи другой порции желудочного сока, поставленного на холод, следовательно остающегося без изменения. Этот опыт вполне убедил нас, что в термостате нет никакого преимущественного разрушения свертывающегося фермента, напротив, мы видели вполне параллельное падение как свертывающей, так и растворяющей силы желудочного сока (табл. 3).
Вы видите, что в контрольном опыте свертывающая и переваривающая силы остаются все время одинаковыми, а в соке, находящемся в термостате, и та и другая медленно, но постепенно падают, причем в этом падении наблюдается самый строгий параллелизм. Почему у Гамарстена вышло иначе, судить не берусь.
Второй способ разделения ферментов, как говорит и сам Гамарстен, более труден. Мы не будем передавать вам его в подробности, скажем только, что он основан на расчете, что нерастворимая углемагнезиальная соль увлечет с собою пепсин. С помощью повторного прибавления этой соли и фильтрования в конце концов удается получить такой фильтрат, который не имеет переваривающей силы, в котором остается только химозин. Проверяя этот опыт, мы, однако, не могли получить такого фильтрата, в котором не было бы пепсина. Последний всегда был налицо, но для обнаружения его требовались некоторые приемы. Если взять фильтрат (после повторного фильтрования) и прямо исследовать его ферментативное действие, то мы, действительно, находим только химозин, пепсина же не будет. Но стоит только соответственно разбавить фильтрат подкисленной водой, и пепсинное действие сейчас открывается. Как же объяснить, что при прямом определении открыть его не удается? Вы знаете, что щелочи действуют разрушающим образом на действие пепсина, а углекислая магнезия есть щелочь. Кроме того, она как соль также может задержать пепсинное действие. Очевидно, часть ее растворяется и мешает проявиться этому пепсинному действию. Как только концентрация щелочи уменьшится, это действие выступает с ясностью. Следовательно, и здесь нет того разделения, о котором говорили прежние авторы. Однако мы знаем даже в продаже препараты, которые претендуют на такое разделение ферментного действие, и нам оставалось испытать эти препараты. Мы занялись их исследованием, и я должен сознаться, что первые опыты в этом направлении чрезвычайно поражали. Особенно в одном препарате - рижском сычужке, казалось, существует полное разделение двух ферментов, так как, при сильнейшем свертывающем действии этого препарата на молоко, растворяющее его действие совершенно отсутствовало не только на куриный белок, но и на фибрин. Но на самом деле и это было не так. Стоило нам разбавить этот сычужок в 100 раз 0.2% HCI, и переваривание белков выразилось цифрой 1.3 мм меттовского столбика. Рассчитывая величину по правилу Шюца и Борисова, мы видим, что переваривающая сила пепсина в этом сычужке также максимыльная.

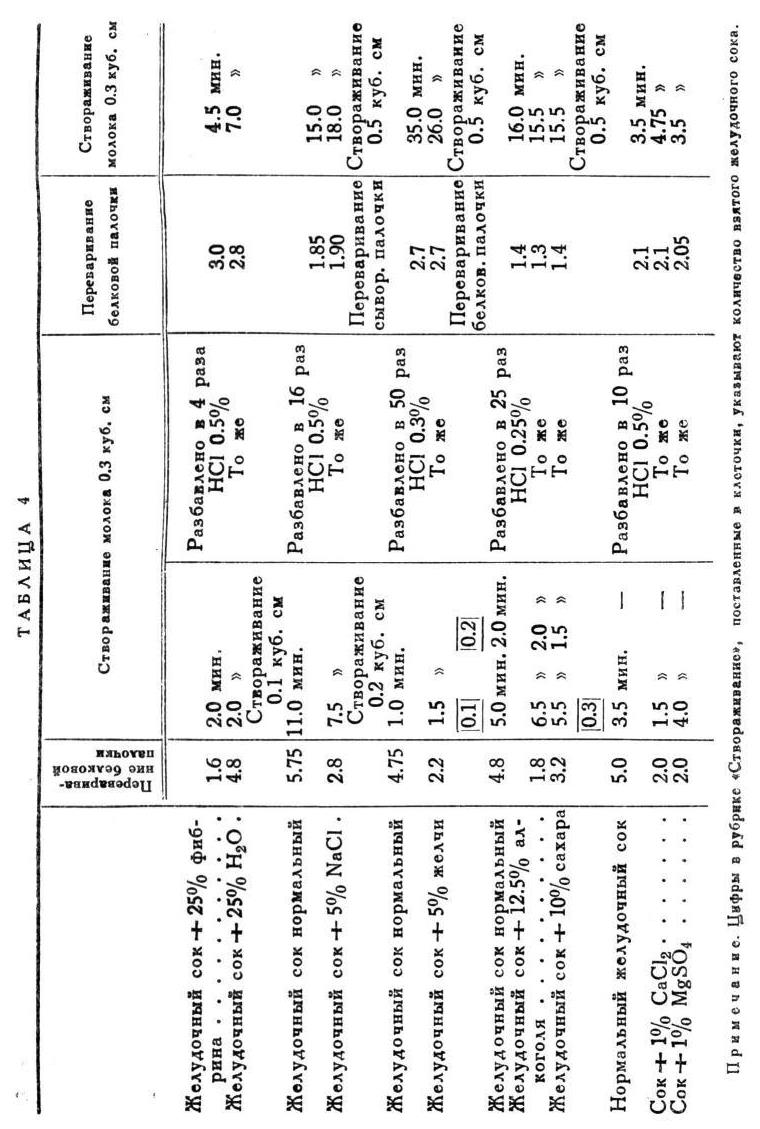 Итак, мы видим полный параллелизм между действием пепсина и химозина и не имеем никаких указаний на разделение этих ферментов.
Почему же прежние исследователи давали противоположные указания? Мы видим эту причину в химических условиях среды. Есть масса таких условий, которые могут направить действие фермента в одну сторону, затормозив его противоположное действие. Чтобы доказать это ad oculos, были произведены также опыты. Если взять желудочный сок и прибавить к нему фибрина. то свертывающая сила его останется без изменения, тогда как переваривающая резко ослабеет. Однако пепсин здесь весь налицо, и в этом можно убедиться, разведя этот сок. Если нормальный (контрольный) сок разбавить во столько же раз, во
Единство пепсина и химозина 343 сколько разбавлен наш сок, то цифры переваривания получатся почти одинаковые. Это давний опыт.
Наши дальнейшие опыты состояли в следующем: прибавка к обыкновенному желудочному соку NaCl уничтожает переваривающую способность его, хотя фермент и здесь также налицо, в чем можно убедиться разведением сока, когда переваривание снова начинается. Совершенно то же получается и при прибавке магнезиальных, известковых солей, желчи и многих других веществ.
Результаты всех этих опытов показывает вам следующая таблица (4).
Итак, мы имеем ряд фактов, которые высокой вероятностью, чтобы не сказать - решающим образом, указывают на то, что и переваривающая и свертывающая белки функции желудочного сока обязаны одному веществу, но могут проявиться отдельно в силу химических свойств среды. При одних условиях оно гидратирует белки, при других - их дегидратирует. В этом едва ли можно сомневаться, тем более, что на других ферментах такая двойственность уже доказана. Особенно ясно это видно на олеопсине. Этот последний разлагает жир на жирные кислоты и глицерин, но и синтезирует их. В настоящее время имеются уже факты, которые указывают, что олеопсин и в органах действует в двух направлениях. Так, в «Journal of Physiology» недавно появилось сообщение, что в молочной железе олеопсина находится так много, что в этом отношении она превосходит поджелудочную. Значит, в поджелудочной железе олеопсин разлагает жиры, в тканях организма он строит, синтезирует жиры.
Эта точка зрения представляет большую научную выгоду, так как вам известны уже указания на нахождение пепсина и трипсина в тканях. Возможно, чтони-то и являются там синтезирующими веществами.
Итак, мы видим полный параллелизм между действием пепсина и химозина и не имеем никаких указаний на разделение этих ферментов.
Почему же прежние исследователи давали противоположные указания? Мы видим эту причину в химических условиях среды. Есть масса таких условий, которые могут направить действие фермента в одну сторону, затормозив его противоположное действие. Чтобы доказать это ad oculos, были произведены также опыты. Если взять желудочный сок и прибавить к нему фибрина. то свертывающая сила его останется без изменения, тогда как переваривающая резко ослабеет. Однако пепсин здесь весь налицо, и в этом можно убедиться, разведя этот сок. Если нормальный (контрольный) сок разбавить во столько же раз, во
Единство пепсина и химозина 343 сколько разбавлен наш сок, то цифры переваривания получатся почти одинаковые. Это давний опыт.
Наши дальнейшие опыты состояли в следующем: прибавка к обыкновенному желудочному соку NaCl уничтожает переваривающую способность его, хотя фермент и здесь также налицо, в чем можно убедиться разведением сока, когда переваривание снова начинается. Совершенно то же получается и при прибавке магнезиальных, известковых солей, желчи и многих других веществ.
Результаты всех этих опытов показывает вам следующая таблица (4).
Итак, мы имеем ряд фактов, которые высокой вероятностью, чтобы не сказать - решающим образом, указывают на то, что и переваривающая и свертывающая белки функции желудочного сока обязаны одному веществу, но могут проявиться отдельно в силу химических свойств среды. При одних условиях оно гидратирует белки, при других - их дегидратирует. В этом едва ли можно сомневаться, тем более, что на других ферментах такая двойственность уже доказана. Особенно ясно это видно на олеопсине. Этот последний разлагает жир на жирные кислоты и глицерин, но и синтезирует их. В настоящее время имеются уже факты, которые указывают, что олеопсин и в органах действует в двух направлениях. Так, в «Journal of Physiology» недавно появилось сообщение, что в молочной железе олеопсина находится так много, что в этом отношении она превосходит поджелудочную. Значит, в поджелудочной железе олеопсин разлагает жиры, в тканях организма он строит, синтезирует жиры.
Эта точка зрения представляет большую научную выгоду, так как вам известны уже указания на нахождение пепсина и трипсина в тканях. Возможно, чтони-то и являются там синтезирующими веществами.
 Из этой таблицы (1) видно, что чем слабее растворяющее действие, тем дольше не наступает свертывание молока.
Тот же самый параллелизм мы видели и в бруннеровском соке [табл. 2].
Из этой таблицы (1) видно, что чем слабее растворяющее действие, тем дольше не наступает свертывание молока.
Тот же самый параллелизм мы видели и в бруннеровском соке [табл. 2].
 Таким образом мы должны признать, что при всех физиологических условиях во всех четырех соках, содержащих белковый фермент, растворяющее и свертывающее действия никогда не расходятся. Но чем более мы убеждались в этом, тем требовательнее становились к себе. Параллелизм обеих функций сока есть факт, но, для того чтобы признать тождество функций, надо иметь не только параллелизм, а и пропорциональность. Такой пропорциональности при наших опытах в первое время не было. Посмотрим следующую таблицу (3).
Таким образом мы должны признать, что при всех физиологических условиях во всех четырех соках, содержащих белковый фермент, растворяющее и свертывающее действия никогда не расходятся. Но чем более мы убеждались в этом, тем требовательнее становились к себе. Параллелизм обеих функций сока есть факт, но, для того чтобы признать тождество функций, надо иметь не только параллелизм, а и пропорциональность. Такой пропорциональности при наших опытах в первое время не было. Посмотрим следующую таблицу (3).
 Параллелизм обеих функций сразу бросается в глаза (табл. 3). Но есть ли пропорциональность? Берем переваривающую силу соков, выраженную в миллиметрах белковой палочки, и, пользуясь правилом Шюца- Борисова, находим, что хлебный сок почти в 5 раз сильнее молочного сока. Обращаясь к свертывающему действию тех же соков, видим, что хлебный сок свертывает молоко в 6/2 раз скорее, чем молочный.
Таким образом параллелизм - налицо, но пропорциональности нет. Но на самом деле это не так: пропорциональность существует, но нужно выбрать такие условия, чтобы ее легко было обнаружить. Ведь мы имеем перед собою две противоположных реакции, идущие при разных условиях и измеряемые по разным правилам.Для растворяющего действия мы имеем правило (Шюца-Борисова), что количество фермента, или его концентрация, прямо пропорционально квадратам миллиметров переваренного белка; для свертывающего действия - правило, что сила сока обратно пропорциональна времени свертывания молока. Каждое правило имеет для своего действия пределы, за которыми оно теряет свой безусловный характе, и может быть принято лишь приблизительно. Вот один из таких пределов: если молоко не свернется в течение десяти минут, мы - за пределом. В этом случае свертывание наступит гораздо позже, чем оно должно бы быть по правилу.
Поэтому, для того чтобы обнаружить пропорциональность свертывающего и растворяющего действия сока, мы, определив количественное отношение ферментов в соках по растворяющему действию, брали затем для свертывания молока эквивалентные по содержанию фермента количества сравниваемых соков. Поясню примером. В предыдущей таблице молочный сок оказался почти в 5 раз слабее хлебного (правило Шюца - Борисова). Если я беру для свертывания молока 0.1 куб. см хлебного сока, то эквивалентное ему количество молочного сока будет в 5 раз больше, т. е. 0.5 куб. см. Но этого недостаточно: взяв в 5 раз больше молочного сока, я, конечно, взял больше и кислоты. Поэтому мы уравниваем кислотность прибавлением к 0.1 куб. см хлебного сока известного количества HCI, точно определенного титрованием. Теперь уже мы имеем два раствора соков, эквивалентные как по количеству содержащегося в них: фермента, так и по кислотности. Но и это еще не все. Представьте, что два соответственных эквивалентных раствора с одинаковым содержанием фермента и с уравненной кислотностью будут разниться по количеству куб. сантиметров, например 1.0 и 0.5 куб. см. Приливая 1.0 куб. см раствора сока к 10.0 куб. см молока, мы больше разжижаем молоко, чем в другой пробирке, где к 10.0 куб. см молока прилито 0.5 куб. см сока. Чтобы уравнять все условия, мы должны уравнять и разведение. С этой целью мы в последнем случае доводим количество раствора сока с 0.5 куб. см до 1.0 куб. см прибавлением воды. Теперь уже мы имеем два раствора соков, эквивалентные по содержанию фермента, по кислотности и одинаковые по разведению. Свертывая молоко этими растворами и получая свертывание в одно и то же время, мы должны признать пропорциональность свертывающего и растворяющего действия (табл. 4).
Параллелизм обеих функций сразу бросается в глаза (табл. 3). Но есть ли пропорциональность? Берем переваривающую силу соков, выраженную в миллиметрах белковой палочки, и, пользуясь правилом Шюца- Борисова, находим, что хлебный сок почти в 5 раз сильнее молочного сока. Обращаясь к свертывающему действию тех же соков, видим, что хлебный сок свертывает молоко в 6/2 раз скорее, чем молочный.
Таким образом параллелизм - налицо, но пропорциональности нет. Но на самом деле это не так: пропорциональность существует, но нужно выбрать такие условия, чтобы ее легко было обнаружить. Ведь мы имеем перед собою две противоположных реакции, идущие при разных условиях и измеряемые по разным правилам.Для растворяющего действия мы имеем правило (Шюца-Борисова), что количество фермента, или его концентрация, прямо пропорционально квадратам миллиметров переваренного белка; для свертывающего действия - правило, что сила сока обратно пропорциональна времени свертывания молока. Каждое правило имеет для своего действия пределы, за которыми оно теряет свой безусловный характе, и может быть принято лишь приблизительно. Вот один из таких пределов: если молоко не свернется в течение десяти минут, мы - за пределом. В этом случае свертывание наступит гораздо позже, чем оно должно бы быть по правилу.
Поэтому, для того чтобы обнаружить пропорциональность свертывающего и растворяющего действия сока, мы, определив количественное отношение ферментов в соках по растворяющему действию, брали затем для свертывания молока эквивалентные по содержанию фермента количества сравниваемых соков. Поясню примером. В предыдущей таблице молочный сок оказался почти в 5 раз слабее хлебного (правило Шюца - Борисова). Если я беру для свертывания молока 0.1 куб. см хлебного сока, то эквивалентное ему количество молочного сока будет в 5 раз больше, т. е. 0.5 куб. см. Но этого недостаточно: взяв в 5 раз больше молочного сока, я, конечно, взял больше и кислоты. Поэтому мы уравниваем кислотность прибавлением к 0.1 куб. см хлебного сока известного количества HCI, точно определенного титрованием. Теперь уже мы имеем два раствора соков, эквивалентные как по количеству содержащегося в них: фермента, так и по кислотности. Но и это еще не все. Представьте, что два соответственных эквивалентных раствора с одинаковым содержанием фермента и с уравненной кислотностью будут разниться по количеству куб. сантиметров, например 1.0 и 0.5 куб. см. Приливая 1.0 куб. см раствора сока к 10.0 куб. см молока, мы больше разжижаем молоко, чем в другой пробирке, где к 10.0 куб. см молока прилито 0.5 куб. см сока. Чтобы уравнять все условия, мы должны уравнять и разведение. С этой целью мы в последнем случае доводим количество раствора сока с 0.5 куб. см до 1.0 куб. см прибавлением воды. Теперь уже мы имеем два раствора соков, эквивалентные по содержанию фермента, по кислотности и одинаковые по разведению. Свертывая молоко этими растворами и получая свертывание в одно и то же время, мы должны признать пропорциональность свертывающего и растворяющего действия (табл. 4).
 Из этой таблицы (4) видно, что пропорциональность несомненна, отсюда же видно и то, что для обнаружения пропорциональности нужно уравнять все условия свертывания молока сравниваемыми соками.
Какой результат опытов с желудочным соком заставил нас сделать проверку прежних опытов с другими пищеварительными соками, и мы убедились в том же самом. Получилась такая же пропорциональность растворяющего и свертывающего действия в соке пилорическом и в соке бруннеровых желез, которые приравнивались к эквивалентным количеством фундального желудочного сока и затем испытывались так, как я указал выше, на свертывание молока. Все это не оставляет никакого сомнения в полном тождестве противоположных функций белкового фермента.
Дальнейшей нашей задачей было показать, что утверждение о двойственности белковых ферментов основано на ошибке, и затем выяснить, в чем заключается эта ошибка.
Двойственность белковых ферментов доказывалась разными способами. Первое доказательство защитника этой двойственности, Гамарстена, касается желудочного сока и заключается в том, что разрушающийся постепенно в термостате желудочный сок через некоторое время совершенно утрачивает свертывающее действие на молоко, сохраняя, однако, растворяющее действие на белок. Я упомянул уже, что в наших опытах разрушавшийся в термостате желудочный сок обнаружил полный параллелизм падения обоих действий - растворяющего свертывающего. В чем же заключается ошибка Гамарстена? В том прежде всего, что он, стараясь доказать, что свертывание молока обязано ферменту, а не кислоте сока, усердно нейтрализовал и даже подщелачивал желудочный сок, не обращая внимания на то, что это подщелачивание гибельно действует на фермент. А до какой степени чувствителен фермент к реакции среды, свидетельствует следующий наш опыт. Если мы возьмем две порции одного и того же желудочного сока и одну порцию доведем до нейтральной реакции с помощью Natrium bicarbonicum (NaHCO3), а другую --- Natrium carbonicum (Na2CO3), то, несмотря на то, что обе порции будут одинаково нейтральны, порция с Natrium carbonicum как с солью более щелочной обнаружит более слабое ферментное действие, чем порция с Natrium bicarbonicum. Гамарстен в своем стремлении исключить действие кислоты не обратил на это должного внимания и пришел к ошибочному выводу. Второе обстоятельство, помешавшее ему заметить ошибку, заключалось в том, что правило для свертывающего действия фермента (обратно пропорционально времени свертывания) имеет силу только до известного предела, за которым время свертывания увеличивается гораздо скорее, чем уменьшается фермент. Гамарстен не оценил этого и, конечно, впал в ошибку.
B недавнее время из лаборатории Гофмейстера появилась работа Глэснера, в которой он дает новый способ разделения ферментов в желудочном соке. Но и этот способ по проверке оказался ошибочным. Методика Глэснера заключалась в прибавлении к раствору профермента двух солей: фосфорнокислого натра и уксуснокислого уранила. При этом в растворе происходит между солями реакция взаимного обмена и образуется осадок фосфорнокислого уранила, который, по мнению Глэснера, увлекает с собой один фермент, другой же фермент остается в растворе. И действительно, если испытывать раствор на переваривающую функцию, то мы ее не получим, но тем не менее никакого разделения тут не существует. Автор не обратил внимания на то, что при его способе в растворе должен образоваться уксуснокислый натрий, который значительной степени задерживает, как бы уничтожает растворяющее действие фермента. Что же касается молоко свертывающего действия, то при оценке его автор пользовался крайне грубым способом определения. Он брал литр молока, и если это молоко не свертывалась от сока в течение получаса, то он отрицал в соке свертывающее действие.
Как проверить эти заключения? Мы проделали опыты Глэснера. При этом мы прибавляли к соку фосфорнокислого натрия и уксуснокислого уранила в одном случае столько, чтобы получился лишь очень небольшой осадок, который не может увлечь фермента, а в другом случае - больше, так чтобы получился очень большой осадок, при образовании которого можно рассчитывать на увлечение им фермента. Испытывая затем соки на переваривание и свертывание, мы увидели полный параллелизм ослабления и переваривающей и свертывающей функции. Таким образом и здесь открывается как ошибочность методики, так и источник ошибки.
Следующий способ, которым думали разделить ферменты, принадлежит Гамарстену. Заключается он в том, что нейтрализованный желудочный сок взбалтывается с порошком углекислой магнезии. По Гамарстену, пепсин увлекается порошком магнезиальной соли, а химозин остается в растворе. И действительно, фильтрат сока, обработанного углекислой магнезией, молоко свертывает, но переваривающего действия не обнаруживает; следовательно, параллелизм как будто нарушен. На самом деле и этого нет. Стоит только разбавить фильтрат и соответственно подкислить, параллелизм восстанавливается. Таким образом мы имеем здесь дело не с исчезновением из фильтрата пепсина, а с замаскированием растворяющего действия сока вследствие неблагоприятных условий для его проявления.
Таким образом параллелизм растворяющего и свертывающего действий желудочного сока при такой методике восстановлен; этого мало: надо было показать и пропорциональность обоих действий. Эта задача долгое время не поддавалась разрешению; в то время как свертывающее действие фильтрата на молоко уменьшалось, по сравнению с контрольным - нейтральным соком, сравнительно немного (в 15-30 раз), растворяющее действие фильтрата падало в огромное количество раз (в 500 2000 3000 раз). Такой результат давал повод, пожалуй, сделать заключение о самостоятельности каждой функции, о существовании двух ферментов, и, надо сознаться, нам пришлось много побиться, чтобы уничтожить такую непропорциональность обеих функций и сблизить их. Решение этой задачи далось нам в руки, наконец, почти случайно.
Дело в том, что, обрабатывая сок углекислой магнезией, мы подщелачиваем его. Свертывающее действие фильтрата и идет в этой реакции. Для определения растворяющего действия мы должны были подкислить фильтрат, мы подкисляли и получали то во много раз слабейшее действие, о котором я сейчас упоминал. После долгих попыток усилить это действие мы убедились, что для этого надо подкислять не сразу, в два приема: сначала довести щелочной фильтрат до нейтральной реакции, оставить его в таком виде на некоторое время и затем уже подкислить. В этом случае переваривающее действие фильтрата значительно вырастает. Этот в высшей степени интересный факт иллюстрируется следующей таблицей (5).
Из табл. 5 видно, что во втором случае, где фильтрат был сперва нейтрализован и затем подкислен, растворяющая сила его ослабела гораздо меньше, чем свертывающая функция. Между тем при подкислении фильтрата сразу бывает наоборот.
Раз проведение фильтрата через нейтральную стадию до подкисления его значительно увеличивает растворяющее действие, естественно было посмотреть, как влияет продолжительность нейтрального состояния на переваривающую силу. С этой целью поставлен опыт (табл. 6): щелочной фильтрат нейтрализован и из него через разные промежутки времени брались порции и тотчас подкислялись.
Отсюда (табл. 6) ясно, что чем дольше стоял фильтрат в нейтральном виде, тем больше вырастала его переваривающая сила.
Из этой таблицы (4) видно, что пропорциональность несомненна, отсюда же видно и то, что для обнаружения пропорциональности нужно уравнять все условия свертывания молока сравниваемыми соками.
Какой результат опытов с желудочным соком заставил нас сделать проверку прежних опытов с другими пищеварительными соками, и мы убедились в том же самом. Получилась такая же пропорциональность растворяющего и свертывающего действия в соке пилорическом и в соке бруннеровых желез, которые приравнивались к эквивалентным количеством фундального желудочного сока и затем испытывались так, как я указал выше, на свертывание молока. Все это не оставляет никакого сомнения в полном тождестве противоположных функций белкового фермента.
Дальнейшей нашей задачей было показать, что утверждение о двойственности белковых ферментов основано на ошибке, и затем выяснить, в чем заключается эта ошибка.
Двойственность белковых ферментов доказывалась разными способами. Первое доказательство защитника этой двойственности, Гамарстена, касается желудочного сока и заключается в том, что разрушающийся постепенно в термостате желудочный сок через некоторое время совершенно утрачивает свертывающее действие на молоко, сохраняя, однако, растворяющее действие на белок. Я упомянул уже, что в наших опытах разрушавшийся в термостате желудочный сок обнаружил полный параллелизм падения обоих действий - растворяющего свертывающего. В чем же заключается ошибка Гамарстена? В том прежде всего, что он, стараясь доказать, что свертывание молока обязано ферменту, а не кислоте сока, усердно нейтрализовал и даже подщелачивал желудочный сок, не обращая внимания на то, что это подщелачивание гибельно действует на фермент. А до какой степени чувствителен фермент к реакции среды, свидетельствует следующий наш опыт. Если мы возьмем две порции одного и того же желудочного сока и одну порцию доведем до нейтральной реакции с помощью Natrium bicarbonicum (NaHCO3), а другую --- Natrium carbonicum (Na2CO3), то, несмотря на то, что обе порции будут одинаково нейтральны, порция с Natrium carbonicum как с солью более щелочной обнаружит более слабое ферментное действие, чем порция с Natrium bicarbonicum. Гамарстен в своем стремлении исключить действие кислоты не обратил на это должного внимания и пришел к ошибочному выводу. Второе обстоятельство, помешавшее ему заметить ошибку, заключалось в том, что правило для свертывающего действия фермента (обратно пропорционально времени свертывания) имеет силу только до известного предела, за которым время свертывания увеличивается гораздо скорее, чем уменьшается фермент. Гамарстен не оценил этого и, конечно, впал в ошибку.
B недавнее время из лаборатории Гофмейстера появилась работа Глэснера, в которой он дает новый способ разделения ферментов в желудочном соке. Но и этот способ по проверке оказался ошибочным. Методика Глэснера заключалась в прибавлении к раствору профермента двух солей: фосфорнокислого натра и уксуснокислого уранила. При этом в растворе происходит между солями реакция взаимного обмена и образуется осадок фосфорнокислого уранила, который, по мнению Глэснера, увлекает с собой один фермент, другой же фермент остается в растворе. И действительно, если испытывать раствор на переваривающую функцию, то мы ее не получим, но тем не менее никакого разделения тут не существует. Автор не обратил внимания на то, что при его способе в растворе должен образоваться уксуснокислый натрий, который значительной степени задерживает, как бы уничтожает растворяющее действие фермента. Что же касается молоко свертывающего действия, то при оценке его автор пользовался крайне грубым способом определения. Он брал литр молока, и если это молоко не свертывалась от сока в течение получаса, то он отрицал в соке свертывающее действие.
Как проверить эти заключения? Мы проделали опыты Глэснера. При этом мы прибавляли к соку фосфорнокислого натрия и уксуснокислого уранила в одном случае столько, чтобы получился лишь очень небольшой осадок, который не может увлечь фермента, а в другом случае - больше, так чтобы получился очень большой осадок, при образовании которого можно рассчитывать на увлечение им фермента. Испытывая затем соки на переваривание и свертывание, мы увидели полный параллелизм ослабления и переваривающей и свертывающей функции. Таким образом и здесь открывается как ошибочность методики, так и источник ошибки.
Следующий способ, которым думали разделить ферменты, принадлежит Гамарстену. Заключается он в том, что нейтрализованный желудочный сок взбалтывается с порошком углекислой магнезии. По Гамарстену, пепсин увлекается порошком магнезиальной соли, а химозин остается в растворе. И действительно, фильтрат сока, обработанного углекислой магнезией, молоко свертывает, но переваривающего действия не обнаруживает; следовательно, параллелизм как будто нарушен. На самом деле и этого нет. Стоит только разбавить фильтрат и соответственно подкислить, параллелизм восстанавливается. Таким образом мы имеем здесь дело не с исчезновением из фильтрата пепсина, а с замаскированием растворяющего действия сока вследствие неблагоприятных условий для его проявления.
Таким образом параллелизм растворяющего и свертывающего действий желудочного сока при такой методике восстановлен; этого мало: надо было показать и пропорциональность обоих действий. Эта задача долгое время не поддавалась разрешению; в то время как свертывающее действие фильтрата на молоко уменьшалось, по сравнению с контрольным - нейтральным соком, сравнительно немного (в 15-30 раз), растворяющее действие фильтрата падало в огромное количество раз (в 500 2000 3000 раз). Такой результат давал повод, пожалуй, сделать заключение о самостоятельности каждой функции, о существовании двух ферментов, и, надо сознаться, нам пришлось много побиться, чтобы уничтожить такую непропорциональность обеих функций и сблизить их. Решение этой задачи далось нам в руки, наконец, почти случайно.
Дело в том, что, обрабатывая сок углекислой магнезией, мы подщелачиваем его. Свертывающее действие фильтрата и идет в этой реакции. Для определения растворяющего действия мы должны были подкислить фильтрат, мы подкисляли и получали то во много раз слабейшее действие, о котором я сейчас упоминал. После долгих попыток усилить это действие мы убедились, что для этого надо подкислять не сразу, в два приема: сначала довести щелочной фильтрат до нейтральной реакции, оставить его в таком виде на некоторое время и затем уже подкислить. В этом случае переваривающее действие фильтрата значительно вырастает. Этот в высшей степени интересный факт иллюстрируется следующей таблицей (5).
Из табл. 5 видно, что во втором случае, где фильтрат был сперва нейтрализован и затем подкислен, растворяющая сила его ослабела гораздо меньше, чем свертывающая функция. Между тем при подкислении фильтрата сразу бывает наоборот.
Раз проведение фильтрата через нейтральную стадию до подкисления его значительно увеличивает растворяющее действие, естественно было посмотреть, как влияет продолжительность нейтрального состояния на переваривающую силу. С этой целью поставлен опыт (табл. 6): щелочной фильтрат нейтрализован и из него через разные промежутки времени брались порции и тотчас подкислялись.
Отсюда (табл. 6) ясно, что чем дольше стоял фильтрат в нейтральном виде, тем больше вырастала его переваривающая сила.
 Покончив с этим пунктом, мы должны перейти к следующему возражению. Нам говорят: какое может быть тождество между пепсином и химозином, когда растворение белка и свертывание молока идут по двум разным правилам? Какое может быть тождество, когда по растворяющему действию сока количество фермента прямо пропорционально квадрату миллиметров переваренной белковой палочки, а по свертывающему действию мерой служит обратная пропорциональность временам свертывания? Но против такого возражения можно выставить теоретическое рассуждение. Тождества правил можно требовать для одинаковых реакций. В данном же случае одинаковые правила вовсе не обязательны, так как самые реакции-то разные. Для хода реакции имеет громадное значение химическая среда. Мы видим уже, что известные химические манипуляции с соком реакцию растворения сводят к нулю, оставляя в силе реакцию свертывания. Далее, изменяя условия среды, нетрудно заставить каждую из этих двух реакций пойти по разным правилам. Возьмем реакцию свертывания. Вот ее обычный тип (табл. 7).
Покончив с этим пунктом, мы должны перейти к следующему возражению. Нам говорят: какое может быть тождество между пепсином и химозином, когда растворение белка и свертывание молока идут по двум разным правилам? Какое может быть тождество, когда по растворяющему действию сока количество фермента прямо пропорционально квадрату миллиметров переваренной белковой палочки, а по свертывающему действию мерой служит обратная пропорциональность временам свертывания? Но против такого возражения можно выставить теоретическое рассуждение. Тождества правил можно требовать для одинаковых реакций. В данном же случае одинаковые правила вовсе не обязательны, так как самые реакции-то разные. Для хода реакции имеет громадное значение химическая среда. Мы видим уже, что известные химические манипуляции с соком реакцию растворения сводят к нулю, оставляя в силе реакцию свертывания. Далее, изменяя условия среды, нетрудно заставить каждую из этих двух реакций пойти по разным правилам. Возьмем реакцию свертывания. Вот ее обычный тип (табл. 7).

 Здесь (табл. 7) реакция свертывания идет по правилу обратной пропорциональности временам свертывания: где сычуга взято втрое больше, там и время свертывания молока втрое короче. Но это правило легко нарушить, что видео из следующей таблицы (8).
Взяв кислый сок, мы получим уже новое правило (табл. 8). Если 0.1 куб. см сока свернул молоко в 11 минут, то по предыдущему правилу 0.2 этого сока, содержащее вдвое больше фермента, должно бы свернуть вдвое скорее, т. е. в 5.5 минуты. А на самом деле свернуло гораздо скорей - в 3 минуты. Получилось новое правило - обратное отношение квадратным корням из времеи свертывания. Чем же объясняется это? Тем, что, приливая к молоку вдвое больше сока, приливал не только вдвое большее количество фермента, но и вдвое больше кислоты, которая и ускоряет ход реакции. Но я могу и замедлить реакцию.
Здесь (табл. 7) реакция свертывания идет по правилу обратной пропорциональности временам свертывания: где сычуга взято втрое больше, там и время свертывания молока втрое короче. Но это правило легко нарушить, что видео из следующей таблицы (8).
Взяв кислый сок, мы получим уже новое правило (табл. 8). Если 0.1 куб. см сока свернул молоко в 11 минут, то по предыдущему правилу 0.2 этого сока, содержащее вдвое больше фермента, должно бы свернуть вдвое скорее, т. е. в 5.5 минуты. А на самом деле свернуло гораздо скорей - в 3 минуты. Получилось новое правило - обратное отношение квадратным корням из времеи свертывания. Чем же объясняется это? Тем, что, приливая к молоку вдвое больше сока, приливал не только вдвое большее количество фермента, но и вдвое больше кислоты, которая и ускоряет ход реакции. Но я могу и замедлить реакцию.
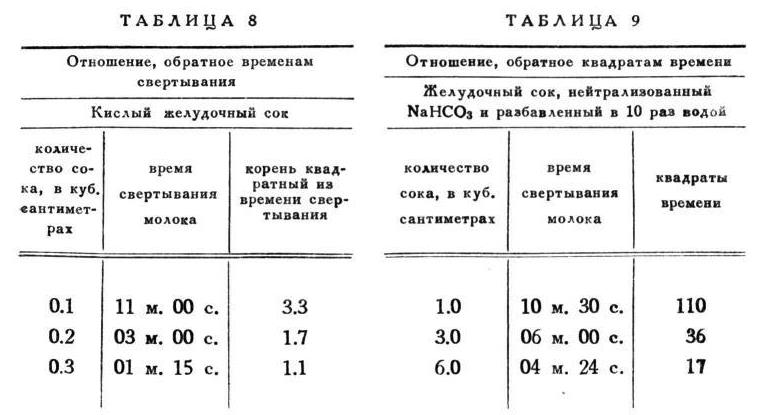 Здесь (табл. 9) я получаю отношение, обратное квадратам времени свертывания. Здесь берутся большие количества сока, и сока нейтрального. Чем больше я его прибавлю к молоку, тем больше я и разжижаю молоко, затрудняя его свертывание. Таким образом и создается третье правило для свертывающего действия.
Здесь (табл. 9) я получаю отношение, обратное квадратам времени свертывания. Здесь берутся большие количества сока, и сока нейтрального. Чем больше я его прибавлю к молоку, тем больше я и разжижаю молоко, затрудняя его свертывание. Таким образом и создается третье правило для свертывающего действия.
 То же самое можно показать и для другой реакции -- реакции растворения (табл. 10).
Из этой таблицы (10) видна прямая пропорциональность между количеством фермента и квадратами миллиметров переваренной белковой палочки - правило Шюца-Борисова.
Теперь разбавим желудочный сок кислотой, а водой (табл. 11).
То же самое можно показать и для другой реакции -- реакции растворения (табл. 10).
Из этой таблицы (10) видна прямая пропорциональность между количеством фермента и квадратами миллиметров переваренной белковой палочки - правило Шюца-Борисова.
Теперь разбавим желудочный сок кислотой, а водой (табл. 11).
 Из этих цифр (табл. 11) с очевидностью видно, что получилось уже не отношение квадратов, простое. Почему? Потому что мы изменили условия реакции, что на первый взгляд может показаться и несущественным. В первом случае, разбавляя сок соляной кислотой, мы постепенно изменяли отношение кислоты к ферменту, а во втором случае это отношение оставалось неизменным: разбавляя сок, например, в 8 раз водой, мы в 8 раз уменьшали концентрацию фермента и во столько же раз уменьшали и концентрацию кислоты. И в результате получили новое правило.
Таким образом, изменяя реакцию среды, мы можем каждую функцию фермента направить по разным правилам.
После всех этих опытов, которыми мы не только выяснили ошибочность заключений сторонников двойственности ферментов, но и указали также источники этих ошибок, не должно оставаться никакого сомнения в том, что две эти функции пищеварительных соков представляют собою проявление одного и того же фермента.
Н. П. К р а в к о в: На основании этих исследований нельзя ли также сделать предположение о единстве трех ферментов панкреатического сока?
И. П. П а в л о в: Нет, ибо в панкреатическом соке нет того параллелизма, на который указывал здесь. и крахмальный ферменты разрушаются быстро, а белковый остается.
Из этих цифр (табл. 11) с очевидностью видно, что получилось уже не отношение квадратов, простое. Почему? Потому что мы изменили условия реакции, что на первый взгляд может показаться и несущественным. В первом случае, разбавляя сок соляной кислотой, мы постепенно изменяли отношение кислоты к ферменту, а во втором случае это отношение оставалось неизменным: разбавляя сок, например, в 8 раз водой, мы в 8 раз уменьшали концентрацию фермента и во столько же раз уменьшали и концентрацию кислоты. И в результате получили новое правило.
Таким образом, изменяя реакцию среды, мы можем каждую функцию фермента направить по разным правилам.
После всех этих опытов, которыми мы не только выяснили ошибочность заключений сторонников двойственности ферментов, но и указали также источники этих ошибок, не должно оставаться никакого сомнения в том, что две эти функции пищеварительных соков представляют собою проявление одного и того же фермента.
Н. П. К р а в к о в: На основании этих исследований нельзя ли также сделать предположение о единстве трех ферментов панкреатического сока?
И. П. П а в л о в: Нет, ибо в панкреатическом соке нет того параллелизма, на который указывал здесь. и крахмальный ферменты разрушаются быстро, а белковый остается.
 Кислотность и разведение уравнены только в каждом горизонтальном ряду (табл. 1).
Из табл. 1 следует, что в различных сортах сока оба действия идут вполне параллельно. (Нельзя при этом не обратить внимания на курьезное обстоятельство, что химозина -- молочного фермента, - если стоять на общепринятой точке зрения, как раз всего меньше выделяется на субстрат его действия). Однако в той же таблице бросается в глаза резкая непропорциональность в действиях. При переходе от мясного сока к молочному молоко свертывающее действие падает гораздо значительнее, чем белок растворяющее. Оставляя объяснение этого обстоятельства до позднейших частей нашего изложения, мы приводим сейчас такую вариацию опыта, которая дает нам несомненную пропорциональность там, где имеется на первый взгляд только грубый параллелизм. Приводим таблицу этого опыта (табл. 2).
Кислотность и разведение уравнены только в каждом горизонтальном ряду (табл. 1).
Из табл. 1 следует, что в различных сортах сока оба действия идут вполне параллельно. (Нельзя при этом не обратить внимания на курьезное обстоятельство, что химозина -- молочного фермента, - если стоять на общепринятой точке зрения, как раз всего меньше выделяется на субстрат его действия). Однако в той же таблице бросается в глаза резкая непропорциональность в действиях. При переходе от мясного сока к молочному молоко свертывающее действие падает гораздо значительнее, чем белок растворяющее. Оставляя объяснение этого обстоятельства до позднейших частей нашего изложения, мы приводим сейчас такую вариацию опыта, которая дает нам несомненную пропорциональность там, где имеется на первый взгляд только грубый параллелизм. Приводим таблицу этого опыта (табл. 2).
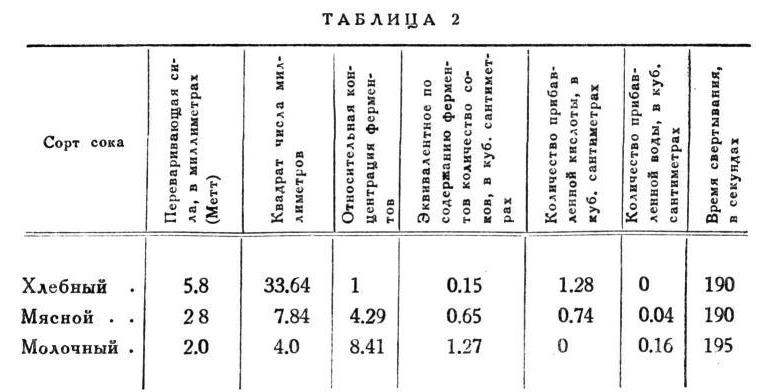 В первом столбце (табл. 2) обозначен сорт сока. Во втором столбце таблицы мы имеем переваривающую силу, в миллиметрах белковой палочки, по Мэтту, различных сортов, разбавленных в 10 раз раствором 0.2% соляной кислоты и стоявших в термостате одинаковое число часов (16 часов). В третьем столбце взяты квадраты чисел, изображающих переваривающую силу, т. e. относительные количества фермента. В четвертом столбце числа показывают, во сколько мясной и молочный соки слабее хлебного. В пятом приведены количества соков, эквивалентные по содержанию фермента. В шестом столбце приведены цифры прибавленной к порциям сока предшествующего ряда 0.2% соляной кислоты для того, чтобы во всех этих порциях кислоты было одинаковое количество. Количество прибавляемой кислоты рассчитывалось на основании титрования исходных разбавленных соков. В седьмом столбце имеются количества дистиллированной воды для того, чтобы во всех этих порциях уравнять массу жидкости. Таким образом мы имели три порции сока трех вышеозначенных сортов, в которых содержалось одинаковое количество фермента, переваривающего белки, одинаковое количество кислоты и одинаковое количество жидкости. Когда эти порции были прилиты, каждая к 10 куб. см молока, то получились цифры свертывания молока в секундах, приведенные в восьмом столбце таблицы. Из этого столбца ясно, что эквивалентные по содержанию белок растворяющего фермента количества хлебного, мясного и молочного соков, при прочих равных химических условиях, проявили совершенно одинаковое молоко свертывающее действие, т. е. оба наши действия в различных сортах сока содержатся строго пропорционально.
Затем испытывались часовые порции соков, излившихся на разные сорта еды. На прилагаемой таблице (3) все порции сока собраны от одной собаки и уравнены в отношении кислотности и разбавления. Для свертывания молока бралось по 0.3 куб. см сока (табл. 3). Отчетливый параллелизм выступает и здесь. Неболыние отклонения, встречающиеся то там, то сям, суть естественные следствия недостаточности методики, при небольших колебаниях в величине переваривания и при больших сроках времени свертывания. Такие небольшие отклонения то чаще, то реже встречались и в других подобных опытах. Случайность их происхождения, несомненно, доказывается тем, что они никогда не приурочивались ни к определенному сорту еды, ни к определенному часу отделительного периода.
В первом столбце (табл. 2) обозначен сорт сока. Во втором столбце таблицы мы имеем переваривающую силу, в миллиметрах белковой палочки, по Мэтту, различных сортов, разбавленных в 10 раз раствором 0.2% соляной кислоты и стоявших в термостате одинаковое число часов (16 часов). В третьем столбце взяты квадраты чисел, изображающих переваривающую силу, т. e. относительные количества фермента. В четвертом столбце числа показывают, во сколько мясной и молочный соки слабее хлебного. В пятом приведены количества соков, эквивалентные по содержанию фермента. В шестом столбце приведены цифры прибавленной к порциям сока предшествующего ряда 0.2% соляной кислоты для того, чтобы во всех этих порциях кислоты было одинаковое количество. Количество прибавляемой кислоты рассчитывалось на основании титрования исходных разбавленных соков. В седьмом столбце имеются количества дистиллированной воды для того, чтобы во всех этих порциях уравнять массу жидкости. Таким образом мы имели три порции сока трех вышеозначенных сортов, в которых содержалось одинаковое количество фермента, переваривающего белки, одинаковое количество кислоты и одинаковое количество жидкости. Когда эти порции были прилиты, каждая к 10 куб. см молока, то получились цифры свертывания молока в секундах, приведенные в восьмом столбце таблицы. Из этого столбца ясно, что эквивалентные по содержанию белок растворяющего фермента количества хлебного, мясного и молочного соков, при прочих равных химических условиях, проявили совершенно одинаковое молоко свертывающее действие, т. е. оба наши действия в различных сортах сока содержатся строго пропорционально.
Затем испытывались часовые порции соков, излившихся на разные сорта еды. На прилагаемой таблице (3) все порции сока собраны от одной собаки и уравнены в отношении кислотности и разбавления. Для свертывания молока бралось по 0.3 куб. см сока (табл. 3). Отчетливый параллелизм выступает и здесь. Неболыние отклонения, встречающиеся то там, то сям, суть естественные следствия недостаточности методики, при небольших колебаниях в величине переваривания и при больших сроках времени свертывания. Такие небольшие отклонения то чаще, то реже встречались и в других подобных опытах. Случайность их происхождения, несомненно, доказывается тем, что они никогда не приурочивались ни к определенному сорту еды, ни к определенному часу отделительного периода.
 Мы заключаем, что при различных физиологических условиях работы пепсиновых желез, как бы ни колебались абсолютные количества фермента в соке, оба действия его всегда идут параллельно.
Мы переходим к различным изменениям сока вне организма.
Кислый сок, полученный методом так называемого мнимого кормления собак, ставился в термостат, где он подвергался медленному разрушению. Из него каждый день или через день-два брались отдельные порции для исследования. Для контроля белковых палочек употреблялась особая порция того же или другого сока, оставшаяся стоять при комнатной температуре, как это делается в лаборатории постоянно для исключения каких-либо случайностей. Из нее также брался сок для свертывания молока. Гак как кислотность сока от стояния термостате или при комнатной температуре не изменяется сколько-нибудь заметно, то различные порции наших соков могли быть прямо сравниваемы как в отношении переваривающей силы, так и в отношении свертывающего действия. Свертывание посредством термостатного сока начиналось с небольших количеств сока. Когда время свертывания значительно удлинялось, переходили на большие количества. Когда белковые палочки перестали перевариваться термостатным соком за 10 часов, переходили к фибрину (табл. 4).
Совершенно параллельные отношения обоих действий сока, при долговременном стоянии его вне организма, бьют в глаза. В то время как в термостатном соке оба действия совершенно параллельно в течение 41-го дня постепенно приближаются к нулю, в соке, стоявшем при комнатной температуре, эти действия, в течение того же времени, остаются совершенно нетронутыми. Этот опыт совершенно с тем же результатом был проделан еще раз. Точно такой же опыт был произведен еще с новою порцией сока, с тою только разницей, что сок, употребляемый для свертывания молока, всякий раз нейтрализовался углекислым барием. Результат опыта был совершенно одинаков с предшествующим.
Наконец в следующем подобном ряде определений желудочный сок, применявшийся для свертывания молока, нейтрализовался содой. В этом случае свертывающее действие саморазрушающегося в термостате сока исчезало раньше, сравнительно с растворяющим. Причина этого явления будет объяснена впоследствии.
Мы заключаем, что при различных физиологических условиях работы пепсиновых желез, как бы ни колебались абсолютные количества фермента в соке, оба действия его всегда идут параллельно.
Мы переходим к различным изменениям сока вне организма.
Кислый сок, полученный методом так называемого мнимого кормления собак, ставился в термостат, где он подвергался медленному разрушению. Из него каждый день или через день-два брались отдельные порции для исследования. Для контроля белковых палочек употреблялась особая порция того же или другого сока, оставшаяся стоять при комнатной температуре, как это делается в лаборатории постоянно для исключения каких-либо случайностей. Из нее также брался сок для свертывания молока. Гак как кислотность сока от стояния термостате или при комнатной температуре не изменяется сколько-нибудь заметно, то различные порции наших соков могли быть прямо сравниваемы как в отношении переваривающей силы, так и в отношении свертывающего действия. Свертывание посредством термостатного сока начиналось с небольших количеств сока. Когда время свертывания значительно удлинялось, переходили на большие количества. Когда белковые палочки перестали перевариваться термостатным соком за 10 часов, переходили к фибрину (табл. 4).
Совершенно параллельные отношения обоих действий сока, при долговременном стоянии его вне организма, бьют в глаза. В то время как в термостатном соке оба действия совершенно параллельно в течение 41-го дня постепенно приближаются к нулю, в соке, стоявшем при комнатной температуре, эти действия, в течение того же времени, остаются совершенно нетронутыми. Этот опыт совершенно с тем же результатом был проделан еще раз. Точно такой же опыт был произведен еще с новою порцией сока, с тою только разницей, что сок, употребляемый для свертывания молока, всякий раз нейтрализовался углекислым барием. Результат опыта был совершенно одинаков с предшествующим.
Наконец в следующем подобном ряде определений желудочный сок, применявшийся для свертывания молока, нейтрализовался содой. В этом случае свертывающее действие саморазрушающегося в термостате сока исчезало раньше, сравнительно с растворяющим. Причина этого явления будет объяснена впоследствии.
 Кислый желудочный сок, полученный методом мнимого кормления, подвергался в течение пяти минут действию более высоких температур, а затем исследовался как на растворяющее, так и на свертывающее его действие (табл. 5). 5).
Кислый желудочный сок, полученный методом мнимого кормления, подвергался в течение пяти минут действию более высоких температур, а затем исследовался как на растворяющее, так и на свертывающее его действие (табл. 5). 5).
 Параллельное отношение обоих действий отчетливо выступает и в этой таблице (5). До 52° падение обоих действий медленное, после 52° оно делается стремительным для обоих, а при 62 20 приближается к нулю.
Далее исследовалось влияние различных химических веществ на ферментное свойство сока. Есть много случаев, где преследуемый нами параллелизм при этом как будто резко нарушается. Если к желудочному соку прибавить разных солей, желчи, алкоголя, сахару и т. д., то переваривающая сила его резко падает, а свертывающее действие может остаться без малейшего изменения или даже усилиться. И лишь только последнее действие не упало, можно быть уверенным, что проявится и нормальная сила растворения, раз только сок будет разживин, т. е. будет понижено в нем процентное содержание вещества, тормозящего реакцию растворения (табл. 6).
Параллельное отношение обоих действий отчетливо выступает и в этой таблице (5). До 52° падение обоих действий медленное, после 52° оно делается стремительным для обоих, а при 62 20 приближается к нулю.
Далее исследовалось влияние различных химических веществ на ферментное свойство сока. Есть много случаев, где преследуемый нами параллелизм при этом как будто резко нарушается. Если к желудочному соку прибавить разных солей, желчи, алкоголя, сахару и т. д., то переваривающая сила его резко падает, а свертывающее действие может остаться без малейшего изменения или даже усилиться. И лишь только последнее действие не упало, можно быть уверенным, что проявится и нормальная сила растворения, раз только сок будет разживин, т. е. будет понижено в нем процентное содержание вещества, тормозящего реакцию растворения (табл. 6).
 В приведенной таблице (6), как видно, употреблено 4 различных нормальных сока и только в каждом ряде с одним и тем же соком разбавление производилось раствором кислоты одной и той же кислотности, а свертывание молока одним и тем же количеством сока, так что все ряды не могут быть сравниваемы между собою в отношении переваривающего и свертывающего действий.
Там же (исключая специальные случаи относительно свертывания и нами еще не исследованные), где падает свертывающее действие, обыкновенно падает и растворяющее, например при действии щелочи и щелочных солей. В таком случае, очевидно, идет дело о разрушении фермента или переводе его в недеятельное состояние. При этом также обращает на себя внимание параллелизм обоих действий. Как известно, ферменты желудочного сока очень чувствительны к щелочам. Даже нейтрализация, и самая осторожная, NaHCO3 и Na2СОз, хотя бы до совершенно одинаковой степени нейтральности, уже дает разницу в ферментном действии сока; причем при нейтрализации Na2CO3 обыкновенно получается меньшее ферментное действие. И это небольшое уменьшение одинаково обнаруживается как на растворяющем, так и на свертывающем действиях (табл. 7).
В приведенной таблице (6), как видно, употреблено 4 различных нормальных сока и только в каждом ряде с одним и тем же соком разбавление производилось раствором кислоты одной и той же кислотности, а свертывание молока одним и тем же количеством сока, так что все ряды не могут быть сравниваемы между собою в отношении переваривающего и свертывающего действий.
Там же (исключая специальные случаи относительно свертывания и нами еще не исследованные), где падает свертывающее действие, обыкновенно падает и растворяющее, например при действии щелочи и щелочных солей. В таком случае, очевидно, идет дело о разрушении фермента или переводе его в недеятельное состояние. При этом также обращает на себя внимание параллелизм обоих действий. Как известно, ферменты желудочного сока очень чувствительны к щелочам. Даже нейтрализация, и самая осторожная, NaHCO3 и Na2СОз, хотя бы до совершенно одинаковой степени нейтральности, уже дает разницу в ферментном действии сока; причем при нейтрализации Na2CO3 обыкновенно получается меньшее ферментное действие. И это небольшое уменьшение одинаково обнаруживается как на растворяющем, так и на свертывающем действиях (табл. 7).

 Панкреатический сок с 10 % кишечного сока поставлен в термостат (t° to 38) в 12 часов 17 минут. В молоко для свертывания на каждые 10 куб. см прилито 0.5 куб. см соляной кислоты 0.5%, а затем в указанные в табл. 9 времена по 0.3 куб. см смеси соков.
Такая же смесь из тех же соков для переваривания белка приготовлена и поставлена в термостат в 12 часов 55 минут. B 1 час 10 минут берется первая порция сока для переваривания палочки с сывороточным белком, по Метту, в течение получаса в водяном термостате. Каждые полчаса берутся дальнейшие порции и таким же образом испытываются на переваривание. В табл. 10 приведены числа переваривания и квадраты их как относительное содержание фермента. знакомились при белок свертывающей функции панкреатического сока. Во-вторых, измеряя и сопоставляя оба действия (6 последних чисел табл. 9 и второй ряд чисел табл. 10) разрушающегося панкреатического сока, мы убеждаемся, что на этот раз существует прямо не только параллелизм в отношении обоих деиствий, но и точная пропорциональность, прочем количество фермента мы вычисляем при молоко свертывающей функции прямо по числам времени, а при белок растворяющей по квадратам чисел миллиметров переваренного сывороточного белка.
Панкреатический сок с 10 % кишечного сока поставлен в термостат (t° to 38) в 12 часов 17 минут. В молоко для свертывания на каждые 10 куб. см прилито 0.5 куб. см соляной кислоты 0.5%, а затем в указанные в табл. 9 времена по 0.3 куб. см смеси соков.
Такая же смесь из тех же соков для переваривания белка приготовлена и поставлена в термостат в 12 часов 55 минут. B 1 час 10 минут берется первая порция сока для переваривания палочки с сывороточным белком, по Метту, в течение получаса в водяном термостате. Каждые полчаса берутся дальнейшие порции и таким же образом испытываются на переваривание. В табл. 10 приведены числа переваривания и квадраты их как относительное содержание фермента. знакомились при белок свертывающей функции панкреатического сока. Во-вторых, измеряя и сопоставляя оба действия (6 последних чисел табл. 9 и второй ряд чисел табл. 10) разрушающегося панкреатического сока, мы убеждаемся, что на этот раз существует прямо не только параллелизм в отношении обоих деиствий, но и точная пропорциональность, прочем количество фермента мы вычисляем при молоко свертывающей функции прямо по числам времени, а при белок растворяющей по квадратам чисел миллиметров переваренного сывороточного белка.


 Наконец абсолютная величина молоко свертываюшего действия пилорического и бруннеровского соков вполне отвечает белок растворяющему их действию. В то время как белок растворяющее действие желудочного и поджелудочного соков, в их наиболее концентрированных сортах, приблизительно одинаково велико и вообще значительно, и молоко свертывающее действие их также большое и приблизительно равное. Белок растворяющее действие пилорического и бруннеровского соков резко меньше, равняется или даже меньше действия самых слабых сортов желудочного и поджелудочного соков. В совершенном соответствии с этим и молоко свертывающее действие первых соков также относительно слабое. Привожу таблицу (12) одного из относящихся сюда опытов.
Наконец абсолютная величина молоко свертываюшего действия пилорического и бруннеровского соков вполне отвечает белок растворяющему их действию. В то время как белок растворяющее действие желудочного и поджелудочного соков, в их наиболее концентрированных сортах, приблизительно одинаково велико и вообще значительно, и молоко свертывающее действие их также большое и приблизительно равное. Белок растворяющее действие пилорического и бруннеровского соков резко меньше, равняется или даже меньше действия самых слабых сортов желудочного и поджелудочного соков. В совершенном соответствии с этим и молоко свертывающее действие первых соков также относительно слабое. Привожу таблицу (12) одного из относящихся сюда опытов.
 В первом столбце (табл. 12) мы имеем обозначение сортов сока: из пепсиновых желез изолированного желудочка, разбавленный в 4 раза водой, затем сок из изолированной привратниковой части желудка, далее сок из изолированного отдела двенадцатиперстной кишки с бруннеровыми железами и, наконец, такой же сок другой собаки. Последние три сока разбавлены в 3 раза 0.5% раствором соляной кислоты. Разбавление всех соков сделано с таким расчетом, чтобы все они имели кислотность около 0.1%. 0/ Во втором столбце приведена переваривающая сила в миллиметрах белковой палочки. В третьем столбце имеются квадраты чисел, выражающих переваривающую силу. B четвертом - стоят цифры, обозначающие, во сколько раз больше содержится фермента в каждом соке сравнительно со слабейшим, который принят за единицy. Пятый столбец содержит эквивалентные по содержанию фермента количества разных соков. В шестом - количества 0.5% соляной кислоты, прибавленной к порциям соков предшествующего столбца, чтобы все эти порции содержали одинаковое количество кислоты. Расчет делался на основании предшествующего точного титрования исходных разбавленных соков. В седьмом - приведено количество воды, прибавленной к предшествующим порциям, чтобы уравнять во всех них общую массу жидкости. Таким образом приготовленные порции разных соков приливались каждая к 10 куб. см молока. Они свернули молоко в число минут, которые приведены в столбце восьмом. Беря крайние числа из этого столбца, находим, что они разнятся друг от друга в полтора раза. Если бы все наши порции свернули молоко в одно и то же время, то это значило бы, понятно, что их белок свертывающее действие точно пропорционально молоко растворяющему. Набоюдавшееся расхождение чисел в полтора раза надо считать очень незначительным, почти не ослабляющим вывода о пропорциональности действий. Надо принять, во-первых, во внимание, что сопоставлялись по молоко свертывающему действию соки, которые по содержанию белок растворяющего фермента в крайних случаях разнились в 25 раз. Во-вторых, растворяющее действие соков, стоящих более близко к нашей таблице, очень мало, а это обыкновенно ведет к ошибке при счете миллиметров, и именно в сторону уменьшения; на числах же свертывания совершенно ясно, что они уклоняются от равенства первым тем больше, чем меньше переваривающая сила порции. Мы не сомневаемся, что при более точном определении переваривающей силы этих соков мы получим совершенно такой же результат, который приведен выше при такой форме опыта над тремя сортами сока из пепсиновых желез. Во всяком же случае на основании этих опытов бесспорно и сейчас, что молоко свертывающее действие пилорического и бруннеровского соков отвечает вполне их белок растворяющему действию, т. е. молоко свертывающее действие их приблизительно в столько же раз меньше соответствующего действия сока из пепсиновых желез, во сколько белок растворяющее действие меньше соответствующей величины сока из пепсиновых желез.
Итак, мы видим, что в четырех пищеварительных жидкостях, обнаруживающих белок растворяющее действие, имеется всегда и молоко свертывающее действие и притом соответствующего первому действию размера. В каждом из них оба действия идут всегда параллельно при всевозможных физиологических условиях работы желез. Эти действия, что касается их скрытого или проявленного состояния, относятся совершенно одинаково, находясь в одинаковой фазе и проявляясь всегда одним и тем же агентом и приблизительно с одной и той же быстротой. При сохранении соков и разрушении их под влиянием температур - низких, как и более высоких, а также при разрушении соков некоторыми агентами оба действия все время относятся совершенно параллельно. Более того, там, где удалось произвести более точные исследования, обнаруживалась и точная пропорциональность.
На основании всего вышеприведенного мы получили право допускать, что оба действия зависят от одного и того же химического агента, суть различные реакции одного и того же фермента.
Но как же отнестись к тем фактам, которыми до настоящего времени устанавливалась обособленность ферментов для двух исследуемых нами физиолого-химических действий?
Очевидно, ввиду нового материала, эти факты надо было вновь проконтролировать и обсудить, тем более, что без труда можно было усмотреть несколько источников ошибок в старых заключениях. Хотя дело касается и более или менее известных фактов, но эти факты не были достаточно оценены при сравнительной методике определения молоко свертывающего и белок растворяющего действия.
Эти факты следующие. Обыкновенно принимается, что количества фермента, свертывающего молоко, относятся обратно пропорционально, как времена свертывания; но нетрудно убедиться, как это и заявляется некоторыми авторами, что для применения этого правила наступает предел в случае слабых растворов фермента. Это особенно скоро обнаруживается при натуральных растворах фермента, т. е. в различных соках.
Привожу таблицу (13) соответствующих опытов с нейтрализованными NaHCO3 хлебным и молочным желудочными соками.
В первом столбце (табл. 12) мы имеем обозначение сортов сока: из пепсиновых желез изолированного желудочка, разбавленный в 4 раза водой, затем сок из изолированной привратниковой части желудка, далее сок из изолированного отдела двенадцатиперстной кишки с бруннеровыми железами и, наконец, такой же сок другой собаки. Последние три сока разбавлены в 3 раза 0.5% раствором соляной кислоты. Разбавление всех соков сделано с таким расчетом, чтобы все они имели кислотность около 0.1%. 0/ Во втором столбце приведена переваривающая сила в миллиметрах белковой палочки. В третьем столбце имеются квадраты чисел, выражающих переваривающую силу. B четвертом - стоят цифры, обозначающие, во сколько раз больше содержится фермента в каждом соке сравнительно со слабейшим, который принят за единицy. Пятый столбец содержит эквивалентные по содержанию фермента количества разных соков. В шестом - количества 0.5% соляной кислоты, прибавленной к порциям соков предшествующего столбца, чтобы все эти порции содержали одинаковое количество кислоты. Расчет делался на основании предшествующего точного титрования исходных разбавленных соков. В седьмом - приведено количество воды, прибавленной к предшествующим порциям, чтобы уравнять во всех них общую массу жидкости. Таким образом приготовленные порции разных соков приливались каждая к 10 куб. см молока. Они свернули молоко в число минут, которые приведены в столбце восьмом. Беря крайние числа из этого столбца, находим, что они разнятся друг от друга в полтора раза. Если бы все наши порции свернули молоко в одно и то же время, то это значило бы, понятно, что их белок свертывающее действие точно пропорционально молоко растворяющему. Набоюдавшееся расхождение чисел в полтора раза надо считать очень незначительным, почти не ослабляющим вывода о пропорциональности действий. Надо принять, во-первых, во внимание, что сопоставлялись по молоко свертывающему действию соки, которые по содержанию белок растворяющего фермента в крайних случаях разнились в 25 раз. Во-вторых, растворяющее действие соков, стоящих более близко к нашей таблице, очень мало, а это обыкновенно ведет к ошибке при счете миллиметров, и именно в сторону уменьшения; на числах же свертывания совершенно ясно, что они уклоняются от равенства первым тем больше, чем меньше переваривающая сила порции. Мы не сомневаемся, что при более точном определении переваривающей силы этих соков мы получим совершенно такой же результат, который приведен выше при такой форме опыта над тремя сортами сока из пепсиновых желез. Во всяком же случае на основании этих опытов бесспорно и сейчас, что молоко свертывающее действие пилорического и бруннеровского соков отвечает вполне их белок растворяющему действию, т. е. молоко свертывающее действие их приблизительно в столько же раз меньше соответствующего действия сока из пепсиновых желез, во сколько белок растворяющее действие меньше соответствующей величины сока из пепсиновых желез.
Итак, мы видим, что в четырех пищеварительных жидкостях, обнаруживающих белок растворяющее действие, имеется всегда и молоко свертывающее действие и притом соответствующего первому действию размера. В каждом из них оба действия идут всегда параллельно при всевозможных физиологических условиях работы желез. Эти действия, что касается их скрытого или проявленного состояния, относятся совершенно одинаково, находясь в одинаковой фазе и проявляясь всегда одним и тем же агентом и приблизительно с одной и той же быстротой. При сохранении соков и разрушении их под влиянием температур - низких, как и более высоких, а также при разрушении соков некоторыми агентами оба действия все время относятся совершенно параллельно. Более того, там, где удалось произвести более точные исследования, обнаруживалась и точная пропорциональность.
На основании всего вышеприведенного мы получили право допускать, что оба действия зависят от одного и того же химического агента, суть различные реакции одного и того же фермента.
Но как же отнестись к тем фактам, которыми до настоящего времени устанавливалась обособленность ферментов для двух исследуемых нами физиолого-химических действий?
Очевидно, ввиду нового материала, эти факты надо было вновь проконтролировать и обсудить, тем более, что без труда можно было усмотреть несколько источников ошибок в старых заключениях. Хотя дело касается и более или менее известных фактов, но эти факты не были достаточно оценены при сравнительной методике определения молоко свертывающего и белок растворяющего действия.
Эти факты следующие. Обыкновенно принимается, что количества фермента, свертывающего молоко, относятся обратно пропорционально, как времена свертывания; но нетрудно убедиться, как это и заявляется некоторыми авторами, что для применения этого правила наступает предел в случае слабых растворов фермента. Это особенно скоро обнаруживается при натуральных растворах фермента, т. е. в различных соках.
Привожу таблицу (13) соответствующих опытов с нейтрализованными NaHCO3 хлебным и молочным желудочными соками.
 Очевидно, к влиянию предела надо отнести вышеотмеченное резкое падение свертывающего действия в молочном соке сравнительно с другими сортами.
То же самое получается при панкреатическом соке и тем более при пилорическом и бруннеровском соках, представляющих самые слабые растворы фермента. Таким образом очень легко потерять из глаз фермент, на самом деле несомненно существующий в среде.
Второе обстоятельство, игравшие роль в рассматриваемом нами вопросе, - это разрушающее действие щелочей (о своеобразности этого действия речь еще впереди), даже соды, действие давно хорошо известное. Как показано выше, даже тщательное нейтрализование, ни разу не переходящее за пределы нейтральной реакции Na2CO3, постоянно связано с разрушением фермента; и только при такой же нейтрализации NaHCO3 фермент может быть полностью сохранен. Так как это различие большею частью не принималось во внимание, причем не только не преследовали идеальной нейтрализации, аопускали небольшой перевес щелочной реакции, стараясь вполне отграничить ферментное свертывание от кислотного, то неизбежно должно было существовать значительное разрушение фермента.
Оба приведенные обстоятельства, соединяясь вместе, легко подделывали как бы исчезание того фермента, к которому относили молоко свертывающее действие. Это, очевидно, было причиной того, что у Гамарстена при стоянии искусственного желудочного сока в термостате растворяющее действие оставалось, а свертывающее исчезало, т. е. как бы происходило разделение ферментов. То же видели и мы при этих условиях в натуральном соке, когда мы испытывали его на свертывание после нейтрализации содой.
Ясно, что для исследования обоих действий были взяты методы совершенно различной чувствительности, нисколько между собой ни сопоставленные, ни соразмеренные. Для растворяющего действия применялся большей частью чувствительнейший реагент - фибрин, реагент без какого-либо предела. Для свертывающего действия пользовались нейтрализованным соком, при котором очень быстро с уменьшением количества наступает предел действия, тем более, что авторы брали совершенно произвольно предельное, вообще короткое, время свертывания, очевидно, боясь самопроизвольного подкисления молока. Простой пример бесспорно доказывает это: возьмите крепкий хлебный сок, разбавьте его в несколько сот, даже тысяч раз и вы все же получите довольно быстрое растворение им фибрина - в час, полтора, два. И тот же хлебный сок, нейтрализованный NaHCO3 и разбавленный только в 10-20 раз, даст вам свертывание только после многих часов, а дальше разбавленный не свернет молока и совсем. А между тем очевидно: фермент, свертывающий молоко, никуда исчезнуть из него не мог, не разрушился, и присутствие его там легко можно доказать, сделав благоприятными условия реакции свертывания. Из сказанного ясно, что при правильной методике определения свертывающего действия фермента обязательно повышать чувствительность реакции свертывания, прибавляя или к испытуемым жидкостям, или к молоку вещества, ускоряющие свертывание, как: кислоты, соли кальция, бария и т. д. Конечно, сравниваемые порции испытуемых соков должны быть тщательно уравниваемы в отношении повышающих чувствительность реактивов, самостоятельное, свертывающее действие этих агентов, если таковое существует, должно быть всегда контролируемо. Никакой запутанности из применения такой методики не произойдет, потому что благоприятствующие свертыванию условия действуют, как мы увидим ниже, по определенным правилам. Когда ферментное свертывающее действие в данной среде ослабевает, то усилить это действие только увеличиванием количества жидкости, приливаемой к молоку, удается только до известного предела, потому что рядом с увеличиванием количества фермента в молоке начинает давать себя знать разжижение молока, конечно, резко понижающее способность его свертываться. Когда желудочный сок, стоя в термостате, постепенно разрушается и, нейтрализованный содой, все медленнее свертывает молоко, то сначала вы помогаете делу, увеличивая количество прибавляемого сока к молоку. Но затем, когда вы достигаете трех, четырех куб. см на 10 куб. см молока, этот прием начинает вам отказывать, а в это же время употребление кислого сока в количестве 0.5--1.0 куб. см дает вам свертывание еще в течение нескольких минут или десятков минут. Что в этом случае кислота вас не обманывает, что вы дейстительно имеете перед собой еще ферментное действие, только без кислоты не обнаруживающееся, - это доказывает простейший контрольный опыт. Стоит вам слабый сок нагреть до 70°, to и 1 куб. см его свернет молоко только после десяти часов стояния в термостате, совершенно так, как это сделал бы 1 куб. см 0.5% соляной кислоты.
Третье обстоятельство, недостаточно оцениваемое при методике сравнения молоко свертывающего и белок растворяющего действия различных ферментных препаратов, - это указанное выше задерживающее действие солей и многих других веществ на реакцию растворения белка, отнюдь не простирающееся на молоко свертывающее действие.
Очевидно, на этом обстоятельстве основан главным образом фабричный прием изготовления препаратов сычужного фермента. Одному из нас (И. Павлову) живо припоминается следующее: когда впервые явилась мысль о тождестве химозина и пепсина, профессор Ненцкий возразил против этой мысли фактом, передавая имевшийся у него препарат сычуга из Рижскойфабрики и предлагая показать ему в нем пепсинное действие. Жидкости в высшей степени энергично свертывала молоко, но, как мы ее ни подкисляли и ни разжижали (от 2 до 10 раз), фибрин в ней не только не переваривался, но даже уплотнялся.
В то время так и не удалось разрушить это возражение. Только позже, уже после смерти дорогого товарища, оказалось, что задача в данном случае была совсем легкая. Стоило разбавить этот препарат в 100 раз соляной кислотой 0.2%, и он переварил даже яичный вареный белок -- 1.3 мм за 10 часов (Мэтт). Рассчитывая переваривающую силу самого фабричного препарата по правилу Шюца и Борисова, ее надо было определить в 13 мм, что вполне отвечало бы огромное свертывающей силе этого препарата. Что здесь все дело заключается в прибавлении к экстракту сычужного желудка какого-то задерживающего вещества, доказывается следующею вариациею опыта (табл. 14).
То же самое мы получили и на многих других испытанных нами продажных препаратах сычужного фермента, в которых слабое или совсем отсутствующее растворяющее белки действие достигало размера, соответствующего молоко свертывающему действию их, раз только применялся прием надлежащего их подкисления и разжижения.
Надо думать, что, вследствие недостаточной оценки первого и третьего из перечисляемых обстоятельств, и совсем недавно предложенный метод, долженствующий доказать обособленность белок растворяющего и молоко свертывающего проферментов, метод Глэснера [58] отнюдь не достигает цели. Мы многократно применяли этот метод, строжайше следуя указаниям автора, и, однако, ни разу не достигли ни малейшего действительного разделения ферментов. Мы варьировали количество употреблявшихся для образования осадка уксуснокислого уранила и фоссернокислого натра, как только возможно, и имели все степени захватывания фермента осадком. При большом осадке фильтрат от него не обнаруживал никакого действия, зато оба действия оказывались в извлечении из осадка. При небольшом осадке совершенно наоборот. Случаи средних осадков дали все переходы между этими крайностями. Прилагаю таблицу (15).
Очевидно, к влиянию предела надо отнести вышеотмеченное резкое падение свертывающего действия в молочном соке сравнительно с другими сортами.
То же самое получается при панкреатическом соке и тем более при пилорическом и бруннеровском соках, представляющих самые слабые растворы фермента. Таким образом очень легко потерять из глаз фермент, на самом деле несомненно существующий в среде.
Второе обстоятельство, игравшие роль в рассматриваемом нами вопросе, - это разрушающее действие щелочей (о своеобразности этого действия речь еще впереди), даже соды, действие давно хорошо известное. Как показано выше, даже тщательное нейтрализование, ни разу не переходящее за пределы нейтральной реакции Na2CO3, постоянно связано с разрушением фермента; и только при такой же нейтрализации NaHCO3 фермент может быть полностью сохранен. Так как это различие большею частью не принималось во внимание, причем не только не преследовали идеальной нейтрализации, аопускали небольшой перевес щелочной реакции, стараясь вполне отграничить ферментное свертывание от кислотного, то неизбежно должно было существовать значительное разрушение фермента.
Оба приведенные обстоятельства, соединяясь вместе, легко подделывали как бы исчезание того фермента, к которому относили молоко свертывающее действие. Это, очевидно, было причиной того, что у Гамарстена при стоянии искусственного желудочного сока в термостате растворяющее действие оставалось, а свертывающее исчезало, т. е. как бы происходило разделение ферментов. То же видели и мы при этих условиях в натуральном соке, когда мы испытывали его на свертывание после нейтрализации содой.
Ясно, что для исследования обоих действий были взяты методы совершенно различной чувствительности, нисколько между собой ни сопоставленные, ни соразмеренные. Для растворяющего действия применялся большей частью чувствительнейший реагент - фибрин, реагент без какого-либо предела. Для свертывающего действия пользовались нейтрализованным соком, при котором очень быстро с уменьшением количества наступает предел действия, тем более, что авторы брали совершенно произвольно предельное, вообще короткое, время свертывания, очевидно, боясь самопроизвольного подкисления молока. Простой пример бесспорно доказывает это: возьмите крепкий хлебный сок, разбавьте его в несколько сот, даже тысяч раз и вы все же получите довольно быстрое растворение им фибрина - в час, полтора, два. И тот же хлебный сок, нейтрализованный NaHCO3 и разбавленный только в 10-20 раз, даст вам свертывание только после многих часов, а дальше разбавленный не свернет молока и совсем. А между тем очевидно: фермент, свертывающий молоко, никуда исчезнуть из него не мог, не разрушился, и присутствие его там легко можно доказать, сделав благоприятными условия реакции свертывания. Из сказанного ясно, что при правильной методике определения свертывающего действия фермента обязательно повышать чувствительность реакции свертывания, прибавляя или к испытуемым жидкостям, или к молоку вещества, ускоряющие свертывание, как: кислоты, соли кальция, бария и т. д. Конечно, сравниваемые порции испытуемых соков должны быть тщательно уравниваемы в отношении повышающих чувствительность реактивов, самостоятельное, свертывающее действие этих агентов, если таковое существует, должно быть всегда контролируемо. Никакой запутанности из применения такой методики не произойдет, потому что благоприятствующие свертыванию условия действуют, как мы увидим ниже, по определенным правилам. Когда ферментное свертывающее действие в данной среде ослабевает, то усилить это действие только увеличиванием количества жидкости, приливаемой к молоку, удается только до известного предела, потому что рядом с увеличиванием количества фермента в молоке начинает давать себя знать разжижение молока, конечно, резко понижающее способность его свертываться. Когда желудочный сок, стоя в термостате, постепенно разрушается и, нейтрализованный содой, все медленнее свертывает молоко, то сначала вы помогаете делу, увеличивая количество прибавляемого сока к молоку. Но затем, когда вы достигаете трех, четырех куб. см на 10 куб. см молока, этот прием начинает вам отказывать, а в это же время употребление кислого сока в количестве 0.5--1.0 куб. см дает вам свертывание еще в течение нескольких минут или десятков минут. Что в этом случае кислота вас не обманывает, что вы дейстительно имеете перед собой еще ферментное действие, только без кислоты не обнаруживающееся, - это доказывает простейший контрольный опыт. Стоит вам слабый сок нагреть до 70°, to и 1 куб. см его свернет молоко только после десяти часов стояния в термостате, совершенно так, как это сделал бы 1 куб. см 0.5% соляной кислоты.
Третье обстоятельство, недостаточно оцениваемое при методике сравнения молоко свертывающего и белок растворяющего действия различных ферментных препаратов, - это указанное выше задерживающее действие солей и многих других веществ на реакцию растворения белка, отнюдь не простирающееся на молоко свертывающее действие.
Очевидно, на этом обстоятельстве основан главным образом фабричный прием изготовления препаратов сычужного фермента. Одному из нас (И. Павлову) живо припоминается следующее: когда впервые явилась мысль о тождестве химозина и пепсина, профессор Ненцкий возразил против этой мысли фактом, передавая имевшийся у него препарат сычуга из Рижскойфабрики и предлагая показать ему в нем пепсинное действие. Жидкости в высшей степени энергично свертывала молоко, но, как мы ее ни подкисляли и ни разжижали (от 2 до 10 раз), фибрин в ней не только не переваривался, но даже уплотнялся.
В то время так и не удалось разрушить это возражение. Только позже, уже после смерти дорогого товарища, оказалось, что задача в данном случае была совсем легкая. Стоило разбавить этот препарат в 100 раз соляной кислотой 0.2%, и он переварил даже яичный вареный белок -- 1.3 мм за 10 часов (Мэтт). Рассчитывая переваривающую силу самого фабричного препарата по правилу Шюца и Борисова, ее надо было определить в 13 мм, что вполне отвечало бы огромное свертывающей силе этого препарата. Что здесь все дело заключается в прибавлении к экстракту сычужного желудка какого-то задерживающего вещества, доказывается следующею вариациею опыта (табл. 14).
То же самое мы получили и на многих других испытанных нами продажных препаратах сычужного фермента, в которых слабое или совсем отсутствующее растворяющее белки действие достигало размера, соответствующего молоко свертывающему действию их, раз только применялся прием надлежащего их подкисления и разжижения.
Надо думать, что, вследствие недостаточной оценки первого и третьего из перечисляемых обстоятельств, и совсем недавно предложенный метод, долженствующий доказать обособленность белок растворяющего и молоко свертывающего проферментов, метод Глэснера [58] отнюдь не достигает цели. Мы многократно применяли этот метод, строжайше следуя указаниям автора, и, однако, ни разу не достигли ни малейшего действительного разделения ферментов. Мы варьировали количество употреблявшихся для образования осадка уксуснокислого уранила и фоссернокислого натра, как только возможно, и имели все степени захватывания фермента осадком. При большом осадке фильтрат от него не обнаруживал никакого действия, зато оба действия оказывались в извлечении из осадка. При небольшом осадке совершенно наоборот. Случаи средних осадков дали все переходы между этими крайностями. Прилагаю таблицу (15).
 По нашему мнению, фактически противоречие результатов наших и Глэснера имело свое основание в том, что он, во-первых, в фильтрате не считался с задерживающим действием на реакцию растворения образующегося уксуснокислого натра (см. табл. 6), во-вторых, для свертывающего действия был взят совершенно произвольный и очень ограниченный предел - свертывание литра молока в 30 минут. Представим себе такой случай: пусть с осадком захвачено 43-44 фермента, а в фильтрате осталось или 9/4 3/ его. Уксуснокислый натр почти совершенно мешает обнаружиться растворяющему действию фермента, так что проявляется только одно молоко свертывающее действие. Извлеченный из осадка фермент, благодаря разбавлению извлекающею жидкостью, освободился из-под влияния уксуснокислого натрия, т. е. снова стал растворять белок в значительной степени. Молоко же свертывающее действие экстракта, вследствие значительного разжижения, оказалось за пределами данной методики.
По нашему мнению, фактически противоречие результатов наших и Глэснера имело свое основание в том, что он, во-первых, в фильтрате не считался с задерживающим действием на реакцию растворения образующегося уксуснокислого натра (см. табл. 6), во-вторых, для свертывающего действия был взят совершенно произвольный и очень ограниченный предел - свертывание литра молока в 30 минут. Представим себе такой случай: пусть с осадком захвачено 43-44 фермента, а в фильтрате осталось или 9/4 3/ его. Уксуснокислый натр почти совершенно мешает обнаружиться растворяющему действию фермента, так что проявляется только одно молоко свертывающее действие. Извлеченный из осадка фермент, благодаря разбавлению извлекающею жидкостью, освободился из-под влияния уксуснокислого натрия, т. е. снова стал растворять белок в значительной степени. Молоко же свертывающее действие экстракта, вследствие значительного разжижения, оказалось за пределами данной методики.
 Точно таким же образом при точной проверке оказался недостигающим цели и второй метод Гамарстена. Он, как известно, начинается с многократного взбалтывания искусственного сока с порошком углекислого магния, при этом, по автору, пепсин должен был из жидкости увлекаться порошком, а химозин оставаться в жидкости, и затем следовала дальнейшая процедура очищения химозина. Мы проделали первую фазу методики с натуральным желудочным соком и видели то, что было перед глазами у Гамарстена. Фильтрат (3-й, 4-й и 5-й), хотя и меньше нормальной жидкости, но все же значительно быстро свертывал молоко, между тем как подкисленный до нормальной кислотности (0.5) без разжижения или только при малом разжижении совершенно не переваривал сырого фибрина даже за целые сутки. Однако стоило только разбавить фильтрат в 5-10 раз кислотой 0.1-0.2 для того, чтобы растворяющее действие фибрина обнаружилось совершенно отчетливо. И сколько раз мы ни взбалтывали наш предварительно нейтрализованный сок с углекислым магнием, мы не могли получить такого фильтрата, который при указанных условиях был бы лишен растворяющего действия на фибрин. Таким образом исчезнувший было параллелизм обоих действий для нас восстановился и мы было сочли себя удовлетворенными. Впоследствии, когда мы вместо параллелизма стали настаивать на пропорциональности, хотя бы приблизительной, этот пункт представил нам огромные затруднения и вызвал было даже в нас горькие сомнения относительно основного нашего положения, хотя это было в конце длиннейшего ряда фактов, нами полученных и до тех пор нам благоприятных. Когда мы попробовали более точно сопоставить молоко свертывающее и белок растворяющее действие фильтрата после взбалтывания с углекислым магнием, мы не могли не видеть огромной непропорциональности в обоих действиях. В то время как молоко свертывающее действие фильтрата указывало на уменьшение фермента в несколько раз, самое большее - в несколько десятков раз, по белок растворяющему действию фермента в жидкости оставалось в несколько сот и даже тысяч раз меньше. Мы обратили внимание на все мыслимые для нас обстоятельства. Мы определили и устранили задерживающее влияние на растворение того хлористого натрия, который образовался при нейтрализации сока. Мы определили и приняли во внимание благоприятствующее свертыванию молока действие того небольшого количества углекислого магния, которое растворялось в нашей жидкости при взбалтывании. Мы испробовали всякие степени кислотности, но факт в общем оставался прежним. Указанные меры изменяли результат только очень незначительно. Однако при этом постепенно накоплялись наблюдения, направлявшие мысль в надлежащий пункт. Титруя последовательные фильтраты, мы заметили, что их щелочность (вследствие небольшого растворения углекислого магния с щелочной реакцией) постепенно возрастает до известного предела, а с нею вместе понижалось белок растворяющее действие фильтрата. Это наводило на мысль: не играет ли в данном случае роль именно щелочность фильтрата? Поэтому прежде всего мы обратились к осадку углекислого магния после взбалтывания: действительно ли с ним увлечен из жидкости пепсин? Растворив этот осадок в соляной кислоте, разжижив его в надлежащей степени, чтобы устранить задерживающее действие хлористого магния, и подкислив соответственно, мы легко и точно на многих опытах убедились, что значительного захватывания пепсина осадком не происходило. Полученный раствор обнаруживал ничтожные следы растворяющего и молоко свертываютего действия, сравнительно с фильтратом. Стало действительно ясно, что механизм наблюдавшихся явлений в первой фазе гамарстеновского метода совершенно не тот, каким его представляли до сих пор. Это было давно известное разрушение фермента щелочью, разрушение или какое-то химическое изменение фермента под влиянием щелочи. Но если свойства фильтрата и были результатом действия щелочи, то все же, ввиду факта расхождения наших действий в фильтрате, приходилось думать так, что щелочь сильнее разрушает фермент, растворяющий белок, чем фермент, свертывающий молоко. Однако, принимая во внимание некоторые особенности, бросившиеся нам в глаза, когда мы занимались опытами с разрушающим действием щелочей на ферменты желудочного сока, мы продолжали стоять на своей точке зрения и старались добиться восстановления пропорциональности между обоими действиями, которая нам так упрямо не давалась. Наконец наше упорство было награждено: при, повидимому, совершенно ничего не значащей вариации опыта наш фильтрат проявил, наконец, растворяющее белки действие, нисколько не меньшее, а скорее большее, чем какое можно было ожидать на основании его молоко свертывающего действия. До сих пор мы Фильтрат, сделавшийся щелочным от взбалтывания с углекислым магнием, для переваривания сразу делали кислом до известной степени. Теперь же мы сперва его только нейтрализовали, таким оставляли стоять некоторое время и лишь затем подкислили до той же степени. И в этом-то фильтрате обнаружилось давно желанное значительное растворяющее действие (табл. 16).
Точно таким же образом при точной проверке оказался недостигающим цели и второй метод Гамарстена. Он, как известно, начинается с многократного взбалтывания искусственного сока с порошком углекислого магния, при этом, по автору, пепсин должен был из жидкости увлекаться порошком, а химозин оставаться в жидкости, и затем следовала дальнейшая процедура очищения химозина. Мы проделали первую фазу методики с натуральным желудочным соком и видели то, что было перед глазами у Гамарстена. Фильтрат (3-й, 4-й и 5-й), хотя и меньше нормальной жидкости, но все же значительно быстро свертывал молоко, между тем как подкисленный до нормальной кислотности (0.5) без разжижения или только при малом разжижении совершенно не переваривал сырого фибрина даже за целые сутки. Однако стоило только разбавить фильтрат в 5-10 раз кислотой 0.1-0.2 для того, чтобы растворяющее действие фибрина обнаружилось совершенно отчетливо. И сколько раз мы ни взбалтывали наш предварительно нейтрализованный сок с углекислым магнием, мы не могли получить такого фильтрата, который при указанных условиях был бы лишен растворяющего действия на фибрин. Таким образом исчезнувший было параллелизм обоих действий для нас восстановился и мы было сочли себя удовлетворенными. Впоследствии, когда мы вместо параллелизма стали настаивать на пропорциональности, хотя бы приблизительной, этот пункт представил нам огромные затруднения и вызвал было даже в нас горькие сомнения относительно основного нашего положения, хотя это было в конце длиннейшего ряда фактов, нами полученных и до тех пор нам благоприятных. Когда мы попробовали более точно сопоставить молоко свертывающее и белок растворяющее действие фильтрата после взбалтывания с углекислым магнием, мы не могли не видеть огромной непропорциональности в обоих действиях. В то время как молоко свертывающее действие фильтрата указывало на уменьшение фермента в несколько раз, самое большее - в несколько десятков раз, по белок растворяющему действию фермента в жидкости оставалось в несколько сот и даже тысяч раз меньше. Мы обратили внимание на все мыслимые для нас обстоятельства. Мы определили и устранили задерживающее влияние на растворение того хлористого натрия, который образовался при нейтрализации сока. Мы определили и приняли во внимание благоприятствующее свертыванию молока действие того небольшого количества углекислого магния, которое растворялось в нашей жидкости при взбалтывании. Мы испробовали всякие степени кислотности, но факт в общем оставался прежним. Указанные меры изменяли результат только очень незначительно. Однако при этом постепенно накоплялись наблюдения, направлявшие мысль в надлежащий пункт. Титруя последовательные фильтраты, мы заметили, что их щелочность (вследствие небольшого растворения углекислого магния с щелочной реакцией) постепенно возрастает до известного предела, а с нею вместе понижалось белок растворяющее действие фильтрата. Это наводило на мысль: не играет ли в данном случае роль именно щелочность фильтрата? Поэтому прежде всего мы обратились к осадку углекислого магния после взбалтывания: действительно ли с ним увлечен из жидкости пепсин? Растворив этот осадок в соляной кислоте, разжижив его в надлежащей степени, чтобы устранить задерживающее действие хлористого магния, и подкислив соответственно, мы легко и точно на многих опытах убедились, что значительного захватывания пепсина осадком не происходило. Полученный раствор обнаруживал ничтожные следы растворяющего и молоко свертываютего действия, сравнительно с фильтратом. Стало действительно ясно, что механизм наблюдавшихся явлений в первой фазе гамарстеновского метода совершенно не тот, каким его представляли до сих пор. Это было давно известное разрушение фермента щелочью, разрушение или какое-то химическое изменение фермента под влиянием щелочи. Но если свойства фильтрата и были результатом действия щелочи, то все же, ввиду факта расхождения наших действий в фильтрате, приходилось думать так, что щелочь сильнее разрушает фермент, растворяющий белок, чем фермент, свертывающий молоко. Однако, принимая во внимание некоторые особенности, бросившиеся нам в глаза, когда мы занимались опытами с разрушающим действием щелочей на ферменты желудочного сока, мы продолжали стоять на своей точке зрения и старались добиться восстановления пропорциональности между обоими действиями, которая нам так упрямо не давалась. Наконец наше упорство было награждено: при, повидимому, совершенно ничего не значащей вариации опыта наш фильтрат проявил, наконец, растворяющее белки действие, нисколько не меньшее, а скорее большее, чем какое можно было ожидать на основании его молоко свертывающего действия. До сих пор мы Фильтрат, сделавшийся щелочным от взбалтывания с углекислым магнием, для переваривания сразу делали кислом до известной степени. Теперь же мы сперва его только нейтрализовали, таким оставляли стоять некоторое время и лишь затем подкислили до той же степени. И в этом-то фильтрате обнаружилось давно желанное значительное растворяющее действие (табл. 16).
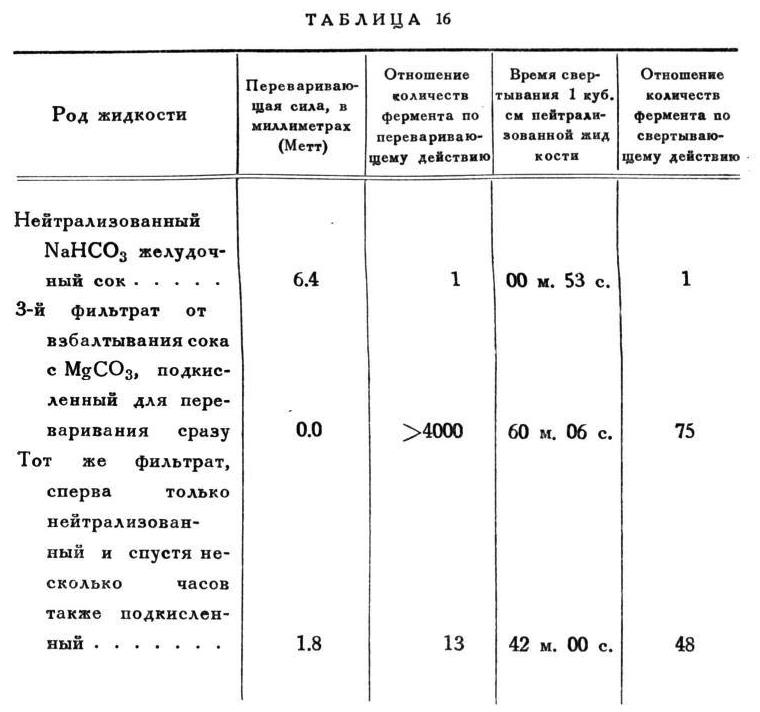 Таким образом (табл. 16) и в этом приеме Гамарстена не оказалось ни малейшего намека на разделение фермента. Такой результат переисследования первой фазы приема естественно исключал надобность изучения второй фазы.
Самый же факт, обнаружившийся неожиданно ири этом, конечно, был достоин всяческого внимания. Мы имели перед собою под влиянием щелочной реакции переход фермента из деятельного состояния в недеятельное, из явного в скрытое, причем теперь обратный переход - из недеятельного в деятельное состояние - требует некоторой продолжительности нейтральной реакции среды (каковая и делалась молоком). Специально относительно этого пункта нами было сейчас же поставлено несколько опытов, один из которых приводим здесь (табл. 17).
Таким образом (табл. 16) и в этом приеме Гамарстена не оказалось ни малейшего намека на разделение фермента. Такой результат переисследования первой фазы приема естественно исключал надобность изучения второй фазы.
Самый же факт, обнаружившийся неожиданно ири этом, конечно, был достоин всяческого внимания. Мы имели перед собою под влиянием щелочной реакции переход фермента из деятельного состояния в недеятельное, из явного в скрытое, причем теперь обратный переход - из недеятельного в деятельное состояние - требует некоторой продолжительности нейтральной реакции среды (каковая и делалась молоком). Специально относительно этого пункта нами было сейчас же поставлено несколько опытов, один из которых приводим здесь (табл. 17).
 При этих опытах нельзя не заметить, что при подкислении щелочной жидкости в один прием жидкость мутится и дает осадок, чего не бывает в случае, когда подкисление ведется в две фазы, c значительной остановкой (в несколько часов) на нейтральной реакции. Так как ясно, что факт стоит в ближайшем отношении к различным вопросам, касающимся фермента: действия щелочей, различного состояния фермента и т. д., то его изучение составило предмет дальнейшего и сейчас ведущегося в лаборатории исследования.
Весь изложенный материал, как нам кажется, отнимает всякую почву у ходячего мнения, что белки растворяющее и молоко свертывающее ферментные действия принадлежат особым ферментам. Факт тот, что эти действия никогда прочно не разделимы ни в организме, ни вне его ни при каких условиях. Есть, понятно, еще некоторые частные указания на прочную раздельность действий, но, не проверив их еще до сих пор, мы считали себя, однако, вправе не придавать им особенной силы ввиду обнаруженных нами недостатков современной методики сравнения этих действий.
Но мы давно уже остановились на общем вопросе: не противоречит ли нашему выводу, что оба исследуемые действия принадлежат одному и тому жерменту, то обстоятельство, что эти действия обнаруживаются по двум различным правилам зависимости от количества фермента? Это же представлялось существенным возражением против нашего взгляда и Гамарстену, что мы прочли уже во время писания этой статьи в последнем издании его учебника физиологической химии.
Наши обе реакции - растворение белка и свертывание молока - во всяком случае разные реакции, а если это так, то нет никакой необходимости в тождестве правил количественной зависимости времени реакции от концентрации ферментов. Скорость реакции определяется условиями реакции, которые то увеличивают, то уменьшают скорость. Соответственно характеру условий должны изменяться и правила зависимости скорости реакции от количества фермента в данном случае. С целью получить фактический материал, подтверждающий это, мы поставили несколько рядов опытов, где, меняя условия реакций, достигли соответствующего изменения правил. Нейтральный раствор молоко свертывающего фермента, раз он находится в жидкости в известной достаточной концентрации, свертывает молоко по правилу обратной пропорциональности между количествами фермента и временами свертывания. Хотя это давно известно, но мы позволяем себе, ради наглядного сравнения с последующим, привести таблицу (18), относящуюся к этому опыту.
Если мы берем кислый раствор того же фермента и прибавляем различное количество его к молоку, то теперь мы изменяем в молоке в совершенно той же пропорции количество как фермента, так и кислоты, т. е. если мы, например, увеличим вдвое количество фермента, то вместе с тем увеличиваем вдвое и количество кислоты в нем.
При этих опытах нельзя не заметить, что при подкислении щелочной жидкости в один прием жидкость мутится и дает осадок, чего не бывает в случае, когда подкисление ведется в две фазы, c значительной остановкой (в несколько часов) на нейтральной реакции. Так как ясно, что факт стоит в ближайшем отношении к различным вопросам, касающимся фермента: действия щелочей, различного состояния фермента и т. д., то его изучение составило предмет дальнейшего и сейчас ведущегося в лаборатории исследования.
Весь изложенный материал, как нам кажется, отнимает всякую почву у ходячего мнения, что белки растворяющее и молоко свертывающее ферментные действия принадлежат особым ферментам. Факт тот, что эти действия никогда прочно не разделимы ни в организме, ни вне его ни при каких условиях. Есть, понятно, еще некоторые частные указания на прочную раздельность действий, но, не проверив их еще до сих пор, мы считали себя, однако, вправе не придавать им особенной силы ввиду обнаруженных нами недостатков современной методики сравнения этих действий.
Но мы давно уже остановились на общем вопросе: не противоречит ли нашему выводу, что оба исследуемые действия принадлежат одному и тому жерменту, то обстоятельство, что эти действия обнаруживаются по двум различным правилам зависимости от количества фермента? Это же представлялось существенным возражением против нашего взгляда и Гамарстену, что мы прочли уже во время писания этой статьи в последнем издании его учебника физиологической химии.
Наши обе реакции - растворение белка и свертывание молока - во всяком случае разные реакции, а если это так, то нет никакой необходимости в тождестве правил количественной зависимости времени реакции от концентрации ферментов. Скорость реакции определяется условиями реакции, которые то увеличивают, то уменьшают скорость. Соответственно характеру условий должны изменяться и правила зависимости скорости реакции от количества фермента в данном случае. С целью получить фактический материал, подтверждающий это, мы поставили несколько рядов опытов, где, меняя условия реакций, достигли соответствующего изменения правил. Нейтральный раствор молоко свертывающего фермента, раз он находится в жидкости в известной достаточной концентрации, свертывает молоко по правилу обратной пропорциональности между количествами фермента и временами свертывания. Хотя это давно известно, но мы позволяем себе, ради наглядного сравнения с последующим, привести таблицу (18), относящуюся к этому опыту.
Если мы берем кислый раствор того же фермента и прибавляем различное количество его к молоку, то теперь мы изменяем в молоке в совершенно той же пропорции количество как фермента, так и кислоты, т. е. если мы, например, увеличим вдвое количество фермента, то вместе с тем увеличиваем вдвое и количество кислоты в нем.
 В таком случае первое правило обращается во второе: именно теперь количества фермента относятся обратно пропорционально, как квадратные корни их времен свертывания, т. е. реакция правильно ускоряется. Привожу таблицу (19).
В таком случае первое правило обращается во второе: именно теперь количества фермента относятся обратно пропорционально, как квадратные корни их времен свертывания, т. е. реакция правильно ускоряется. Привожу таблицу (19).
 Совершенно то же самое наблюдается и вом случае, если в растворе фермента имеются соли, ускоряющие свертывание. Если, например, натуральный желудочный сок нейтрализовался углекислым барием и такая нейтрализованная жидкость приливается в разных количествах к молоку, то она также свертывает молоко по второму правилу. Приводим таблицу (20).
Совершенно то же самое наблюдается и вом случае, если в растворе фермента имеются соли, ускоряющие свертывание. Если, например, натуральный желудочный сок нейтрализовался углекислым барием и такая нейтрализованная жидкость приливается в разных количествах к молоку, то она также свертывает молоко по второму правилу. Приводим таблицу (20).
 Наконец, если взять значительно разведенный нейтральный раствор фермента и приливать его к молоку разными, но большими количествами, чтобы избежать предела, то часто наблюдается действие третьего правила: количества ферментов относятся теперь, как квадраты времен свертывания. Очевидно, разведение молока, идущее строго пропорционально вместе с увеличением фермента, правильно замедляет скорость реакции. Приводим таблицу (21).
Наконец, если взять значительно разведенный нейтральный раствор фермента и приливать его к молоку разными, но большими количествами, чтобы избежать предела, то часто наблюдается действие третьего правила: количества ферментов относятся теперь, как квадраты времен свертывания. Очевидно, разведение молока, идущее строго пропорционально вместе с увеличением фермента, правильно замедляет скорость реакции. Приводим таблицу (21).
 Совершенно так же и правило растворения белка в зависимости от количества фермента может видоизменяться, смотря по условиям реакции. Обыкновенно в известных пределах концентрации фермента количество фермента связано с действием числом миллиметров переваренной белковой палочки (по Месту) за одно и то же время - правилом прямой пропорциональности между количеством фермента и квадратом числа миллиметров. Опять для наглядного сравнения приводим таблицу (22).
Совершенно так же и правило растворения белка в зависимости от количества фермента может видоизменяться, смотря по условиям реакции. Обыкновенно в известных пределах концентрации фермента количество фермента связано с действием числом миллиметров переваренной белковой палочки (по Месту) за одно и то же время - правилом прямой пропорциональности между количеством фермента и квадратом числа миллиметров. Опять для наглядного сравнения приводим таблицу (22).
 Если же, беря слабые сорта сока, разбавлять его не кислотой, как делалось в предшествующем опыте, а водой, то связь количества фермента с действием (числом миллиметров переваренной белковой палочки) определяет новое правило, правило прямой пропорциональности между количеством фермента и числом (а не квадратом его) миллиметров переваренной белковой палочки. Приводим таблицу (23).
Последнее отношение уясняет основной смысл и первого правила, относящегося до реакции растворения белка. Очевидно, когда мы кислотой разбавляем данный раствор фермента, мы изменяем не один фактор, как нам казалось раньше, - именно концентрацию фермента, а два: кроме изменения количества фермента в среде, мы изменяем, - именно правильно увеличиваем, - количество кислоты относительно фермента, чем правильно ускоряем реакцию. При разведении же водой действительно изменяется в среде тольколичество фермента, отношение же концентрации фермента к концентрации кислоты остается постоянным, т. е. реакция не испытывает ускоряющего влияния. Это и будет случай реакции растворения белка, совершенно аналогичный обыкновенному случаю свертывания молока чистым нейтральным раствором фермента.
Если же, беря слабые сорта сока, разбавлять его не кислотой, как делалось в предшествующем опыте, а водой, то связь количества фермента с действием (числом миллиметров переваренной белковой палочки) определяет новое правило, правило прямой пропорциональности между количеством фермента и числом (а не квадратом его) миллиметров переваренной белковой палочки. Приводим таблицу (23).
Последнее отношение уясняет основной смысл и первого правила, относящегося до реакции растворения белка. Очевидно, когда мы кислотой разбавляем данный раствор фермента, мы изменяем не один фактор, как нам казалось раньше, - именно концентрацию фермента, а два: кроме изменения количества фермента в среде, мы изменяем, - именно правильно увеличиваем, - количество кислоты относительно фермента, чем правильно ускоряем реакцию. При разведении же водой действительно изменяется в среде тольколичество фермента, отношение же концентрации фермента к концентрации кислоты остается постоянным, т. е. реакция не испытывает ускоряющего влияния. Это и будет случай реакции растворения белка, совершенно аналогичный обыкновенному случаю свертывания молока чистым нейтральным раствором фермента.
 Таким образом казавшееся существенным возражение потеряло свою силу. Самый же факт варьирования правил зависимости действий фермента от количества, смотря по условиям, конечно, требует более широкого и систематического исследования, так как все эти правила ограничены определенными пределами.
Не незначительным общим подтверждением правильности руководящей идеи работы служит, помимо всех приведенных частных фактов, и то, что преследование этой мысли, и именно там, где, повидимому, сначала положение дела было решительно против нее, привело к находке новых фактов значительной важности.
По всему приведенному продолжать думать, что оба наши действия исходят от разных ферментов, но всегда в организмах, встречающихся вместе, и притом в одних и тех же количественных отношениях, было бы только привычкой к старому выводу, лишившемуся в настоящее время своих оснований.
Таким образом мы утверждаем, что оба действия принадлежат одному и тому же ферменту. Как понимать эту двойственность действия одного и того же фермента?
Конечно, первый пункт, с которого надо начинать, касается реакции свертывания молока: чтоно такое физиологически и химически? Нельзя не признать, что реакция эта долгое время была совершенно загадочной, что касается ее физиологического смысла. Если у млекопитающих животных она могла быть до известной степени истолковываема как выгодная или надобная при обработке молока в пищеварительном канале, то у всех остальных животных и растений, где совсем не встречается субстрата для молочного фермента, назначение ее было совершенно темно. Мне кажется, в этом нельзя не видеть яркого примера бесстрастного и, конечно уж, незаконного объективизма, который спокойно проходит около в высшей степени курьезного факта, не задаваясь, повидимому, необходимым вопросом: для чего же существует в том или другом животном или растении молочный фермент, который никогда не встречается с молоком? При настойчивой постановке такого вопроса неизбежно следовало бы заключение, что свертывание молока есть частное, для экспериментатора как бы случайное, обнаружение какой-то общей реакции. Честь такого заключения по справедливости, по моему мнению, должна быть приписана профессору А. Данилевскому, который в свертывании молока видел усложнение белковой частицы, обратное тому упрощению, которое наступает при действии белковых ферментов, как пепсин и т. д. И действительно, в соответствии с этим, ему удалось, вместе с учеником его Окуновым, показать и осаждение некоторых альбумоз препаратом сычужного фермента. В самом деле, что могло бы говорить против предположения, что свертывание молока есть наиболее простая стадия из длинного синтеза белковых тел? Против серьезного ничего, а за - нечто есть. Казеин и параказеин всеми признаются за очень близкие химические тела. А между тем, в той же обстановке, одно остается творе, а другое осаждается. Вполне естественно думать в таком случае о более сложной частице второго вещества. Этому представлению в высшей степени благоприятствуют только что опубликованные опыты Ловенгарта. [59] Теперь, когда у старого мнения, что белок растворяющее и молоко свертывающее действия принадлежат разным ферментам, отнято всякое основание, представляется самым естественным думать, что обе реакции суть обратимые реакции белкового фермента. Если предвидимая уже теоретически обратимость ферментных реакций вполне подтвердилась на жировых и углеводных ферментах, то почему бы белковый фермент представлял исключение? А если она должна быть и у него, то зачем ее искать далеко, когда у того же белкового фермента есть загадочная сторона действия, всего проще понимаемая именно с этой точки зрения.
Надо думать, к этой же категории явлений принадлежат и свертывание крови и желатинизирование панкреатического сока, описанные в работе доктора Линтварева.2 [60] Если в зимогенный панкреатический сок, содержащий много белка, прибавляется киназа, то сок превращается часто быстро в такой прозрачноплотный сгусток, что пробирку можно перевернуть вверх дном без того, чтобы содержимое ее вылилось.
Смеем думать, что с признанием обратимых реакций белковых ферментов, которых мы имеем теперь целый ряд, вероятно для различных стадий разложения белка, биологическая химия выйдет на прямую дорогу, ведущую к разрешению ее важнейшего вопроса о синтезе белкового вещества.
Помимо указанного общего значения, доказанная принадлежность обеих наших реакций одному и тому же ферменту обещает большое методическое применение, которым мы уже не раз и пользовались. Для определения количества белкового фермента молоко свертывающее действие представляет огромные выгоды сравнительно с реакцией растворения белка. Она протекает гораздо быстрей, лучше измерима и менее зависима в химическом отношении (более проста).
Мы не приводим обширную литературу по свертыванию молока, потому что не имеем претензии на открытие какихлибо новых фактов, к свертыванию молока относящихся. Центр тяжести нашей работы лежит в применении давно известных фактов к улучшению сравнительной методики исследования белок растворяющего и молоко свертывающего ферментов.
Но зато мы не хотели бы оставлять неясности относительно независимости нашей мысли о тождестве ферментов, свертывающих белок. Появившееся в заграничной прессе раньше заключение Зибера и Ненцкого, 1 [61] с одной стороны, и Пекальхаринга,2 [62] 2 с другой, - о тождестве химозина и пепсина, во-первых, есть нечто другое: эти авторы допускают, так сказать, частичное тождество этих ферментов, принимая разные добавочные группы для каждого отдельного фермента. К тому же почти никаких доказательств (кроме беглого указания на немногие случаи параллелизма) ими не было приведено в пользу этого заключения. Скорее это заключение нужно было авторам для того, чтобы иметь право говорить о полученном ими продукте из натурального желудочного сока, как о химическом индивидууме. Во-вторых, один из нас (И. Павлов) высказал мысль о тождестве этих ферментов в том смысле, как она развита в настоящем исследовании, ранее публикации упомянутых авторов. 3 [63] Заметка эта вызвана тем, что некоторые авторы считают нашу основную мысль и наши данные дальнейшим развитием заключений вышеприведенных авторов.
Приведенные выше результаты нашей работы в коротких сообщениях были заявлены в нескольких заседаниях Общества русских врачей 1902 и 1904 гг., на Съезде натуралистов в Гельсингфорсе в 1902 г. и на Международном съезде в Мадриде в 1903 г.
В постановке последних опытов относительно пропорциональности обоих действий большое участие принимал доктор Н. П. Тихомиров, за что мы приносим ему нашу искреннюю благодарность.
Таким образом казавшееся существенным возражение потеряло свою силу. Самый же факт варьирования правил зависимости действий фермента от количества, смотря по условиям, конечно, требует более широкого и систематического исследования, так как все эти правила ограничены определенными пределами.
Не незначительным общим подтверждением правильности руководящей идеи работы служит, помимо всех приведенных частных фактов, и то, что преследование этой мысли, и именно там, где, повидимому, сначала положение дела было решительно против нее, привело к находке новых фактов значительной важности.
По всему приведенному продолжать думать, что оба наши действия исходят от разных ферментов, но всегда в организмах, встречающихся вместе, и притом в одних и тех же количественных отношениях, было бы только привычкой к старому выводу, лишившемуся в настоящее время своих оснований.
Таким образом мы утверждаем, что оба действия принадлежат одному и тому же ферменту. Как понимать эту двойственность действия одного и того же фермента?
Конечно, первый пункт, с которого надо начинать, касается реакции свертывания молока: чтоно такое физиологически и химически? Нельзя не признать, что реакция эта долгое время была совершенно загадочной, что касается ее физиологического смысла. Если у млекопитающих животных она могла быть до известной степени истолковываема как выгодная или надобная при обработке молока в пищеварительном канале, то у всех остальных животных и растений, где совсем не встречается субстрата для молочного фермента, назначение ее было совершенно темно. Мне кажется, в этом нельзя не видеть яркого примера бесстрастного и, конечно уж, незаконного объективизма, который спокойно проходит около в высшей степени курьезного факта, не задаваясь, повидимому, необходимым вопросом: для чего же существует в том или другом животном или растении молочный фермент, который никогда не встречается с молоком? При настойчивой постановке такого вопроса неизбежно следовало бы заключение, что свертывание молока есть частное, для экспериментатора как бы случайное, обнаружение какой-то общей реакции. Честь такого заключения по справедливости, по моему мнению, должна быть приписана профессору А. Данилевскому, который в свертывании молока видел усложнение белковой частицы, обратное тому упрощению, которое наступает при действии белковых ферментов, как пепсин и т. д. И действительно, в соответствии с этим, ему удалось, вместе с учеником его Окуновым, показать и осаждение некоторых альбумоз препаратом сычужного фермента. В самом деле, что могло бы говорить против предположения, что свертывание молока есть наиболее простая стадия из длинного синтеза белковых тел? Против серьезного ничего, а за - нечто есть. Казеин и параказеин всеми признаются за очень близкие химические тела. А между тем, в той же обстановке, одно остается творе, а другое осаждается. Вполне естественно думать в таком случае о более сложной частице второго вещества. Этому представлению в высшей степени благоприятствуют только что опубликованные опыты Ловенгарта. [59] Теперь, когда у старого мнения, что белок растворяющее и молоко свертывающее действия принадлежат разным ферментам, отнято всякое основание, представляется самым естественным думать, что обе реакции суть обратимые реакции белкового фермента. Если предвидимая уже теоретически обратимость ферментных реакций вполне подтвердилась на жировых и углеводных ферментах, то почему бы белковый фермент представлял исключение? А если она должна быть и у него, то зачем ее искать далеко, когда у того же белкового фермента есть загадочная сторона действия, всего проще понимаемая именно с этой точки зрения.
Надо думать, к этой же категории явлений принадлежат и свертывание крови и желатинизирование панкреатического сока, описанные в работе доктора Линтварева.2 [60] Если в зимогенный панкреатический сок, содержащий много белка, прибавляется киназа, то сок превращается часто быстро в такой прозрачноплотный сгусток, что пробирку можно перевернуть вверх дном без того, чтобы содержимое ее вылилось.
Смеем думать, что с признанием обратимых реакций белковых ферментов, которых мы имеем теперь целый ряд, вероятно для различных стадий разложения белка, биологическая химия выйдет на прямую дорогу, ведущую к разрешению ее важнейшего вопроса о синтезе белкового вещества.
Помимо указанного общего значения, доказанная принадлежность обеих наших реакций одному и тому же ферменту обещает большое методическое применение, которым мы уже не раз и пользовались. Для определения количества белкового фермента молоко свертывающее действие представляет огромные выгоды сравнительно с реакцией растворения белка. Она протекает гораздо быстрей, лучше измерима и менее зависима в химическом отношении (более проста).
Мы не приводим обширную литературу по свертыванию молока, потому что не имеем претензии на открытие какихлибо новых фактов, к свертыванию молока относящихся. Центр тяжести нашей работы лежит в применении давно известных фактов к улучшению сравнительной методики исследования белок растворяющего и молоко свертывающего ферментов.
Но зато мы не хотели бы оставлять неясности относительно независимости нашей мысли о тождестве ферментов, свертывающих белок. Появившееся в заграничной прессе раньше заключение Зибера и Ненцкого, 1 [61] с одной стороны, и Пекальхаринга,2 [62] 2 с другой, - о тождестве химозина и пепсина, во-первых, есть нечто другое: эти авторы допускают, так сказать, частичное тождество этих ферментов, принимая разные добавочные группы для каждого отдельного фермента. К тому же почти никаких доказательств (кроме беглого указания на немногие случаи параллелизма) ими не было приведено в пользу этого заключения. Скорее это заключение нужно было авторам для того, чтобы иметь право говорить о полученном ими продукте из натурального желудочного сока, как о химическом индивидууме. Во-вторых, один из нас (И. Павлов) высказал мысль о тождестве этих ферментов в том смысле, как она развита в настоящем исследовании, ранее публикации упомянутых авторов. 3 [63] Заметка эта вызвана тем, что некоторые авторы считают нашу основную мысль и наши данные дальнейшим развитием заключений вышеприведенных авторов.
Приведенные выше результаты нашей работы в коротких сообщениях были заявлены в нескольких заседаниях Общества русских врачей 1902 и 1904 гг., на Съезде натуралистов в Гельсингфорсе в 1902 г. и на Международном съезде в Мадриде в 1903 г.
В постановке последних опытов относительно пропорциональности обоих действий большое участие принимал доктор Н. П. Тихомиров, за что мы приносим ему нашу искреннюю благодарность.
 Затем, как каждый знает из собственного опыта, слюна часто служит, как и вода, для разведения слишком концентрированных растворов из попадающих в рот веществ, а также для отмывания вредоносных или тошнотворных веществ из ротовой полости. С этим вполне согласуется тот факт, что на отвергаемые животным вещества из слизистых слюнных желез всегда изливается лишь жидкая слюна; ведь, слизь была бы в этих случаях совершенно не нужна. Из различных отвергаемых животным химических веществ некоторые требуют, вследствие своего особенно сильного химического действия, совершенно особых мероприятий; эти меры, очевидно, принимаются околоушной железой, которая выделяет на кислоту и на щелочи, в противоположность всем другим сильно раздражающим веществам, слюну, очень богатую белком; этим до некоторой степени уменьшается разрушающее действие этих веществ на слизистую рта. Что всеми приведенными мерами вредное влияние некоторых веществ действительно нейтрализуется, может быть доказано следующими фактами. Можно много раз и большими порциями вливать собаке в рот 0.5% раствор соляной кислоты, не наблюдая при этом даже малейшего повреждения слизистой рта; если же окунуть язык собаки на несколько минут в чашку, наполненную подобным раствором, то поверхностный слой эпителия сойдет, совсем как при обваривании горячей жидкостью.
Интересно отношение к нерастворимым твердым телам. Камешки не вызывают никакой слюнной секреции, на песок же слюна изливается очень обильно. Ясно, что удаление камешков из ротовой полости не может происходить при помощи слюны; камешки, даже если бы они пристали к стенке ротовой полости, могут быть удалены из нее благодаря своему весу и при помощи мышечного аппарата рта. Песок, наоборот, не мог бы быть легко удален из ротовой полости без помощи жидкости; благодаря примеси слизистой жидкости он соединяется в небольшие кучки, которые выплевываются; отдельные же оставшиеся песчинки выносятся потоком водянистой слюны.
Вода и физиологический раствор поваренной соли не вызывают никакой секреции, что и вполне понятно: к чему бы тут послужила слюна?
Тем, что было до сих пор изложено, далеко не исчерпываются отношения слюнных желез к объектам внешнего мира; существует еще целая группа непосредственно сюда относящихся явлений, которые, однако, как будто совершенно другого рода. Объекты внешнего мира, о которых до сих пор шла речь, действуют на работу слюнных желез также с некоторого расстояния, т. е. и тогда, когда они еще не попали в ротовую полость, и, стало быть через посредство других чувствительных поверхностей - носа, глаза, уха. Это - так называемое психическое раздражение слюнных желез. Уже каждодневное наблюдение издавна установило факт слюноотделения при виде пищи вак у человека, так и у животных. Совершенно так же это явление повторно наблюдалось у человека при виде некоторых неприятных веществ. За последнее время Вульфсон, выводы которого были подтверждены всеми позднейшими авторами (Малуазель, Снарский, Толочинов и т. д.), подверг этот вопрос экспериментальному изучению на животных. Вывод из его опытов был тот, что все вещества, разнообразно влияющие на работу слюнных желез, совершенно так же, но лишь в меньшем размере, влияют и на расстоянии. Сухой мясо-сухарный порошок вызывает у собаки более обильное слюноотделение, чем мясо, хотя животное, очевидно, может быть, гораздо больше заинтересовано видомяса. Вид кислоты вызывает у собаки секрецию жидкой слюны из слизистых слюнных желез, а вид хлеба - секрецию более концентрированной слюны и т. д. Однако Зельгейму удалось установить исключение из этого правила: при виде кислот и щелочей у собаки вместе с секретом околоушной слюнной железы выделяется не больше органических веществ, чем солей, как это наблюдается при их введении в ротовую полость, а меньше.
Случаи так называемого психического раздражения слюнных желез удобнее всего рассматривать как дальнейшее приспособление этих желез к внешнему миру; стоит только данные объекты приблизить ко рту, как эти органы уже начинают превентивно работать соответствующим образом; это может повести в каждом специальном случае лишь к наилучшему выполнению физиологической цели этой работы. Если, например, ко рту животного приблизить отвергаемые вещества, то, с одной стороны, благодаряпредупредительной слюнной секреции, смоченная слизистая защищается от непосредственного соприкосновения с веществом, a, с другой стороны, это последнее тотчас же разжижается и становится более или менее безвредным.
Всем тем, что упомянуто выше, мы, естественно, еще далеко не исчерпали те тонкие, специальные отношения, которые существуют между химическими свойствами, формой и физическим состоянием попадающих в ротовую полость веществ и особым ходом работы слюнных желез. Мы знаем очень мало о всех ферментах слюны и о колебаниях их содержания при различных условиях; так же и остальные составные части слюны для начала анализированы лишь в грубых чертах. Но в общем надо допустить, что все химические и физические свойства слюны (как и каждого физиологического объекта) должны иметь свое применение, свое место и свое значение в системе организма. Уже установленных фактов достаточно, однако, чтобы не оставить никакого сомнения относительно общего положения о специфичности и приспособляемости слюнных желез, функционирующих как пищеварительные органы.
Кроме их участия в пищеварительной системе, слюнные железы с их секретом служат организму еще и иным образом. Так, мы видим, например, что животные обмываются слюной и пользуются ею при внешних повреждениях. Не невероятно, что в последнем случае применение слюны имеет лечебное значение, причем рана не только очищается, но и защищается слоем слюны от внешнего мира и, может быть, вместе с тем приобретает нечто исцеляющее (оксидаза?). В согласии с этим наблюдением мы видим, как у фистульных животных при каждом разрушающем воздействии на кожу (ожоги, уколы и т. д.) наступает более или менее значительная слюнная секреция. То же явление наблюдается также каждый раз, когда животному показывают, что собираются нанести ему кожное повреждение, т. е. под действием психического эффекта (Толочинов 1). [73]
Наконец следует еще упомянуть, что и некоторые внутренние процессы, как, например, задушение, отравление собственными или извне введенными ядами, вызывают работу слюнных желез. Физиологическое значение этих явлений заключается, вероятно, в том, что некоторые вещества удаляются из организма со слюной.
Затем, как каждый знает из собственного опыта, слюна часто служит, как и вода, для разведения слишком концентрированных растворов из попадающих в рот веществ, а также для отмывания вредоносных или тошнотворных веществ из ротовой полости. С этим вполне согласуется тот факт, что на отвергаемые животным вещества из слизистых слюнных желез всегда изливается лишь жидкая слюна; ведь, слизь была бы в этих случаях совершенно не нужна. Из различных отвергаемых животным химических веществ некоторые требуют, вследствие своего особенно сильного химического действия, совершенно особых мероприятий; эти меры, очевидно, принимаются околоушной железой, которая выделяет на кислоту и на щелочи, в противоположность всем другим сильно раздражающим веществам, слюну, очень богатую белком; этим до некоторой степени уменьшается разрушающее действие этих веществ на слизистую рта. Что всеми приведенными мерами вредное влияние некоторых веществ действительно нейтрализуется, может быть доказано следующими фактами. Можно много раз и большими порциями вливать собаке в рот 0.5% раствор соляной кислоты, не наблюдая при этом даже малейшего повреждения слизистой рта; если же окунуть язык собаки на несколько минут в чашку, наполненную подобным раствором, то поверхностный слой эпителия сойдет, совсем как при обваривании горячей жидкостью.
Интересно отношение к нерастворимым твердым телам. Камешки не вызывают никакой слюнной секреции, на песок же слюна изливается очень обильно. Ясно, что удаление камешков из ротовой полости не может происходить при помощи слюны; камешки, даже если бы они пристали к стенке ротовой полости, могут быть удалены из нее благодаря своему весу и при помощи мышечного аппарата рта. Песок, наоборот, не мог бы быть легко удален из ротовой полости без помощи жидкости; благодаря примеси слизистой жидкости он соединяется в небольшие кучки, которые выплевываются; отдельные же оставшиеся песчинки выносятся потоком водянистой слюны.
Вода и физиологический раствор поваренной соли не вызывают никакой секреции, что и вполне понятно: к чему бы тут послужила слюна?
Тем, что было до сих пор изложено, далеко не исчерпываются отношения слюнных желез к объектам внешнего мира; существует еще целая группа непосредственно сюда относящихся явлений, которые, однако, как будто совершенно другого рода. Объекты внешнего мира, о которых до сих пор шла речь, действуют на работу слюнных желез также с некоторого расстояния, т. е. и тогда, когда они еще не попали в ротовую полость, и, стало быть через посредство других чувствительных поверхностей - носа, глаза, уха. Это - так называемое психическое раздражение слюнных желез. Уже каждодневное наблюдение издавна установило факт слюноотделения при виде пищи вак у человека, так и у животных. Совершенно так же это явление повторно наблюдалось у человека при виде некоторых неприятных веществ. За последнее время Вульфсон, выводы которого были подтверждены всеми позднейшими авторами (Малуазель, Снарский, Толочинов и т. д.), подверг этот вопрос экспериментальному изучению на животных. Вывод из его опытов был тот, что все вещества, разнообразно влияющие на работу слюнных желез, совершенно так же, но лишь в меньшем размере, влияют и на расстоянии. Сухой мясо-сухарный порошок вызывает у собаки более обильное слюноотделение, чем мясо, хотя животное, очевидно, может быть, гораздо больше заинтересовано видомяса. Вид кислоты вызывает у собаки секрецию жидкой слюны из слизистых слюнных желез, а вид хлеба - секрецию более концентрированной слюны и т. д. Однако Зельгейму удалось установить исключение из этого правила: при виде кислот и щелочей у собаки вместе с секретом околоушной слюнной железы выделяется не больше органических веществ, чем солей, как это наблюдается при их введении в ротовую полость, а меньше.
Случаи так называемого психического раздражения слюнных желез удобнее всего рассматривать как дальнейшее приспособление этих желез к внешнему миру; стоит только данные объекты приблизить ко рту, как эти органы уже начинают превентивно работать соответствующим образом; это может повести в каждом специальном случае лишь к наилучшему выполнению физиологической цели этой работы. Если, например, ко рту животного приблизить отвергаемые вещества, то, с одной стороны, благодаряпредупредительной слюнной секреции, смоченная слизистая защищается от непосредственного соприкосновения с веществом, a, с другой стороны, это последнее тотчас же разжижается и становится более или менее безвредным.
Всем тем, что упомянуто выше, мы, естественно, еще далеко не исчерпали те тонкие, специальные отношения, которые существуют между химическими свойствами, формой и физическим состоянием попадающих в ротовую полость веществ и особым ходом работы слюнных желез. Мы знаем очень мало о всех ферментах слюны и о колебаниях их содержания при различных условиях; так же и остальные составные части слюны для начала анализированы лишь в грубых чертах. Но в общем надо допустить, что все химические и физические свойства слюны (как и каждого физиологического объекта) должны иметь свое применение, свое место и свое значение в системе организма. Уже установленных фактов достаточно, однако, чтобы не оставить никакого сомнения относительно общего положения о специфичности и приспособляемости слюнных желез, функционирующих как пищеварительные органы.
Кроме их участия в пищеварительной системе, слюнные железы с их секретом служат организму еще и иным образом. Так, мы видим, например, что животные обмываются слюной и пользуются ею при внешних повреждениях. Не невероятно, что в последнем случае применение слюны имеет лечебное значение, причем рана не только очищается, но и защищается слоем слюны от внешнего мира и, может быть, вместе с тем приобретает нечто исцеляющее (оксидаза?). В согласии с этим наблюдением мы видим, как у фистульных животных при каждом разрушающем воздействии на кожу (ожоги, уколы и т. д.) наступает более или менее значительная слюнная секреция. То же явление наблюдается также каждый раз, когда животному показывают, что собираются нанести ему кожное повреждение, т. е. под действием психического эффекта (Толочинов 1). [73]
Наконец следует еще упомянуть, что и некоторые внутренние процессы, как, например, задушение, отравление собственными или извне введенными ядами, вызывают работу слюнных желез. Физиологическое значение этих явлений заключается, вероятно, в том, что некоторые вещества удаляются из организма со слюной.
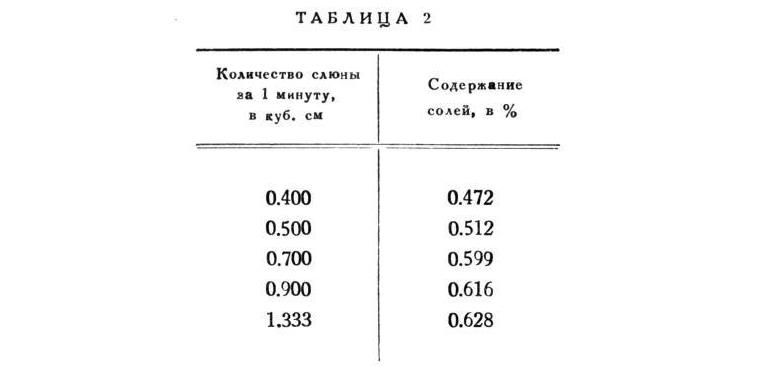 Те же отношения можно констатировать и на табл. 1 при нормальной слюнной секреции: несмотря на разнородность раздражений, процентное содержание солей всегда параллельно быстроте секреции.
Что касается процентного содержания органических веществ, то здесь отношения складываются гораздо сложнее. Если раздражать железу в состоянии покоя, то при каждом увеличении железистой секреции, вызванном усилением раздражения, одновременно увеличивается и процентное содержание органических веществ в слюне. Но так как при непрерывно продолжающейся секреции железа истощается, то это отношение не сохраняется, т. е. теперь при усилении секреции процентное содержание органических веществ остается либо тем же, либо даже уменьшается (Гейденгайн [87] +).
Доказательством этого пусть послужит табл. 3.
Те же отношения можно констатировать и на табл. 1 при нормальной слюнной секреции: несмотря на разнородность раздражений, процентное содержание солей всегда параллельно быстроте секреции.
Что касается процентного содержания органических веществ, то здесь отношения складываются гораздо сложнее. Если раздражать железу в состоянии покоя, то при каждом увеличении железистой секреции, вызванном усилением раздражения, одновременно увеличивается и процентное содержание органических веществ в слюне. Но так как при непрерывно продолжающейся секреции железа истощается, то это отношение не сохраняется, т. е. теперь при усилении секреции процентное содержание органических веществ остается либо тем же, либо даже уменьшается (Гейденгайн [87] +).
Доказательством этого пусть послужит табл. 3.
 В первых четырех опытах с усилением раздражения нарастают как быстрота секреции, так и процентное содержание органических веществ; но в опытах 6 и 8 усиление раздражения, наряду с ускорением секреции, ведет к уменьшению процентного содержания органических веществ, и только в опыте 9, где раздражение было усилено очень значительно, вновь устанавливается прежнее отношение. Процентное содержание солей каждый раз нарастает вместе с быстротой секреции.
To Тогда как процентное содержание солей всегда идет параллельно силе раздражения и быстроте секреции, действие сильного раздражения, вызвавшего увеличение содержания солей, не ограничивается количеством слюны, которая при этом выделилась, но сказывается и на следующем за ним более слабом раздражении; а именно: на него выделяется слюна с более значительным содержанием органических веществ, чем на такое же раздражение, но имевшее место до сильного раздражения (Тейденгайн 1). [88]
Табл. 4 иллюстрирует эти отношения.
В первых четырех опытах с усилением раздражения нарастают как быстрота секреции, так и процентное содержание органических веществ; но в опытах 6 и 8 усиление раздражения, наряду с ускорением секреции, ведет к уменьшению процентного содержания органических веществ, и только в опыте 9, где раздражение было усилено очень значительно, вновь устанавливается прежнее отношение. Процентное содержание солей каждый раз нарастает вместе с быстротой секреции.
To Тогда как процентное содержание солей всегда идет параллельно силе раздражения и быстроте секреции, действие сильного раздражения, вызвавшего увеличение содержания солей, не ограничивается количеством слюны, которая при этом выделилась, но сказывается и на следующем за ним более слабом раздражении; а именно: на него выделяется слюна с более значительным содержанием органических веществ, чем на такое же раздражение, но имевшее место до сильного раздражения (Тейденгайн 1). [88]
Табл. 4 иллюстрирует эти отношения.
 При прочих равных условиях процентное содержание органических веществ падает тем сильнее, чем дольше длится раздражение (Бехер и Людвиг 1). [89]
Табл. 5 заимствована из сообщений этих авторов.
При прочих равных условиях процентное содержание органических веществ падает тем сильнее, чем дольше длится раздражение (Бехер и Людвиг 1). [89]
Табл. 5 заимствована из сообщений этих авторов.
 Однако процентное содержание солей изменяется лишь незначительно.
Стало быть, и в случае искусственного возбуждения центрабежных без нервов могут также быть вызваны значительные колебания в содержании органических веществ в слюне совершенно так же, как это чрезвычайно ярко можно наблюдать при нормальном возбуждении деятельности слюнных желез (ср. табл. 1).
При прогрессивном, нарастающем сужении сосудов, снабжающих железу кровью, во время раздражения chorda при прочих равных условиях количество слюны постепенно уменьшается и может упасть до нуля, причем и после восстановления кровообращения прежнее раздражение лишь постепенно достигает своего прежнего действия; если же кровообращение было прервано надолго, то раздражение вообще может перестать действовать. Содержание органических веществ в слюне при такой задержке кровообращения также не нарастает (Гейденгайн [90]
Доказательством этого служит следующий опыт этого автора: art. carotis обнажена, Р. к. - 150 до 85, 0.28 куб. см в минуту, содержание плогных веществ 1.37%; art. carotis зажата, Р. к. - 85 до 70, 0.13 куб. см в минуту содержание плотных веществ 1.33%.
Однако при несколько измененных условиях могли быть достигнуты результаты, которые говорят несколько иное в смысле содержания органических веществ. Если впрыснуть животному пилокарпин, который значительно возбуждает слюнную секрецию, то после кровопускания выделяется слюна с значительно С большим содержанием органических солей, чем до него (Лэнгли, [91] Флетчер) (ср. табл. 6, заимствованную из сообщений этих авторов; стр. 478).
При возбуждении chorda tympani после перерезки спинного мозга, следствием которой, как известно, является значительное замедление кровообращения, слюна также выделяется гораздо медленнее и содержит больше органических веществ (Цернер 3 [92] 9).
При сужении сосудов и кровопусканиях процентное содержание солей повышается.
В высшей степени желательно и интересно установить дальнейшими опытами, как выразятся здесь отношения, почему два рода по существу, казалось бы, однородных опытов ведут к различным результатам.
Однако процентное содержание солей изменяется лишь незначительно.
Стало быть, и в случае искусственного возбуждения центрабежных без нервов могут также быть вызваны значительные колебания в содержании органических веществ в слюне совершенно так же, как это чрезвычайно ярко можно наблюдать при нормальном возбуждении деятельности слюнных желез (ср. табл. 1).
При прогрессивном, нарастающем сужении сосудов, снабжающих железу кровью, во время раздражения chorda при прочих равных условиях количество слюны постепенно уменьшается и может упасть до нуля, причем и после восстановления кровообращения прежнее раздражение лишь постепенно достигает своего прежнего действия; если же кровообращение было прервано надолго, то раздражение вообще может перестать действовать. Содержание органических веществ в слюне при такой задержке кровообращения также не нарастает (Гейденгайн [90]
Доказательством этого служит следующий опыт этого автора: art. carotis обнажена, Р. к. - 150 до 85, 0.28 куб. см в минуту, содержание плогных веществ 1.37%; art. carotis зажата, Р. к. - 85 до 70, 0.13 куб. см в минуту содержание плотных веществ 1.33%.
Однако при несколько измененных условиях могли быть достигнуты результаты, которые говорят несколько иное в смысле содержания органических веществ. Если впрыснуть животному пилокарпин, который значительно возбуждает слюнную секрецию, то после кровопускания выделяется слюна с значительно С большим содержанием органических солей, чем до него (Лэнгли, [91] Флетчер) (ср. табл. 6, заимствованную из сообщений этих авторов; стр. 478).
При возбуждении chorda tympani после перерезки спинного мозга, следствием которой, как известно, является значительное замедление кровообращения, слюна также выделяется гораздо медленнее и содержит больше органических веществ (Цернер 3 [92] 9).
При сужении сосудов и кровопусканиях процентное содержание солей повышается.
В высшей степени желательно и интересно установить дальнейшими опытами, как выразятся здесь отношения, почему два рода по существу, казалось бы, однородных опытов ведут к различным результатам.
 Исключительно в интересах дальнейшего анализа нервных воздействий мы приведем еще следующие факты. Весь дальнейший материал, полученный при возбуждении нервов и характеризующий секреторный процесс, должен, естественно, найти место в особом отделе руководства, посвященном внутренней работе желез.
Сюда относится факт, установленный Людвигом одновременно с открытием секреторного действия chorda tympani, что давление, под которым слюна изливается из выводного протока, превосходит давление, существующее одновременно в кровеносных сэсудах. Отсюда следует, что слюнную секреции нельзя рассматривать как процесс фильтрации.
Людвиг 1 [93] далее доказал, что слюнная секреция идет рука об руку с значительной теплопродукцией в железе. Этот факт подтвержден другими авторами, однако позднейшие опыты Бейлиса и Хилла 2 [94] привели к определенно отрицательном результатам, почему и необходимы дальнейшие исследования, которые должны объяснить это противоречие фактических данных.
Действие chorda tympani на железу обнаруживается также и в электрических явлениях; а именно: наблюдаемый при спокойном состоянии железы ток (внешняя поверхность железы ведет себя отрицательно по отношению к хилусу железы) значительно нарастает при возбуждении этого нерва (Бейлис и Брэдфорд, [95] Брэдфорд 2 [96] 2).
Чрезвычайно важное гистологическое открытие сделано Гейденгайном. [97] Этот исследователь показал на соответственно обработанных препаратах, что состояние покоя и деятельность железы резко различаются микроскопически друг от друга. Лэнгли [98] подтвердил эти данные на свежих препаратах, причем оказалось, что находящиеся в состоянии покоя зернистые железистые клетки постепенно теряют свой зернистый вид во время секреторного периода и что на периферии клеток образуется светлая кайма, в которой не видно никаких зерен. По этим микроскопическим картинам заключили, что во время спокойного состояния железы в клетках образуются специфические вещества, накапливающиеся в виде гранул; во время секреции эти вещества служат органической составной частью слюны и откладываются на периФерии клетки в виде новых химических веществ из крови.
В каком же количественном отношении стоят эти процессы друг к другу? Уже вышеупомянутый факт, что содержание органических веществ в слюне падает при длительног секреции, говорит за то, что откладывание нового материала из крови в железу отстает от выделения специфических веществ из железы в слюну. Это подтверждается и микроскопическими данными, так как во время секреции клетки уменьшаются в размере. Дальнейшим доказательством служит тот факт, что работающая железа весит меньше, чем парная покоящаяся, причем первая к тому же показывает, кроме того, более значительное содержание воды (Тейденгайн 1). [99] К этому результату привели количественные определения азота в работающих и покоящихся железах (Павлов, 2 [100] Верховский, [101] Гендерсон [102] *). Если исключить влияние асимметрии желез, то уменьшение общего азота в работающей железе по сравнению с общим азотом покоящейся железа может достигнуть в крайнем случае 26%. Если же прибавить общий азот работающей железы к азоту слюны, то получится цифра, превосходящая цифру общего азота покоящейся железы, т. е. время секреции часть потраченного вещества возмещается воспринятыми из крови веществами. Как процентный размер действительной потери веса железы, так и ее возмещение в значительной мере колеблются, так что может быть поставлен вопрос: как обстоит дело при нормальной секреции - не иначе ли? Верховский вывел из своих цифр заключение, что относительное восстановление железы тем полнее, чем больше органических веществ железа выделила со слюной, т. е. что восстановление тем энергичнее, чем значительнее распад.
При возбуждении chorda в железе увеличивается в 3-4 раза как потребление кислорода, так и выделение углекислоты (Баркрофт [103] 5).
Влияние chorda tympani на клетку парализуется впрыскиванием атропина (Кейхель 9). [104] Если отравление слабое, то при возбуждении chorda tympani уменьшаются как количество слюны, так и содержание органических веществ (Лэнгли 7 "). [105]
После атропинизации потребление кислорода железой при раздражении chorda не изменяется, но зато в течение известного промежутка времени нарастает выделение углекислоты (Баркрофт [106] 1).
Кроме возбуждения слюнной секреции, chorda tympani вызывает усиление кровообращения в железе, так что при электрическом раздражении этого нерва количество вытекающей из вены крови увеличивается во много ра13, кровь принимает светлокрасную окраску и пульсирует при вытекании (Клод Бернар [107] 2). После отравления атропином влияние chorda tympani на кровеносные сосуды сохраняется (Гейденгайн 3 3). [108]
При возбуждении chorda tympani ток лимфы из железы увеличивается; при отравлении атропином этот эффект отсутствует (Бейнбридж 4 [109] +).
При перерезке chorda tympani полная дегенерация происходит гораздо медленнее, чем у многих других нервов, причем у некоторых животных она тянется несколько десятков дней. Это, повидимому, зависит от многочисленных нервных клеток, прерывающих ход нерва к хилусу железы (Лэнгли [110]
Спустя несколько дней после перерезки chorda tympani (через один-три дня) железа начинает непрерывно, в течение нескольких недель, выделять слюну в небольших количествах (Клод Бернар [111] 6) - так называемая паралитическая слюнная секреция. Одновременно железа прогрессивно убавляется в весе, атрофируется.
Диспноэ усиливает, а апноэ уменьшает паралитическую слюнную секрецию (Лэнгли [112] Перерезке шейного симпатического нерва не прекращает этой секреции.
Шейный симпатический нерв всегда возбуждает слюнную секрецию в меньшей степени (и это у всех животных), чем chorda tympani. Секреция начинается довольно энергично, но очень скоро замедляется и, наконец, совсем прекращается, несмотря на продолжающееся возбуждение. Однако можно получить достаточное количество слюны, если тетанизировать нерв с равномерными и часто повторяющимися паузами; приостановившаяся было сначала секреция вновь восстанавливается и продолжается в течение всего времени возбуждения в медленном темпе (Гейденгайн [113] 1).
Другой характерный признак симпатической секреции подчелюстной железы собаки состоит в том, что в этом случае состав слюны другой, а именно, она содержит в 2-3 раза больше органических веществ, чем хордальная слюна (Эккард 2 [114] 2); соответственно с этим слюна вязкая и мутная. Следует при этом упомянуть, что у кошки, наоборот, симпатическая слюна обычно менее концентрирована, чем хордальная (Ленгли 3). [115]
При долго длящейся секреции и в симпатической слюне постепенно уменьшается содержание органических веществ, причем соответственно с этим изменяется и ее вид, так что она постепенно становится похожей на хордальную (Гейденгайн +). [116]
Следующий опыт Гейденгайна иллюстрирует эти отношения: слюна собиралась в течение 4'/2 часов, первая порция содержала 3.734%, 0/ а последняя -- 1.488% органических веществ.
Секреторное давление симпатичеслюны примерно такое же, как хордальной (Лэнгли 8). [117]
Возбуждение симпатического нерва вызывает в железе ток, противоположный в смысле направления тому, который возникает после раздражения chorda tympani, т. е. в этом случае внешняя поверхность железы ведет себя по отношению к хилусу положительно (Брэдфорд 8). [118]
Секреторное действие симпатического нерва собаки либо вовсе не парализуется атропином, либо парализуется очень большими его дозами (Гейденгайн 1). [119] Из сравнения парализующего действия атропина на хорду и отсутствия или слабого действия его на n. sympathicus заключили, что атропин оказывает влияние не на клетки железы, но на периферические окончания нерва, так как на самый нерв он не действует. Так как между атропином и пилокарпином существует взаимный антагонизм, то из этого далее заключают, что пилокарпин, который сильно возбуждает слюнную секрецию даже после дегенерации хорды, имеет свой пункт приложения также в периферических нервных окончаниях.
Симпатический нерв оказывает на кровеносные сосуды железы суживающее действие, которое при сильных раздражениях может совершенно приостановить кровообращение в железе.
Возбуждение n. sympathicus, как и раздражение хорды, усиливает ток лимфы в железе (Бейнбридж 2). [120]
Симпатические нервные волокна прерываются в верхнем шейном ганглии нервными клетками. Если перерезать n. sympathicus на шее, то через 3-4 дня его верхний конец утрачивает свою раздражимость; при возбуждении нерва по ту сторону ганглия его действие опять ясно выступает. Но после экстирпации верхнего шейного ганглия симпатический нерв торяет раздражимость на всем протяжении (Лэнгли [121]
Перерезка n. sympathicus не влечет за собою ни паралитической секреции, ни атрофии железы, но ведот скорее к гипертрофии последней (Брэдфорд +). [122]
Взаимное действие обоих секреторных нервов проявляется во многом.
Введенное между двумя возбуждениями chorda tympani электрическое раздражение n. sympathicus в значительной мере уменьшает содержание органических веществ при втором раздражении, а также и наоборот (Гейденгайн [123] ). Это подтверждается следующим опытом (табл. 7).
Исключительно в интересах дальнейшего анализа нервных воздействий мы приведем еще следующие факты. Весь дальнейший материал, полученный при возбуждении нервов и характеризующий секреторный процесс, должен, естественно, найти место в особом отделе руководства, посвященном внутренней работе желез.
Сюда относится факт, установленный Людвигом одновременно с открытием секреторного действия chorda tympani, что давление, под которым слюна изливается из выводного протока, превосходит давление, существующее одновременно в кровеносных сэсудах. Отсюда следует, что слюнную секреции нельзя рассматривать как процесс фильтрации.
Людвиг 1 [93] далее доказал, что слюнная секреция идет рука об руку с значительной теплопродукцией в железе. Этот факт подтвержден другими авторами, однако позднейшие опыты Бейлиса и Хилла 2 [94] привели к определенно отрицательном результатам, почему и необходимы дальнейшие исследования, которые должны объяснить это противоречие фактических данных.
Действие chorda tympani на железу обнаруживается также и в электрических явлениях; а именно: наблюдаемый при спокойном состоянии железы ток (внешняя поверхность железы ведет себя отрицательно по отношению к хилусу железы) значительно нарастает при возбуждении этого нерва (Бейлис и Брэдфорд, [95] Брэдфорд 2 [96] 2).
Чрезвычайно важное гистологическое открытие сделано Гейденгайном. [97] Этот исследователь показал на соответственно обработанных препаратах, что состояние покоя и деятельность железы резко различаются микроскопически друг от друга. Лэнгли [98] подтвердил эти данные на свежих препаратах, причем оказалось, что находящиеся в состоянии покоя зернистые железистые клетки постепенно теряют свой зернистый вид во время секреторного периода и что на периферии клеток образуется светлая кайма, в которой не видно никаких зерен. По этим микроскопическим картинам заключили, что во время спокойного состояния железы в клетках образуются специфические вещества, накапливающиеся в виде гранул; во время секреции эти вещества служат органической составной частью слюны и откладываются на периФерии клетки в виде новых химических веществ из крови.
В каком же количественном отношении стоят эти процессы друг к другу? Уже вышеупомянутый факт, что содержание органических веществ в слюне падает при длительног секреции, говорит за то, что откладывание нового материала из крови в железу отстает от выделения специфических веществ из железы в слюну. Это подтверждается и микроскопическими данными, так как во время секреции клетки уменьшаются в размере. Дальнейшим доказательством служит тот факт, что работающая железа весит меньше, чем парная покоящаяся, причем первая к тому же показывает, кроме того, более значительное содержание воды (Тейденгайн 1). [99] К этому результату привели количественные определения азота в работающих и покоящихся железах (Павлов, 2 [100] Верховский, [101] Гендерсон [102] *). Если исключить влияние асимметрии желез, то уменьшение общего азота в работающей железе по сравнению с общим азотом покоящейся железа может достигнуть в крайнем случае 26%. Если же прибавить общий азот работающей железы к азоту слюны, то получится цифра, превосходящая цифру общего азота покоящейся железы, т. е. время секреции часть потраченного вещества возмещается воспринятыми из крови веществами. Как процентный размер действительной потери веса железы, так и ее возмещение в значительной мере колеблются, так что может быть поставлен вопрос: как обстоит дело при нормальной секреции - не иначе ли? Верховский вывел из своих цифр заключение, что относительное восстановление железы тем полнее, чем больше органических веществ железа выделила со слюной, т. е. что восстановление тем энергичнее, чем значительнее распад.
При возбуждении chorda в железе увеличивается в 3-4 раза как потребление кислорода, так и выделение углекислоты (Баркрофт [103] 5).
Влияние chorda tympani на клетку парализуется впрыскиванием атропина (Кейхель 9). [104] Если отравление слабое, то при возбуждении chorda tympani уменьшаются как количество слюны, так и содержание органических веществ (Лэнгли 7 "). [105]
После атропинизации потребление кислорода железой при раздражении chorda не изменяется, но зато в течение известного промежутка времени нарастает выделение углекислоты (Баркрофт [106] 1).
Кроме возбуждения слюнной секреции, chorda tympani вызывает усиление кровообращения в железе, так что при электрическом раздражении этого нерва количество вытекающей из вены крови увеличивается во много ра13, кровь принимает светлокрасную окраску и пульсирует при вытекании (Клод Бернар [107] 2). После отравления атропином влияние chorda tympani на кровеносные сосуды сохраняется (Гейденгайн 3 3). [108]
При возбуждении chorda tympani ток лимфы из железы увеличивается; при отравлении атропином этот эффект отсутствует (Бейнбридж 4 [109] +).
При перерезке chorda tympani полная дегенерация происходит гораздо медленнее, чем у многих других нервов, причем у некоторых животных она тянется несколько десятков дней. Это, повидимому, зависит от многочисленных нервных клеток, прерывающих ход нерва к хилусу железы (Лэнгли [110]
Спустя несколько дней после перерезки chorda tympani (через один-три дня) железа начинает непрерывно, в течение нескольких недель, выделять слюну в небольших количествах (Клод Бернар [111] 6) - так называемая паралитическая слюнная секреция. Одновременно железа прогрессивно убавляется в весе, атрофируется.
Диспноэ усиливает, а апноэ уменьшает паралитическую слюнную секрецию (Лэнгли [112] Перерезке шейного симпатического нерва не прекращает этой секреции.
Шейный симпатический нерв всегда возбуждает слюнную секрецию в меньшей степени (и это у всех животных), чем chorda tympani. Секреция начинается довольно энергично, но очень скоро замедляется и, наконец, совсем прекращается, несмотря на продолжающееся возбуждение. Однако можно получить достаточное количество слюны, если тетанизировать нерв с равномерными и часто повторяющимися паузами; приостановившаяся было сначала секреция вновь восстанавливается и продолжается в течение всего времени возбуждения в медленном темпе (Гейденгайн [113] 1).
Другой характерный признак симпатической секреции подчелюстной железы собаки состоит в том, что в этом случае состав слюны другой, а именно, она содержит в 2-3 раза больше органических веществ, чем хордальная слюна (Эккард 2 [114] 2); соответственно с этим слюна вязкая и мутная. Следует при этом упомянуть, что у кошки, наоборот, симпатическая слюна обычно менее концентрирована, чем хордальная (Ленгли 3). [115]
При долго длящейся секреции и в симпатической слюне постепенно уменьшается содержание органических веществ, причем соответственно с этим изменяется и ее вид, так что она постепенно становится похожей на хордальную (Гейденгайн +). [116]
Следующий опыт Гейденгайна иллюстрирует эти отношения: слюна собиралась в течение 4'/2 часов, первая порция содержала 3.734%, 0/ а последняя -- 1.488% органических веществ.
Секреторное давление симпатичеслюны примерно такое же, как хордальной (Лэнгли 8). [117]
Возбуждение симпатического нерва вызывает в железе ток, противоположный в смысле направления тому, который возникает после раздражения chorda tympani, т. е. в этом случае внешняя поверхность железы ведет себя по отношению к хилусу положительно (Брэдфорд 8). [118]
Секреторное действие симпатического нерва собаки либо вовсе не парализуется атропином, либо парализуется очень большими его дозами (Гейденгайн 1). [119] Из сравнения парализующего действия атропина на хорду и отсутствия или слабого действия его на n. sympathicus заключили, что атропин оказывает влияние не на клетки железы, но на периферические окончания нерва, так как на самый нерв он не действует. Так как между атропином и пилокарпином существует взаимный антагонизм, то из этого далее заключают, что пилокарпин, который сильно возбуждает слюнную секрецию даже после дегенерации хорды, имеет свой пункт приложения также в периферических нервных окончаниях.
Симпатический нерв оказывает на кровеносные сосуды железы суживающее действие, которое при сильных раздражениях может совершенно приостановить кровообращение в железе.
Возбуждение n. sympathicus, как и раздражение хорды, усиливает ток лимфы в железе (Бейнбридж 2). [120]
Симпатические нервные волокна прерываются в верхнем шейном ганглии нервными клетками. Если перерезать n. sympathicus на шее, то через 3-4 дня его верхний конец утрачивает свою раздражимость; при возбуждении нерва по ту сторону ганглия его действие опять ясно выступает. Но после экстирпации верхнего шейного ганглия симпатический нерв торяет раздражимость на всем протяжении (Лэнгли [121]
Перерезка n. sympathicus не влечет за собою ни паралитической секреции, ни атрофии железы, но ведот скорее к гипертрофии последней (Брэдфорд +). [122]
Взаимное действие обоих секреторных нервов проявляется во многом.
Введенное между двумя возбуждениями chorda tympani электрическое раздражение n. sympathicus в значительной мере уменьшает содержание органических веществ при втором раздражении, а также и наоборот (Гейденгайн [123] ). Это подтверждается следующим опытом (табл. 7).
 Если раздражать слабым электрическим током оба нерва одновременно, то выделившееся при этом количество слюны соответствует примерно сумме тех количеств слюны, которые выделяются при возбуждении обоих нервов отдельно. При сильном одновременном электрическом раздражении отделяется меньше слюны, чем при возбуждении одной chorda; n. sympathicus оказывает на хордальную секрецию тормозящее действие (Лэнгли [124] 2).
Наоборот, предшествующее возбуждение хорды в значительной мере усиливает секреторное действие следующего за ним возбуждения n. sympathicus. Это усиливающее действие хорды сохраняется спустя несколько минут после того, как прекращается хордальная секреция. После атропинизации животного хорда утрачивает это свое действие (Лэнгли [125] 3) предшествующего раздражения хорды, при зажатом выводном протоке железы. следующее за этим возбуждение n. sympathicus ведет сначала к явному увеличению объема железы, которое позднее уступает место уменьшению последнего вследствие наступающего сужения сосудов (Бэнч 1). [127] ное слюноотделение (Овсяницкий 2), которое парализуется
Если раздражать слабым электрическим током оба нерва одновременно, то выделившееся при этом количество слюны соответствует примерно сумме тех количеств слюны, которые выделяются при возбуждении обоих нервов отдельно. При сильном одновременном электрическом раздражении отделяется меньше слюны, чем при возбуждении одной chorda; n. sympathicus оказывает на хордальную секрецию тормозящее действие (Лэнгли [124] 2).
Наоборот, предшествующее возбуждение хорды в значительной мере усиливает секреторное действие следующего за ним возбуждения n. sympathicus. Это усиливающее действие хорды сохраняется спустя несколько минут после того, как прекращается хордальная секреция. После атропинизации животного хорда утрачивает это свое действие (Лэнгли [125] 3) предшествующего раздражения хорды, при зажатом выводном протоке железы. следующее за этим возбуждение n. sympathicus ведет сначала к явному увеличению объема железы, которое позднее уступает место уменьшению последнего вследствие наступающего сужения сосудов (Бэнч 1). [127] ное слюноотделение (Овсяницкий 2), которое парализуется
 Если вырезать у собаки подчелюстную железу и затем пропустить через нее ток крови, го начинается оживленное спонтанатропином (Мэтьюс [126] нервов и n. sympathicus к осталь
Отношения черепномозговых поскольку они были исследованы, с только что описанными, так что об этом ничего особого эта железа, с одной стороны, креции. Но длительное его возно точно. Обычно раздражение ным слюнным железам сходны, сообщать не приходится. Здесь следует привести лишь некоторые результаты произведенных на gl. parotis исследований, ибо представляет нечто особенное, а, с другой стороны, в некоторых отношениях исследована особенэлектрическим током n. sympathicus не вызывает никакой себуждение вызывает значительные изменения в микроскопической картине железы. В согласии с этим одновременное (или предшествующее) возбуждение симпатического нерва и черепномозгового нерва ведет к резкому повышению процентного содержанианических веществ в слюне по сравнению с тем, которое наблюдалось при возбуждении одного лишь черепномозгового нерва (Гейденгайн [128] ). Табл. 8 изображает опыт этого автора.
Прекращение кровообращения посредством перевязки кровеносного сосуда не ведет к повышенному содержанию органических веществ, что ясно видно из табл. 9, изображающей опыт того же автора.
С другой стороны, было показано (Лэнгли [129] 2), что и раздражение n. sympathicus у собаки, если ему незадолго перед тем предшествовало раздражение n. Jacobsonii, каждый раз вызывает слюноотделение из gl. parotis, как это наблюдалось и на подчелюстной железе.
Как же теперь истолковать весь переданный здесь в главных чертах опытный материал, касающийся нервов, влияющих на слюнную секрецию? Если принять во внимание все, что недавно или в настоящее время высказано об этом в физиологической литературе, то здесь можно найти очень различные высказывания: с одной стороны, секреторные нервы, т. е. такие, которые непосредственно действуют на железистые клетки, совсем отрицаются и признаются только сосудистые нервы и нервы, влияющие на контрактильные элементы железы (правда, это мнение высказано лишь единично, например Мэтьюсом [130] с другой же стороны, кроме сосудистых нервов, признаются три рода нервных волокон, непосредственно действующих на слюнные железы, а именно: 1) волокна, которые обусловливают выделение жидкости, несущей с собою неорганические вещества (секреторные волокна); 2) волокна, обусловливающие накопление органических веществ в слюне (трофические волокна), и 3) волокна, которые, с одной стороны, вызывают восстановление железы после ее функционального разрушения, а с другой стороны, тормозят секрецию (анаболические, тормозящие волокна).
Если вырезать у собаки подчелюстную железу и затем пропустить через нее ток крови, го начинается оживленное спонтанатропином (Мэтьюс [126] нервов и n. sympathicus к осталь
Отношения черепномозговых поскольку они были исследованы, с только что описанными, так что об этом ничего особого эта железа, с одной стороны, креции. Но длительное его возно точно. Обычно раздражение ным слюнным железам сходны, сообщать не приходится. Здесь следует привести лишь некоторые результаты произведенных на gl. parotis исследований, ибо представляет нечто особенное, а, с другой стороны, в некоторых отношениях исследована особенэлектрическим током n. sympathicus не вызывает никакой себуждение вызывает значительные изменения в микроскопической картине железы. В согласии с этим одновременное (или предшествующее) возбуждение симпатического нерва и черепномозгового нерва ведет к резкому повышению процентного содержанианических веществ в слюне по сравнению с тем, которое наблюдалось при возбуждении одного лишь черепномозгового нерва (Гейденгайн [128] ). Табл. 8 изображает опыт этого автора.
Прекращение кровообращения посредством перевязки кровеносного сосуда не ведет к повышенному содержанию органических веществ, что ясно видно из табл. 9, изображающей опыт того же автора.
С другой стороны, было показано (Лэнгли [129] 2), что и раздражение n. sympathicus у собаки, если ему незадолго перед тем предшествовало раздражение n. Jacobsonii, каждый раз вызывает слюноотделение из gl. parotis, как это наблюдалось и на подчелюстной железе.
Как же теперь истолковать весь переданный здесь в главных чертах опытный материал, касающийся нервов, влияющих на слюнную секрецию? Если принять во внимание все, что недавно или в настоящее время высказано об этом в физиологической литературе, то здесь можно найти очень различные высказывания: с одной стороны, секреторные нервы, т. е. такие, которые непосредственно действуют на железистые клетки, совсем отрицаются и признаются только сосудистые нервы и нервы, влияющие на контрактильные элементы железы (правда, это мнение высказано лишь единично, например Мэтьюсом [130] с другой же стороны, кроме сосудистых нервов, признаются три рода нервных волокон, непосредственно действующих на слюнные железы, а именно: 1) волокна, которые обусловливают выделение жидкости, несущей с собою неорганические вещества (секреторные волокна); 2) волокна, обусловливающие накопление органических веществ в слюне (трофические волокна), и 3) волокна, которые, с одной стороны, вызывают восстановление железы после ее функционального разрушения, а с другой стороны, тормозят секрецию (анаболические, тормозящие волокна).
 Что вообще говорит за то, что существуют особые, отличные от сосудистых нервов, секреторные нервы?
Прежде всего нет никакого теоретического основания для того, чтобы представлять себе нервный аппарат желез особенно простым. Раз мышечная ткань находится под действием специальных, независимых от сосудистых нервов, моторных нервов, почему бы и для железистой ткани не могли бы существовать те же отношения? Отношения слюнных желез, например, к организму и к внешнему миру, как было показано в начале этой главы, во всяком случае не проще, чем те же отношения мышечной ткани. Во-вторых, мы теперь имеем в своем распоряжении точные гистологические данные, доказывающие непосредственную и к тому же многообразную связь нервных волокон с железистыми клетками. В-третьих, в настоящее время констатирован целый ряд случаев, где процессы кровообращения и слюноотделения либо совершенно, либо в значительной мере расходятся. Раздражение chorda tympani на отрезанной собачьей голове вызывает ясную, длящуюся некоторое время слюнную секрецию. Перерезка n. sympathicus, правда, возбуждает кровообращение в железах, не вызывая, однако, ни малейшего слюноотделения, тогда как слабое раздражение chorda tympani, лишь незначительно возбуждающее кровообращение, тотчас же пускает в ход слюнную секрецию. Как упомянуто выше, атропин, парализующий секреторную функцию хорды, абсолютно не нарушает ее сосудорасширяющего действия.
Мэтьюс сомневается в доказательности этого случая несовпадения обеих функций. Спонтанная секреция, наступающая после полного прекращения и последующего за ним восстановления кровообращения, парализуется атропином. Так как остановка кровообращения может длиться от 15 до 30 минут, то Мэтьюс считает себя вправе утверждать, что здесь не может быть речи о нервных элементах и что парализующее действие атропина относится, таким образом, к самой секреторной клетке. Однако допущение полного отмирания всех нервных элементов (нервных волокон, нервных клеток и нервных окончаний) совершенно произвольно и никоим образом не доказано. Как бы ни была желательна и похвальна постоянная критика ходячих физиологических воззрений, как ни тщательно подобрано и заманчиво множество доказательств, выставленных Мэтьюсом в его работе против учения об особых секреторных нервах велез, все же эта работа, как нам кажется, вряд ли может заставить усомниться в правильности этой гипотезы. При сложности физиологического материала можно многое оспаривать или допускать, просто сопоставляя факто из совершенно различных групп явлений. Пользуясь этим же методом, однако с большим правом, ибо дело идет о том же животном и той же области пищеварительных желез, можно указать на факты из физиологии поджелудочной железы. Атропин парализует секреторное действие как n. vagus, так и n. sympathicus на pancreas и в то же время абсолютно не влияет на секреторное действие кислоты с кишечника на ту же железу. Стало быть, атропин парализует фактически какие-то части нервного аппарата, но не самое секретирующую клетку.
Казалось бы, что лучшим доказательством наличия сосудистых волокон различных секреторных нервов мог бы служить факт секреторного действия n. sympathicus; в этом нерве эта функция спарена с сосудосуживающей, что не позволяет рассматривать секреторную работу железы как результат усиленного кровообращения. Особые свойства симпатической слюнной секреции, которая, как известно, очень скудна и останавливается уже обычно во время раздражения, дали повод некоторым физиологам рассматривать n. sympathicus, так сказать, как механический секреторный нерв и толковать его секреторное действие таким образом, что при его раздражении слюна, уже продуцированная без участия этого нерва, выдавливается из железы. Что это не так, что, напротив того, n. sympathicus является настоящим секреторным нервом, выявляется двумя вышеприведенными фактами: долго длящейся (в течение нескольких часов) неудержимой секрецией, которую Гейденгайн получил при раздражении n. sympathicus, и изменением химического состава слюны в течение долго длящегося раздражения n. sympathicus и при включении симпатической секреции между двумя раздражениями chorda tympani. Ясно, что здесь дело идет о самостоятельной секреторной работе, а не о выдавливании старого материала. В том же смысле нужно толковать вышеупомянутые факты (увеличение объема железы при раздражении n. sympathicus и одновременной перевязке выводного протока и усиление тока лимфы при раздражении обоих железистых нервов).
Очень занимал физиологов другой вопрос: что обусловливает разницу в составе слюны, выделенной собакий из одной и той же подчелюстной железы при раздражении chorda tympani и n. sympathicus? Гейденгайн [131] держится взгляда, что все факты, относящиеся к действию нервов на железу, могут быть проще всего объяснены, если признать наличие двух родов секреторных волокон: обусловливающих ток жидкости секреторных волокон и обусловливающих переход органических веществ железы в растворенное состояние трофических волокон. Согласно этому взгляду, в chorda tympani количественный перевес на стороне секреторных волокон, а в n. sympathicus - на стороне трофических; это полностью объясняет разницу в количестве и составе слюны при раздражении того или другого нерва. Это предположение объясняетакже без всякой натяжки случаи отсутствия параллельности между количеством слюны и содержанием солей, с одной стороны, и содержанием органических веществ, с другой стороны, при различных условиях раздражения одного и того же нерва. Особенно ярким был в этом отношении факт, что раздражение n. sympathicus у собаки, которое обычно никогда не дает повода к слюнной секреции из gl. parotis, тем не менее обусловливает значительные изменения в микроскопической картине покоящейся железы и ведет к значительному содержанию органических веществ в слюне, выделенной при следующем затем возбуждении церебрального нерва.
Если раздражать оба нерва одновременно, то выделяется много слюны с высоким содержанием органических веществ.
В духе гейденгайновской гипотезы следует также рассматривать электрические явления в железах. Оба различных процесса сопровождаются двумя идущими в противоположном направлении токами. Один из них соответствует секреции жидкости, другой - выделению органических веществ; один из них совпадает по времени с функцией секреторного нерва, другой - с функцией трофического нерва.
Другое объяснение разницы в действии обоих нервов на железу могло бы состоять лишь в следующим: именно, можно было бы предположить, не зависит ли она от различных комбинаций секреторных волокон с вазомоторными, в хорде - с сосудорасширяющими, в п. sympathicus - с сосудосуживающими волокнами. Это означает, что один раз деятельность железы возбуждается в условиях кровенаполнения, в другой раз - в условиях недостаточного кровообращения. Количество слюны во всяком случае могло бы в известной мере зависеть от этого условия. Как же обстоит дело с химическим составом слюны? Поставленные непосредственно в этом направлении спыты дали, как мы видели выше, повод Гейденгайну отбросить это второе толкование как не соответствующее действительной сущности дела. В подчелюстной железе, так же как в gl. parotis, раздражение черепных нервов одновременно с полным зажатием кровеносных сосудов железы нисколько не повышает содержания органических веществ в слюне по сравнению с возбуждением тех же желез при нормальном кровообращении. В том же смысле свидетельствует тот факт (Ленгли), что у кошек, у которых сосудистые волокна в железистых нервах распределены точь-в-точь так же, как у собаки, при возбуждении n. sympathicus все-таки выделяется менее концентрированная слюна, чем при возбуждении хорды.
Гипотеза Гейденгайна о двух сортах секреторных волокон может быть, таким образом, принята за основу. Но то обстоятельство, что физиологические явления так сложны, постоянно побуждает нас к особенной осторожности в наших заключениях. Бесконечное варьирование опытов, насколько вообще хватает человеческого остроумия, - вот главное правило физиологической работы. И придуманные Ленгли варианты, очевидно, еще раз подтверждают это правило. Во-первых, оказалось, что при слабом отравлении атропином и сильном раздражении хорды не наблюдается даже и следа несовпадения обоих особых действий на железу (повышения процентного содержания органических веществ при одновременном уменьшении количества слюны), что, однако, можно было ожидать, так как у собаки атропин не парализует симпатическую секрецию. А во-вторых, -то самое главное, при возбуждении железы пилокарпином и одновременном кровопускании имеет место резкое повышение процентного содержания органических веществ по сравнению со слюной, выделенной после отравления пилокарпином, но без кровопускания (Ленгли, Флетчер). Этот результат опыта делает то, что для начала гейденгайновское мнение все еще остается гипотезой и что во всяком случае надо считаться с обеими возможностями: может быть, на самом деле существуют два выдвинутых Гейденгайном сорта железистых волокон, а может быть, существует лишь один сорт таких волокон, которые вместе с двумя разными вазомоторными нервами действуют иначе. Новая обработка этого вопроса крайне желательна.
Слюноотделение, имеющее место при нормальном раздражении, как оно представлено на табл. 1, может быть истолковано с точки зрения обеих гипотез. Мы зачастую видим там одну и ту же быстроту секреции при очень различном содержании органических веществ, причем это может быть как угодно часто повторено в одном и том же опыте. Можно принять для случая, где имеется секреция более концентрированной слюны, одновременное сильное раздражение секреторных и трофических волокон Гейденгайна, а для случая, где имеется секреция жидкой слюны, сильное раздражение секреторных и слабое раздражение трофических волокон. Но можно также истолковать эти данные как результат различно сильного раздражения железистого нерва, с одной стороны, и сосудистых нервов - с другой; для секреции концентрированной слюны потребовалась бы тогда комбинация сильного раздражения железистых нервов и слабого раздражения сосудосуживающих нервов, для секреции же жидкой слюны - комбинация слабого раздражения железистого нерва и сильного раздражения сосудорасширяющих нервов. Но тот факт, что секреция может иметь место с одинаковой быстротой, говорит скорее за гейденгайновскую гипотезу, чем за другую; однако Малуазель [132] показал, что при перерезке n. sympathicus и при действии съедобных веществ слюна остается концентрированной даже больше, чем до перерезки нерва. Дальнейшим материалом для разрешения того же вопроса могут служить следующие установленные Малуазелем факты: при отравлении атропином собаки со слюнной фистулой как на пищевые вещества (мясо), так и на отвергаемые вещества (соль) изливается несколько капель густой слюны и в последнее случае в меньшем количестве, чем в первом, хотя в нормальных условиях дело обстоит как раз наоборот. Те же явления наблюдаются на собаке с регенерирующей после перерезки хордой.
Вопрос о центробежных железистых нервах не ограничивается, однако, вышеупомянутыми пунктами. Некоторые факты заставили предположить наличие особых волокон, тормозящих деятельность желез. Чермак 2 [133] 2 первый высказался за наличие таких волокон в n. sympathicus, а именно по той причине, что одновременное или предшествующее сильное раздражение n. sympathicus либо значительно ослабляет, либо совершенно прекращает секреторное действие хорды. Более поздние авторы, правда, признали только что упомянутый факт, но объясняли его или косвенным действием n. sympathicus, или тем, что концентрированная, вязкая симпатическая слюна затруднена в своем продвижении по узким выводным протокам (Эккард [134] или параличом сецернирующих клеток вследствие недостатка кислорода (Тейденгайн 2 [135] 2), или, наконец, вообще уменьшением кровенаполнения железы (Ленгли 3 [136] 9). Так как при слабом раздражении обоих нервов наблюдается даже суммирование их секреторных эффектов и, кроме того, при угнетении кровообращения, вследствие сужения просвета сосудов, также наблюдается уменьшение секреторного действия хорды, то в этом пункте не кроется никакого основания для признания наличия тормозящих волоков. Однако стали известны другие факты, говорящие за это предположение. Брэдфорд [137] 4 указал на то, что у кошки раздражение n. sympathicus влечет за собою секрецию жидкой слюны, но что перерезка этого нерва не ведет к атрофии этой железы, как перерезка хорды. Стоя на точке зрения гейденгайновского учения, которое гласит, что существуют секреторные волокна двух родов и что вследствие этого в n. sympathicus кошки имеются в достаточном количестве как секреторные, так и трофические волокна, Брэдфорд думал, что можно объяснить разницу результатов перерезки обоих нервов в отношении к последующему состоянию железы тем, что он допустил наличие в хорде особых, третьего рода волокон - анаболических, волокон, обусловливающих восстановление секреторных клеток после их функционального разрушения. Эти же волокна должны, по его мнению, действовать тормозящим образом на секрецию, так как этот процесс является, ведь, противоположным восстановительному процессу. В соответствии с этим он, естественно, рассматривает паралитическую секрецию как результат перерезки этих волокон. Что касается первого предположения, то оно не может претендовать на достаточную доказательность, а именно, во-первых, потому, что оно построено на геиденгайновской гипотезе о двух сортах секреторных волокон, которая еще сама должна быть сначала доказана, и, во-вторых, потому, что упомянутая разница результатов перерезки нервов гораздо проще может быть объяснена таким образом, что после перерезки хорды железа перестает работать; рефлекс на секрецию через оставшийся n. sympathicus не наблюдался до сих пор ни у собаки, ни у кошки (Гейденгайн 1, [138] Павлов 2). 2 [139] Но бездеятельность железы должна естественно вести к ее атрофии. Что касается второго предположения Брэдфорда, 3 [140] то оно заслуживает большего внимания, так как затрагивает вопрос о причинах паралитической секреции. Из всех объяснений этой секреции фактическому положению дела более всего соответствует предположение, что она обусловливается возбуждением местного нервного механизма, исходящим из периферических нервных клеток (Лэнгли [141] *). Если это так, то до известной степени кажется вероятным, что при нормальных отношениях это раздражение тормозится из центральной нервной системы через посредство специального нерва. В том же смысле Овсяницкий [142] истолковывает тот факт, что вырезанная железа при возобновлении кровообращения после временного его прекращения начинает спонтанно секретировать.
Стало быть, из всего нервного аппарата слюнных желез физиология выделила, хотя далеко не достаточно проанализировала, простейшую часть этого аппарата, крейнее его звено, которое непосредственно касается железы и которое при искусственном раздражении в различных условиях вызывает все колебания в функции желез, которые могут наблюдаться при их нормальной деятельности.
Что вообще говорит за то, что существуют особые, отличные от сосудистых нервов, секреторные нервы?
Прежде всего нет никакого теоретического основания для того, чтобы представлять себе нервный аппарат желез особенно простым. Раз мышечная ткань находится под действием специальных, независимых от сосудистых нервов, моторных нервов, почему бы и для железистой ткани не могли бы существовать те же отношения? Отношения слюнных желез, например, к организму и к внешнему миру, как было показано в начале этой главы, во всяком случае не проще, чем те же отношения мышечной ткани. Во-вторых, мы теперь имеем в своем распоряжении точные гистологические данные, доказывающие непосредственную и к тому же многообразную связь нервных волокон с железистыми клетками. В-третьих, в настоящее время констатирован целый ряд случаев, где процессы кровообращения и слюноотделения либо совершенно, либо в значительной мере расходятся. Раздражение chorda tympani на отрезанной собачьей голове вызывает ясную, длящуюся некоторое время слюнную секрецию. Перерезка n. sympathicus, правда, возбуждает кровообращение в железах, не вызывая, однако, ни малейшего слюноотделения, тогда как слабое раздражение chorda tympani, лишь незначительно возбуждающее кровообращение, тотчас же пускает в ход слюнную секрецию. Как упомянуто выше, атропин, парализующий секреторную функцию хорды, абсолютно не нарушает ее сосудорасширяющего действия.
Мэтьюс сомневается в доказательности этого случая несовпадения обеих функций. Спонтанная секреция, наступающая после полного прекращения и последующего за ним восстановления кровообращения, парализуется атропином. Так как остановка кровообращения может длиться от 15 до 30 минут, то Мэтьюс считает себя вправе утверждать, что здесь не может быть речи о нервных элементах и что парализующее действие атропина относится, таким образом, к самой секреторной клетке. Однако допущение полного отмирания всех нервных элементов (нервных волокон, нервных клеток и нервных окончаний) совершенно произвольно и никоим образом не доказано. Как бы ни была желательна и похвальна постоянная критика ходячих физиологических воззрений, как ни тщательно подобрано и заманчиво множество доказательств, выставленных Мэтьюсом в его работе против учения об особых секреторных нервах велез, все же эта работа, как нам кажется, вряд ли может заставить усомниться в правильности этой гипотезы. При сложности физиологического материала можно многое оспаривать или допускать, просто сопоставляя факто из совершенно различных групп явлений. Пользуясь этим же методом, однако с большим правом, ибо дело идет о том же животном и той же области пищеварительных желез, можно указать на факты из физиологии поджелудочной железы. Атропин парализует секреторное действие как n. vagus, так и n. sympathicus на pancreas и в то же время абсолютно не влияет на секреторное действие кислоты с кишечника на ту же железу. Стало быть, атропин парализует фактически какие-то части нервного аппарата, но не самое секретирующую клетку.
Казалось бы, что лучшим доказательством наличия сосудистых волокон различных секреторных нервов мог бы служить факт секреторного действия n. sympathicus; в этом нерве эта функция спарена с сосудосуживающей, что не позволяет рассматривать секреторную работу железы как результат усиленного кровообращения. Особые свойства симпатической слюнной секреции, которая, как известно, очень скудна и останавливается уже обычно во время раздражения, дали повод некоторым физиологам рассматривать n. sympathicus, так сказать, как механический секреторный нерв и толковать его секреторное действие таким образом, что при его раздражении слюна, уже продуцированная без участия этого нерва, выдавливается из железы. Что это не так, что, напротив того, n. sympathicus является настоящим секреторным нервом, выявляется двумя вышеприведенными фактами: долго длящейся (в течение нескольких часов) неудержимой секрецией, которую Гейденгайн получил при раздражении n. sympathicus, и изменением химического состава слюны в течение долго длящегося раздражения n. sympathicus и при включении симпатической секреции между двумя раздражениями chorda tympani. Ясно, что здесь дело идет о самостоятельной секреторной работе, а не о выдавливании старого материала. В том же смысле нужно толковать вышеупомянутые факты (увеличение объема железы при раздражении n. sympathicus и одновременной перевязке выводного протока и усиление тока лимфы при раздражении обоих железистых нервов).
Очень занимал физиологов другой вопрос: что обусловливает разницу в составе слюны, выделенной собакий из одной и той же подчелюстной железы при раздражении chorda tympani и n. sympathicus? Гейденгайн [131] держится взгляда, что все факты, относящиеся к действию нервов на железу, могут быть проще всего объяснены, если признать наличие двух родов секреторных волокон: обусловливающих ток жидкости секреторных волокон и обусловливающих переход органических веществ железы в растворенное состояние трофических волокон. Согласно этому взгляду, в chorda tympani количественный перевес на стороне секреторных волокон, а в n. sympathicus - на стороне трофических; это полностью объясняет разницу в количестве и составе слюны при раздражении того или другого нерва. Это предположение объясняетакже без всякой натяжки случаи отсутствия параллельности между количеством слюны и содержанием солей, с одной стороны, и содержанием органических веществ, с другой стороны, при различных условиях раздражения одного и того же нерва. Особенно ярким был в этом отношении факт, что раздражение n. sympathicus у собаки, которое обычно никогда не дает повода к слюнной секреции из gl. parotis, тем не менее обусловливает значительные изменения в микроскопической картине покоящейся железы и ведет к значительному содержанию органических веществ в слюне, выделенной при следующем затем возбуждении церебрального нерва.
Если раздражать оба нерва одновременно, то выделяется много слюны с высоким содержанием органических веществ.
В духе гейденгайновской гипотезы следует также рассматривать электрические явления в железах. Оба различных процесса сопровождаются двумя идущими в противоположном направлении токами. Один из них соответствует секреции жидкости, другой - выделению органических веществ; один из них совпадает по времени с функцией секреторного нерва, другой - с функцией трофического нерва.
Другое объяснение разницы в действии обоих нервов на железу могло бы состоять лишь в следующим: именно, можно было бы предположить, не зависит ли она от различных комбинаций секреторных волокон с вазомоторными, в хорде - с сосудорасширяющими, в п. sympathicus - с сосудосуживающими волокнами. Это означает, что один раз деятельность железы возбуждается в условиях кровенаполнения, в другой раз - в условиях недостаточного кровообращения. Количество слюны во всяком случае могло бы в известной мере зависеть от этого условия. Как же обстоит дело с химическим составом слюны? Поставленные непосредственно в этом направлении спыты дали, как мы видели выше, повод Гейденгайну отбросить это второе толкование как не соответствующее действительной сущности дела. В подчелюстной железе, так же как в gl. parotis, раздражение черепных нервов одновременно с полным зажатием кровеносных сосудов железы нисколько не повышает содержания органических веществ в слюне по сравнению с возбуждением тех же желез при нормальном кровообращении. В том же смысле свидетельствует тот факт (Ленгли), что у кошек, у которых сосудистые волокна в железистых нервах распределены точь-в-точь так же, как у собаки, при возбуждении n. sympathicus все-таки выделяется менее концентрированная слюна, чем при возбуждении хорды.
Гипотеза Гейденгайна о двух сортах секреторных волокон может быть, таким образом, принята за основу. Но то обстоятельство, что физиологические явления так сложны, постоянно побуждает нас к особенной осторожности в наших заключениях. Бесконечное варьирование опытов, насколько вообще хватает человеческого остроумия, - вот главное правило физиологической работы. И придуманные Ленгли варианты, очевидно, еще раз подтверждают это правило. Во-первых, оказалось, что при слабом отравлении атропином и сильном раздражении хорды не наблюдается даже и следа несовпадения обоих особых действий на железу (повышения процентного содержания органических веществ при одновременном уменьшении количества слюны), что, однако, можно было ожидать, так как у собаки атропин не парализует симпатическую секрецию. А во-вторых, -то самое главное, при возбуждении железы пилокарпином и одновременном кровопускании имеет место резкое повышение процентного содержания органических веществ по сравнению со слюной, выделенной после отравления пилокарпином, но без кровопускания (Ленгли, Флетчер). Этот результат опыта делает то, что для начала гейденгайновское мнение все еще остается гипотезой и что во всяком случае надо считаться с обеими возможностями: может быть, на самом деле существуют два выдвинутых Гейденгайном сорта железистых волокон, а может быть, существует лишь один сорт таких волокон, которые вместе с двумя разными вазомоторными нервами действуют иначе. Новая обработка этого вопроса крайне желательна.
Слюноотделение, имеющее место при нормальном раздражении, как оно представлено на табл. 1, может быть истолковано с точки зрения обеих гипотез. Мы зачастую видим там одну и ту же быстроту секреции при очень различном содержании органических веществ, причем это может быть как угодно часто повторено в одном и том же опыте. Можно принять для случая, где имеется секреция более концентрированной слюны, одновременное сильное раздражение секреторных и трофических волокон Гейденгайна, а для случая, где имеется секреция жидкой слюны, сильное раздражение секреторных и слабое раздражение трофических волокон. Но можно также истолковать эти данные как результат различно сильного раздражения железистого нерва, с одной стороны, и сосудистых нервов - с другой; для секреции концентрированной слюны потребовалась бы тогда комбинация сильного раздражения железистых нервов и слабого раздражения сосудосуживающих нервов, для секреции же жидкой слюны - комбинация слабого раздражения железистого нерва и сильного раздражения сосудорасширяющих нервов. Но тот факт, что секреция может иметь место с одинаковой быстротой, говорит скорее за гейденгайновскую гипотезу, чем за другую; однако Малуазель [132] показал, что при перерезке n. sympathicus и при действии съедобных веществ слюна остается концентрированной даже больше, чем до перерезки нерва. Дальнейшим материалом для разрешения того же вопроса могут служить следующие установленные Малуазелем факты: при отравлении атропином собаки со слюнной фистулой как на пищевые вещества (мясо), так и на отвергаемые вещества (соль) изливается несколько капель густой слюны и в последнее случае в меньшем количестве, чем в первом, хотя в нормальных условиях дело обстоит как раз наоборот. Те же явления наблюдаются на собаке с регенерирующей после перерезки хордой.
Вопрос о центробежных железистых нервах не ограничивается, однако, вышеупомянутыми пунктами. Некоторые факты заставили предположить наличие особых волокон, тормозящих деятельность желез. Чермак 2 [133] 2 первый высказался за наличие таких волокон в n. sympathicus, а именно по той причине, что одновременное или предшествующее сильное раздражение n. sympathicus либо значительно ослабляет, либо совершенно прекращает секреторное действие хорды. Более поздние авторы, правда, признали только что упомянутый факт, но объясняли его или косвенным действием n. sympathicus, или тем, что концентрированная, вязкая симпатическая слюна затруднена в своем продвижении по узким выводным протокам (Эккард [134] или параличом сецернирующих клеток вследствие недостатка кислорода (Тейденгайн 2 [135] 2), или, наконец, вообще уменьшением кровенаполнения железы (Ленгли 3 [136] 9). Так как при слабом раздражении обоих нервов наблюдается даже суммирование их секреторных эффектов и, кроме того, при угнетении кровообращения, вследствие сужения просвета сосудов, также наблюдается уменьшение секреторного действия хорды, то в этом пункте не кроется никакого основания для признания наличия тормозящих волоков. Однако стали известны другие факты, говорящие за это предположение. Брэдфорд [137] 4 указал на то, что у кошки раздражение n. sympathicus влечет за собою секрецию жидкой слюны, но что перерезка этого нерва не ведет к атрофии этой железы, как перерезка хорды. Стоя на точке зрения гейденгайновского учения, которое гласит, что существуют секреторные волокна двух родов и что вследствие этого в n. sympathicus кошки имеются в достаточном количестве как секреторные, так и трофические волокна, Брэдфорд думал, что можно объяснить разницу результатов перерезки обоих нервов в отношении к последующему состоянию железы тем, что он допустил наличие в хорде особых, третьего рода волокон - анаболических, волокон, обусловливающих восстановление секреторных клеток после их функционального разрушения. Эти же волокна должны, по его мнению, действовать тормозящим образом на секрецию, так как этот процесс является, ведь, противоположным восстановительному процессу. В соответствии с этим он, естественно, рассматривает паралитическую секрецию как результат перерезки этих волокон. Что касается первого предположения, то оно не может претендовать на достаточную доказательность, а именно, во-первых, потому, что оно построено на геиденгайновской гипотезе о двух сортах секреторных волокон, которая еще сама должна быть сначала доказана, и, во-вторых, потому, что упомянутая разница результатов перерезки нервов гораздо проще может быть объяснена таким образом, что после перерезки хорды железа перестает работать; рефлекс на секрецию через оставшийся n. sympathicus не наблюдался до сих пор ни у собаки, ни у кошки (Гейденгайн 1, [138] Павлов 2). 2 [139] Но бездеятельность железы должна естественно вести к ее атрофии. Что касается второго предположения Брэдфорда, 3 [140] то оно заслуживает большего внимания, так как затрагивает вопрос о причинах паралитической секреции. Из всех объяснений этой секреции фактическому положению дела более всего соответствует предположение, что она обусловливается возбуждением местного нервного механизма, исходящим из периферических нервных клеток (Лэнгли [141] *). Если это так, то до известной степени кажется вероятным, что при нормальных отношениях это раздражение тормозится из центральной нервной системы через посредство специального нерва. В том же смысле Овсяницкий [142] истолковывает тот факт, что вырезанная железа при возобновлении кровообращения после временного его прекращения начинает спонтанно секретировать.
Стало быть, из всего нервного аппарата слюнных желез физиология выделила, хотя далеко не достаточно проанализировала, простейшую часть этого аппарата, крейнее его звено, которое непосредственно касается железы и которое при искусственном раздражении в различных условиях вызывает все колебания в функции желез, которые могут наблюдаться при их нормальной деятельности.
 Подобное же исследование всего механизма действия других раздражителей (а именно съедобных веществ), которое поставило себе задачей решить, какие элементарные свойства данного объекта оказывают влияние и на какой периферический концевой аппарат они падают, представляет одну из ближайших задач научного исследования. Эта задача никак не может быть отнесена к легким. В острых опытах с отравлениями и свежими повреждениями центральной нервной системы приходится бороться с побочными влияниями этих грубых вмешательств, и едва ли возможно признать, что их можно полностью устранить.
В хронических опытах, поставленных на нормальных животных, сложнонервные, психические отношения выявляются в полной мере. Очевидно, придетстараться раздобывать животных, совершенно оправившихся после удаления верхних отделов центральной нервной системы.
Указывалось на рефлекторное раздражение слюнных желез с желудка, но более точные исследования последних из вышеупомянутых авторов не смогли подтвердить этих данных. Очень распространен взгляд, что слюнная секреция возбуждается рефлекторно со слизистой носа, через посредство обонятельного нерва. Снарскому [151] в его опытах на кураризованных животных удавалось вызывать слюнную секрецию с носа только местно сильно раздражающими газами и парами, например аммиаком, горчичным маслом и т. п. Кроме того, этот рефлекс наблюдался лишь при неповрежденном n. trigeminus; после его перерезки никакие запахи и газы не могли больше вызвать секрецию.
Подобное же исследование всего механизма действия других раздражителей (а именно съедобных веществ), которое поставило себе задачей решить, какие элементарные свойства данного объекта оказывают влияние и на какой периферический концевой аппарат они падают, представляет одну из ближайших задач научного исследования. Эта задача никак не может быть отнесена к легким. В острых опытах с отравлениями и свежими повреждениями центральной нервной системы приходится бороться с побочными влияниями этих грубых вмешательств, и едва ли возможно признать, что их можно полностью устранить.
В хронических опытах, поставленных на нормальных животных, сложнонервные, психические отношения выявляются в полной мере. Очевидно, придетстараться раздобывать животных, совершенно оправившихся после удаления верхних отделов центральной нервной системы.
Указывалось на рефлекторное раздражение слюнных желез с желудка, но более точные исследования последних из вышеупомянутых авторов не смогли подтвердить этих данных. Очень распространен взгляд, что слюнная секреция возбуждается рефлекторно со слизистой носа, через посредство обонятельного нерва. Снарскому [151] в его опытах на кураризованных животных удавалось вызывать слюнную секрецию с носа только местно сильно раздражающими газами и парами, например аммиаком, горчичным маслом и т. п. Кроме того, этот рефлекс наблюдался лишь при неповрежденном n. trigeminus; после его перерезки никакие запахи и газы не могли больше вызвать секрецию.
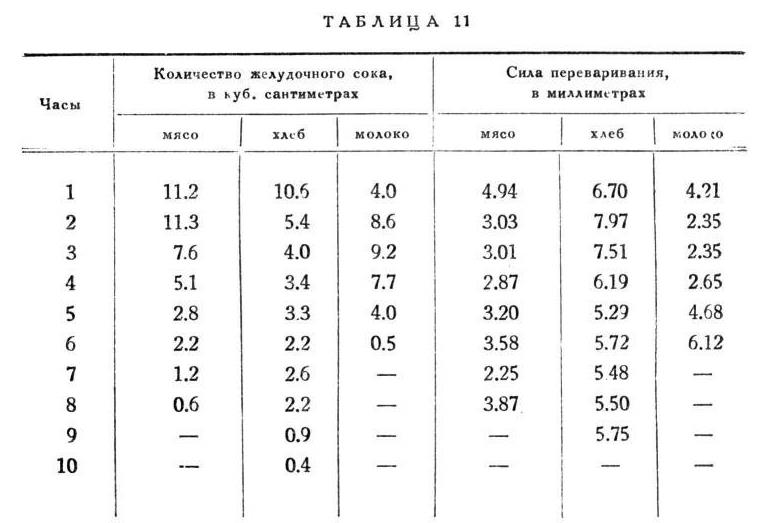 Кислотность желудочного сока соответствует в своих колебаниях быстроте секреции, т. е. количеству сока, выделившегося за единицу времени; чем значительнее быстрота, тем выше кислотность, и наоборот.
Количество желудочного сока, изливающегося в течение всего секреторного периода на одинаковые по весу количества трех упомянутых сортов пищи, наиболее значительно для мяса; оно меньше для хлеба и меньше всего для молока. На эквивалентные по содержанию азота (т. е. примерно по содержанию белковых веществ) количества трех сортов пищи при кормлении хлебом изливается больше всего, при кормлении молоком меньше, а при кормлении мясом - меньше всего желудочного сока.
Очень резко выступает разница в переваривающей силе различных сортов сока. Наиболее концентрированным является хлебный сок, слабейшим - молочный сок; мясной сок занимает среднее положение. Это один из наиболее выдающихся фактов, характеризующих своеобразие работы пепсиновых желез в зависимости от сортапищи. В то же время, как будет видно из дальнейшего, это является неопровержимым фактом (несмотря на то, что количественное определение ферментов вообще очень трудно). Различный состав сортов желудочного сока одинаково отчетливо выступает при исследовании их по различным методикам.
Ввиду важности дела мы его здесь рассмотрим подробнее. Приведенная ниже таблица (12) содержит числовые величины переваривающей силы, взятые из сообщения доктора Керстена [181] и полученные на одной и той же собаке.
Кислотность желудочного сока соответствует в своих колебаниях быстроте секреции, т. е. количеству сока, выделившегося за единицу времени; чем значительнее быстрота, тем выше кислотность, и наоборот.
Количество желудочного сока, изливающегося в течение всего секреторного периода на одинаковые по весу количества трех упомянутых сортов пищи, наиболее значительно для мяса; оно меньше для хлеба и меньше всего для молока. На эквивалентные по содержанию азота (т. е. примерно по содержанию белковых веществ) количества трех сортов пищи при кормлении хлебом изливается больше всего, при кормлении молоком меньше, а при кормлении мясом - меньше всего желудочного сока.
Очень резко выступает разница в переваривающей силе различных сортов сока. Наиболее концентрированным является хлебный сок, слабейшим - молочный сок; мясной сок занимает среднее положение. Это один из наиболее выдающихся фактов, характеризующих своеобразие работы пепсиновых желез в зависимости от сортапищи. В то же время, как будет видно из дальнейшего, это является неопровержимым фактом (несмотря на то, что количественное определение ферментов вообще очень трудно). Различный состав сортов желудочного сока одинаково отчетливо выступает при исследовании их по различным методикам.
Ввиду важности дела мы его здесь рассмотрим подробнее. Приведенная ниже таблица (12) содержит числовые величины переваривающей силы, взятые из сообщения доктора Керстена [181] и полученные на одной и той же собаке.
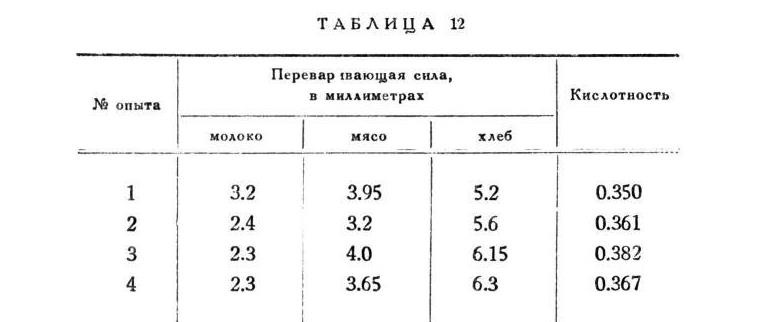 Как видно из табл. 12, только в каждом горизонтальном ряду порции выравнены в отношении кислотности и разведения.
Различные сорта сока сравниваются также на сухой остаток после выпаривания. В нижеприведенной таблице (13), заимствованной тоже у Керстена, приведены числа для молочного много сока, которые больше всего отличаются друг от друга.
Как видно из табл. 12, только в каждом горизонтальном ряду порции выравнены в отношении кислотности и разведения.
Различные сорта сока сравниваются также на сухой остаток после выпаривания. В нижеприведенной таблице (13), заимствованной тоже у Керстена, приведены числа для молочного много сока, которые больше всего отличаются друг от друга.
 Осадки, образующиеся при добывании алкоголя, оказались для различных сортов сока также различными, и именно в том смысле, как это явствует из нижеприводимой таблицы (14).
Осадки, образующиеся при добывании алкоголя, оказались для различных сортов сока также различными, и именно в том смысле, как это явствует из нижеприводимой таблицы (14).
 Совершенно то же самое получается и для осадков, образующихся при кипячении желудочного сока, как доказывает следующая таблица (15) (из сообщения Керстена).
В обоих последних случаях количества осадков вели себя часто как квадраты миллиметровых чисел, представляющих переваривающую силу.
Принимая во внимание только что приведенные числа и соображения, изложенные в методической части, можно совершенно точным образом высчитать количества ферментов, выделяемых пепсиновыми железами на одинаковое количество белка, заключающегося в различныx сортах пищи. Следующая таблица (16) дает соответственные цифры (по Хижину).
Совершенно то же самое получается и для осадков, образующихся при кипячении желудочного сока, как доказывает следующая таблица (15) (из сообщения Керстена).
В обоих последних случаях количества осадков вели себя часто как квадраты миллиметровых чисел, представляющих переваривающую силу.
Принимая во внимание только что приведенные числа и соображения, изложенные в методической части, можно совершенно точным образом высчитать количества ферментов, выделяемых пепсиновыми железами на одинаковое количество белка, заключающегося в различныx сортах пищи. Следующая таблица (16) дает соответственные цифры (по Хижину).
 Если мы возьмем квадраты миллиметровых чисел, то получим 38 для хлеба, 16 для мяса и 10 для молока. Эти последние числа соответствуют относительным количеством ферментов различных сортов сока в единицу объема. Перемиожив эти числа на количество кубических сантиметров сока, мы получим 1600 для хлеба, 430 для мяса и 340 для молока. Эти последние числа представляют, стало быть, относительное количество фермента, изливающееся на одинаковое количество белка, принятого животным с различными сортами пищи.
Что касается кислотности, то здесь последовательность сортов сока другая: здесь во главе стоит сок, изливающийся на мясо, наименьшую кислотность показывает сок, изливающийся на хлеб, среднее положение занимает молочный сок.
Вследствие несовершенства своей методики клиническое исследование, очевидно, не могло до сих пор подтвердить те же отношения на человеке. Хотя некоторые авторы смогли констатировать различное поведение работы желудочных желез в зависимости от сорта пищи, клиницистами не выставлено еще в этом отношении никаких общепринятых положений.
Конечно, очень важным является вопрос: какое значение имеют колебания, отмеченные в работе пепсиновых желез при кормлении собаки различными соругами пищи? Уже a priori нельзя допустить, что эти колебания случайные и что они не стоят ни в какой связи со свойствами съеденной пищи. К сожалению, мы сейчас не располагаем каким бы то ни было материалом, чтобы ответить на этот вопрос строго научно. Но уже в настоящее время нельзя не заметить некоторых целесообразных отношений между характером пищи и желудочной секрецией.
Для белка молока пепсиновые железы поставляют самое малое количество фермента и к тому же в слабейшей концентрации. С этим согласуется известный физиологической химии факт, что наряду с фибрином казеин принадлежит к наиболее легко перевариваемым сортам белка. На хлебный белок изливается сравнительно очень обильный фермант и в очень значительной концентрации. Этому со своей стороны соответствует тот факт, что растительные сорта белков принадлежат к наиболее трудно одолимым ферментами. Мясной белок занимает во всех отношениях, т. е. что касается количества и концентрации фермента, среднее место.
При одинаковых весовых количествах хлеба и мяса на первый выделяется гораздо больше фермента, чем на последний, в то же время оказывается, что если принять во внимание количество и кислотность сока, то на мясо изливается гораздо больше соляной кислоты, чем на хлеб. Это становится понятным, если принять в соображение, что для мяса требуется большее количество кислоты, чтобы скорее растворить содержащуюся в нем соединительную ткань. Для хлеба же избыток кислоты был бы не только не нужен, но мог бы даже повредить перевариванию крахмала как в желудке, так и в дальнейших отрезках желудочно-кишечного канала.
При кормлении мясом и хлебом, там, где принятая пища должна быть тотчас же обработана, за первые часы секреторного периода изливается обильнейшее количество желудочного сока. При кормлении молоком дело происходит совершенно иначе. Попавшее в желудок молоко начинает свертываться уже через несколько минут, а после этого в течение значительного промежутка времени жидкие составные части молока переходят в кишечник; к этому времени секреция желудочного сока очень вялая; только к концу второго часа, а именно в течение третьего часа, когда остаются только хлопья казеина, изливается наибольшее количество желудочного сока; если бы ход секреции был другой, то потребовалось бы гораздо большее количество желудочного сока, чтобы переварить то же количество казеина.
Если мы возьмем квадраты миллиметровых чисел, то получим 38 для хлеба, 16 для мяса и 10 для молока. Эти последние числа соответствуют относительным количеством ферментов различных сортов сока в единицу объема. Перемиожив эти числа на количество кубических сантиметров сока, мы получим 1600 для хлеба, 430 для мяса и 340 для молока. Эти последние числа представляют, стало быть, относительное количество фермента, изливающееся на одинаковое количество белка, принятого животным с различными сортами пищи.
Что касается кислотности, то здесь последовательность сортов сока другая: здесь во главе стоит сок, изливающийся на мясо, наименьшую кислотность показывает сок, изливающийся на хлеб, среднее положение занимает молочный сок.
Вследствие несовершенства своей методики клиническое исследование, очевидно, не могло до сих пор подтвердить те же отношения на человеке. Хотя некоторые авторы смогли констатировать различное поведение работы желудочных желез в зависимости от сорта пищи, клиницистами не выставлено еще в этом отношении никаких общепринятых положений.
Конечно, очень важным является вопрос: какое значение имеют колебания, отмеченные в работе пепсиновых желез при кормлении собаки различными соругами пищи? Уже a priori нельзя допустить, что эти колебания случайные и что они не стоят ни в какой связи со свойствами съеденной пищи. К сожалению, мы сейчас не располагаем каким бы то ни было материалом, чтобы ответить на этот вопрос строго научно. Но уже в настоящее время нельзя не заметить некоторых целесообразных отношений между характером пищи и желудочной секрецией.
Для белка молока пепсиновые железы поставляют самое малое количество фермента и к тому же в слабейшей концентрации. С этим согласуется известный физиологической химии факт, что наряду с фибрином казеин принадлежит к наиболее легко перевариваемым сортам белка. На хлебный белок изливается сравнительно очень обильный фермант и в очень значительной концентрации. Этому со своей стороны соответствует тот факт, что растительные сорта белков принадлежат к наиболее трудно одолимым ферментами. Мясной белок занимает во всех отношениях, т. е. что касается количества и концентрации фермента, среднее место.
При одинаковых весовых количествах хлеба и мяса на первый выделяется гораздо больше фермента, чем на последний, в то же время оказывается, что если принять во внимание количество и кислотность сока, то на мясо изливается гораздо больше соляной кислоты, чем на хлеб. Это становится понятным, если принять в соображение, что для мяса требуется большее количество кислоты, чтобы скорее растворить содержащуюся в нем соединительную ткань. Для хлеба же избыток кислоты был бы не только не нужен, но мог бы даже повредить перевариванию крахмала как в желудке, так и в дальнейших отрезках желудочно-кишечного канала.
При кормлении мясом и хлебом, там, где принятая пища должна быть тотчас же обработана, за первые часы секреторного периода изливается обильнейшее количество желудочного сока. При кормлении молоком дело происходит совершенно иначе. Попавшее в желудок молоко начинает свертываться уже через несколько минут, а после этого в течение значительного промежутка времени жидкие составные части молока переходят в кишечник; к этому времени секреция желудочного сока очень вялая; только к концу второго часа, а именно в течение третьего часа, когда остаются только хлопья казеина, изливается наибольшее количество желудочного сока; если бы ход секреции был другой, то потребовалось бы гораздо большее количество желудочного сока, чтобы переварить то же количество казеина.
 Только через 1 час 10 минут слизь показала кислую реакцию.
Что касается возбуждающего секрецию желудочного сока действия воды и растворов экстрактивных веществ, вводимых непосредственно в желудок, но при свободной проходимости кишечника, то и это явление наблюдалось многочисленными авторами. Только о сыром яичном белке утверждалось (Хижин, Лобасов), что с желудка он действует не как химический раздражитель. Это иное поведение легко объясняется тем, что при свободном прохождении через pylorus (как и было у этих авторов) белок скоро покидает желудок. Как же теперь толковать медленное действие жидкого яичного белка? Он действует либо как очень слабый раздражитель, который может дать себя почувствовать только после долгого суммационного времени, или он непосредственно вообще не действует. В последнем случае можно было бы предположить, что сперва содержащаяся в белке вода вызывает очень скудное сокоотделение и что потом выделившийся при этом сок образует из белка продукты, действующие как главный раздражитель.
Что последнее предположение соответствует действительному положению вещей, за это говорит, между прочим, тот факт, что продукты переваривания белковых веществ возбуждают деятельность пепсиновых желез, факт, который ясно выступает в опытах Соколова. Этот же факт подтверждается также другими авторами.
В полнейшем согласии с этим находится следующий факт: хорошо вываренное мясо, из которого выжать вся вода, введенное непосредственно в желудок, не возбуждает пепсиновых желез. Так как в этом случае из мяса удалено все растворимое в воде и действующее химически, то это последнее действует теперь только как механический раздражитель и поэтому не может возбуждать деятельность пепсиновых желез. Вполне понятно, что непосредственно введенные в желудочную полость кусочки вареного. яичного белка совершенно не влияют на железы (Шифф [189] Лобасов и др.).
Все упомянутые вещества (мясо, вода, растворы экстрактовных веществ, продукты переваривания белковых веществ), если они непосредственно введены в кишечник, причем, конечно, должно быть исключено какое бы то ни было психическое воздействие, или вовсе не возбуждают пепсиновые железы, или в значительно более слабой степени, чем с желудка. (Тот факт, что вообще пепсиновые железы могут быть возбуждены с кишечника, наблюдался мною, 2 [190] 2 позднее Леконтом [191] 9). Подобные опыты могут быть проведены совершенно точным образом на собаках, оперированных по Соколову. Опыты над собаками с боковыми кишечными фистулами, без отделения желудка от кишечника, не могут разрешить этого вопроса удовлетворительным образом, ибо часть кишечного содержимого всегда может быть заброшена обратно, что очень легко происходит при известных условиях.
Мы воспроизводим здесь сравнительный опыт (табл. 18) с мясным фаршем; в этом виде мясо легче всего может быть введено в различные полости пищеварительного канала, без того, чтобы животное что-либо заметило (Соколов).
Только через 1 час 10 минут слизь показала кислую реакцию.
Что касается возбуждающего секрецию желудочного сока действия воды и растворов экстрактивных веществ, вводимых непосредственно в желудок, но при свободной проходимости кишечника, то и это явление наблюдалось многочисленными авторами. Только о сыром яичном белке утверждалось (Хижин, Лобасов), что с желудка он действует не как химический раздражитель. Это иное поведение легко объясняется тем, что при свободном прохождении через pylorus (как и было у этих авторов) белок скоро покидает желудок. Как же теперь толковать медленное действие жидкого яичного белка? Он действует либо как очень слабый раздражитель, который может дать себя почувствовать только после долгого суммационного времени, или он непосредственно вообще не действует. В последнем случае можно было бы предположить, что сперва содержащаяся в белке вода вызывает очень скудное сокоотделение и что потом выделившийся при этом сок образует из белка продукты, действующие как главный раздражитель.
Что последнее предположение соответствует действительному положению вещей, за это говорит, между прочим, тот факт, что продукты переваривания белковых веществ возбуждают деятельность пепсиновых желез, факт, который ясно выступает в опытах Соколова. Этот же факт подтверждается также другими авторами.
В полнейшем согласии с этим находится следующий факт: хорошо вываренное мясо, из которого выжать вся вода, введенное непосредственно в желудок, не возбуждает пепсиновых желез. Так как в этом случае из мяса удалено все растворимое в воде и действующее химически, то это последнее действует теперь только как механический раздражитель и поэтому не может возбуждать деятельность пепсиновых желез. Вполне понятно, что непосредственно введенные в желудочную полость кусочки вареного. яичного белка совершенно не влияют на железы (Шифф [189] Лобасов и др.).
Все упомянутые вещества (мясо, вода, растворы экстрактовных веществ, продукты переваривания белковых веществ), если они непосредственно введены в кишечник, причем, конечно, должно быть исключено какое бы то ни было психическое воздействие, или вовсе не возбуждают пепсиновые железы, или в значительно более слабой степени, чем с желудка. (Тот факт, что вообще пепсиновые железы могут быть возбуждены с кишечника, наблюдался мною, 2 [190] 2 позднее Леконтом [191] 9). Подобные опыты могут быть проведены совершенно точным образом на собаках, оперированных по Соколову. Опыты над собаками с боковыми кишечными фистулами, без отделения желудка от кишечника, не могут разрешить этого вопроса удовлетворительным образом, ибо часть кишечного содержимого всегда может быть заброшена обратно, что очень легко происходит при известных условиях.
Мы воспроизводим здесь сравнительный опыт (табл. 18) с мясным фаршем; в этом виде мясо легче всего может быть введено в различные полости пищеварительного канала, без того, чтобы животное что-либо заметило (Соколов).
 Жидкий яичный белок, введенный прямо в кишечник, совсем не возбуждает секрецию желудочного сока, что видно уже из опытов с введением яичного белка в желудок, из которого он скоро переходит в кишечник.
Наконец все упомянутые вещества вводились per rectum в толстую кишку. Отсюда они не оказали абсолютно никакого действия на желудочные железы. В этом направлении доктор Лобасов продвинулся особенно энергично, применяя в клизмах огромные количества крепчайшего химического возбудителя пепсиновых желез, либиховского экстракта. Факт, что непосредственно введенная в желудок вода и таким же способом введенный раствор экстрактивных веществ действуют возбуждающим образом на работу пепсиновых желез, наблюдался многими клиницистами; совершенно так же они смогли доказать, что при ректальном введении тех же веществ этого действия не наблюдается.
В перечисленных до сих пор фактах, касающихся как акта еды, так и химического возбуждения пепсиновых желез, находит удовлетворительное объяснение нормальный ход желудочной секреции, наблюдаемой после кормления мясом. Возбужденное актом еды сокоотделение, наступающее через 5 минут после его начала, в первые четверти часа переходит в то, которое обусловливается значительным химическим действием мясной массы в жедудке. Этим обусловлено максимальное сокоотделение первых двух часов, из которых то один, то другой несколько перевешивает, смотря по тому, который из моментов одерживает верх. Как только пищевая кашица переходит в кишечник, с которого, как указано выше, она действует гораздо менее возбуждающим образом, секреция понемногу уменьшается и, наконец, падает до нуля, когда еще непереваренный и невсосавшийся остаток попадает в такие отрезки кишечного канала, с которых не проявляется никакого действия на пепсиновые железы.
Хлеб, как и вываренное мясо и вареный яичный белок, введенные непосредственно в желудок (и, конечно, при отсутствии всякого психического возбуждения), не оказывают влияния на пепсиновые железы. Значит, чтобы обусловить особенно длительный при кормлении хлебом секреторный период, необходимы химические раздражители, которые возникают в самом пищеварительном канале. Подобные раздражители могут быть даны либо в ротовой, либо в желудочной полости изливающимися в них соками; последние либо действуют сами секреторновозбуждающим образом, либо, благодаря им, содержащиеся в хлебной массе раздражающие вещества, - предполагая, что они там имеются, приобретают способность обнаруживать свое раздражающее действие. Непосредственно введенный в желудок хлеб не может раздражать пепсиновые железы, ибо он не содержит свободной воды, а содержащаяся в нем вода связана химически или физически. Так что можно ожидать, что слюна, которая, как упомянуто выше, в значительном количестве изливается на хлеб, может возбуждать деятельность пепсиновых желез самостоятельно (посредством своей воды) или также тем, что она растворяет содержашиеся, предположительно, в хлебе химически раздражающие вещества. Соколов поставил на своих собаках несколько опытов, в которых он вливал им непосредственно в желудок известное количество слюны; результат этих опытов был однообразный и точный: введенная в значительном количестве (от 100 до 200 куб. см) в желудок слюна обнаруживает секреторновозбуждающее действие, которое не только не уступает действию воды, но скорее несколько его превосходит.
Кроме воды, хлеб не содержит никаких готовых возбуждающих секрецию химических веществ: смесь накрошенного хлеба с водой действует с желудка не сильнее, а слабее, чем такой же объем воды.
Но на основании вышесказанного после того, как хлеб при нормальном акте еды вошел в соприкосновение с желудочным соком, в нем возникает некоторое количество продуктов переваривания белковых веществ, которые затем возбуждают дальнейшую секрецию желудочного сока. Ввиду того, что растительные белковые вещества очень трудно перевариваются и что к тому же их содержание в хлебе очень незначительно, количество накапливающихся в желудке продуктов переваривания не может быть никогда особенно большим. По Леконту, [192] введенный в кишечник 25% раствор глюкозы тормозит желудочную секрецию; раствор тростникового сахара действует так же, но слабее.
Только что обсужденные факты объясняют нам разницу в размере желудочной секреции после кормления хлебом между началом секреторного периода и его дальнейшим ходом. Вначале, после акта еды, секреторное раздражение обладает той же силой, как и при кормлении мясом, но позже, во время фазы химического действия, оно сразу становится явным образом слабее. Отсюда следует, что в течение второго часа обычно выделяется в два-три раза меньше желудочного сока, чем в течение первого. Чт Что вторая фаза обусловливается действием продуктов, получающихся при переваривании хлеба, может быть доказано еще тем, что у некоторых собак, которые вообще секретируют мало сока из изолированного маленького желудочка, часто наблюдается резкое падение, иногда даже остановка секреции после первого часа, после чего секреция затем опять начинается или нарастает. В уменьшении секреции по истечение первого часа, быть может, играет роль тормозящее влияние образующейся глюкозы.
Что касается химических раздражителей желудочной секреции, содержащихся в молоке, то здесь дело обстоит несколько сложнее. Влитое непосредственно в желудок молоко возбуждает секрецию желудочного сока, значит, оно содержит химически раздражающие вещества. Первым раздражителем является содержащаяся в нем вода. Затем действующие подобным образом вещества возникают из молочного белка, который особенно легко переваривается. Можно также предположить раздражающее действие некоторых других содержащихся в молоке веществ.
Однако к составным частям молока принадлежит еще одна, которая не только не возбуждает секреции желудочного сока, но, напротив, тормозит ее; это - жир. Первое лабораторное указание на то, что жир действует тормозящим образом на работу пепсиновых желез, мы находим у Хижина. Лобасов поставил значительное количество опытов, которые полностью подтверждают эти данные. Если влить собаке желудок 50-100 куб. см прованского масла и затем дать ей через час, например, мяса, то это последнее зачастую даже в течение часа не вызывает никакой желудочной секреции; начинающаяся после этого сегреция остается затем в течение долгого времени (в течение многих часов) очень скудной по сравнению с нормальной. Совершенно так же и в том случае, когда масло вливают в желудок после кормления мясом или хлебом, уже существующая секреция начинает вскоре уменьшаться и может даже совсем остановиться на некоторое время. Тормозящее действие жира на секрецию желудочного сока подтверждено также многочисленными наблюдениями на человеке.
Что это не является результатом механического действия масла, тормозящего доступ химически раздражающих веществ к желудочной стенке, но прямым, непосредственным воздействием на секреторный процесс, доказано в настоящее время многочисленными неопровержимыми опытами. Лобасов попробовал y эзофаготомированной собаки, у которой была желудочная фистула и изолированный маленький желудочек, посредством мнимого кормления, при прочих равных условиях, но один раз при пустом большом желудке, а в другой раз после введения известного количества жидкого масла, вызвать в них секрецию желудочного сока. Во втором ряде опытов из маленького желудочка либо вовсе не было секреции, либо она наступала позже и была скуднее, чем в первом ряде опытов. Эти опыты очень легко истолковать: секреторный процесс, наблюдаемый в железах изолированного желудочка и возбуждаемый актом еды, затормаживается маслом, действующим с остальной части пищеварительного канала.
Не менее убедительны были опыты Соколова, проделанные им на его сложным образом оперированных собаках. В качестве возбудителя сокоотделения в большой желудок вводилось мясо, где оно и оставалось лежать; масло вливалось через фистулу в duodenum, и только в этом случае наблюдалось значительное тормозящие секрецию действие. Из этого опыта ясно, что для возбуждения и торможения секреции желудочного сока служат две различные поверхности пищеварительного канала. И в этом случае, естественно, не может быть речи о косвенном тормозном действии жира. Дальнейшие наблюдения и опыты показали, что той ролью, которую жир играет в деятельности пепсиновых желез, не ограничивается его тормозящие секрецию действие. Нир, непосредственно введенный в желудок, в течение одного и даже двух часов не только не возбуждает деятельности желез, но тормозит, как мы видели, действие других возбуждающих секрецию веществ. Если же наблюдать собаку, получившую в желудок порядочную дозу жира, еще дальше, то можно видеть, что примерно на третьем часе начинает отделяться желудочный сок; эта секреция длится очень долго и дает немалое количество желудочного сока. Этот факт был тщательнейшим образом многократно проверен, дабы совершенно исключить какие бы то ни было психические воздействия. После всего этого должно было быть признано неопровержимым, что после введения жира в желудок очень поздно, не ранее, чем через два часа, возбуждается деятельность пепсиновых желез. Что же она теперь означает? Здесь можно сделать двоякое предположение: во-первых, не действуют ли секреторновозбуждающим образом соки, выбрасываемые из кишечника в желудок (подобное забрасывание пищеварительных соков из кишечника в желудок очень часто имеет место и, в частности, при богатов содержании жира в кишечном содержимом), и, во-вторых, не образуются ли действующие таким образом вещества при расщеплении жиров в кишечнике?
Была испытана правильность этих предположений. С одной стороны, Соколов наблюдал у своих собак значительное секреторновозбуждающее действие панкреатического сока и желчи, введенных в желудок. Однако вышеупомянутое действие жира не может быть во всем своем объеме объяснено, ибо обратное поступление вышеупомянутых соков далеко не так постоянно, как факт, который оно должно объяснить. В самом деле, Пионтковский [193] доказал, что глицерин вовсе не действует на желудочные железы, тогда как, наоборот, мыла, которые образуются из кислотных компонентов жира, являются очень энергичными возбудителями секреции желудочного сока. Они действуют с кишечника и обнаруживают свое действие, когда вводятся в него непосредственно. Если смешать жир с панкреатическим соком и желчью и оставить смесь на некоторое время в термостате, то после этого она возбуждает желудочную секрецию с кишечника столь же скоро. Олеиновая кислота и желчь, которые каждая сама по себе не действуют с кишечника на желудочную секрецию, делают это точнейшим образом, когда они смешаны вместе. Следует упомянуть, что если ввести в желудок или кишечник смесь жира и мыла, то тормозящее действие первого становится резко заметным и что при этом мыло обнаруживает свое действие позже и слабее. При этих обстоятельствах вполне понятно, что жир, который медленно поступает из желудка в кишечник и там окончательно расщепляется, так поздно начинает возбуждать желудочную секрецию. Стало быть, можно считать доказанным, что жир действует с кишечника на пепсиновые железы двояким образом: сам по себе он оказывает значительное тормозящее действие, а через мыла, возникающие из его продуктов расщепления, - сильное секреторновозбуждающее; оба действия проявляются с кишечника.
Упомянутые данные, касающиеся жира, способствуют удовлетворительному для нас объяснению хода секреции при некоторых сортах пищи, содержащих много жира, среди которых молоко, естественно, стоит на первом месте.
Когда узнали о тормозящем действии жира, то естественно пришли к предположению, что очень скудное вначале отделение желудочного сока при кормлении молоком должно быть объяснено действием жира. Это подтверждается разными вариантами опыта. Если дать животному сливок, то секреция еще скуднее и еще дольше остается таковой. Молоко, из которого фильтрацией удалены шарики жира, дает, безразлично - влито ли оно непосредственно в желудок или проглочено естественным образом, совершенно другую кривую секреции желудочного сока, причем в этом случае максимум секреции большею частью соответствует первому часу (Волкович [194] Наконец опыты Соколова, в которых молоко вливалось прямо в желудок оставалось там в течение всего времени, т. е. там, где оно не могло обнаружить своего тормозящего действия (которое ведь возникает только с кишечника), показали, что максимум секреции падал также на первый час.
Стало быть, при кормлении молоком возбуждение, вызванное актом еды и химическим действием других составных частей молока, подавляется жиром. Как же объяснить себе постепенное нарастание секреторного процесса почти до конца третьего часа? Есть ли это ослабление тормозящего действия или нарастание возбуждающего действия, или, наконец, и то и другое вместе? Первое могло бы обусловливаться расщеплением и постепенным исчезновением жира, последнее - постепенным накоплением продуктов переваривания молочного белка и образованием мыл. Более или менее значительное участие первых двух факторов после вышеуказанного ясно само собою. Ноо и последний фактор (образование мыл) играет большую роль в упомянутом явлении, об этом можно заключить из опытов, в которых животных кормили яичным желтком (Соборов 1 [195] и очень жирным мясом: различными натуральными сортами его (гусиным мясом, свининой), или постным, но с добавлением большого количества жира (Виршубский 2). 2 [196] Яичный желток дает огромный, но очень поздно наступающий максимум, падающий на четвертый-пятый час после кормления; совершенно так же ведет себя и жирное мясо. Но сначала секреция как в том, так и в другом случаях очень скудная, и только в течение часов она постепенно нарастает. Что здесь центр тяжести действительно лежит в тормозящем сначала действии жира и в позднейшем возбуждающем действии образующихся мыл, явствует из опытов Соколова. Скормленный собаке яичный желток в том случае, где желудок был связан с кишечником, дал после долго длящегося минимума значительный максимум только на четвертый-пятый час; в том же случае, где связь между желудком кишечником была нарушена, - гораздо меньший максимум и уже в течение первого часа секреции.
Описанные опыты, из которых обнаруживается двоякое действие жира, должны особенно интересовать клиницистов, и именно потому, что они, исходя из секреторнотормозящего действия, применяют жировую диету для лечения гиперсекреции. И здесь центр тяжести заключается в примененной дозе жира и в комбинации последнего с разными другими пищевыми веществами.
Собранного до сих пор материала достаточно, чтобы исчерпывающим образом объяснить касающийся количества желудочного сока ход его секреции при различных даваемых животному чистых сортах пищи.
Остается еще вопрос об изменениях состава желудочного сока в зависимости от различных сортов пищи и от времени секреторного периода.
В экспериментальном материале, касающемся чистого желудочного сока, мы до сегодняшнего дня не находим указаний на то, что секреция кислоты и воды протекает независимо друг от друга. Колебания кислотности желудочного сока наблюдаются постоянно и часто достигают значительной степени; но они всегда зависят от быстроты секреции, и именно следующим образом: чем значительнее быстрота, тем более высокую степень кислотности показывает желудочный сок, и наоборот. Ввиду этого факта можно предположить, что пепсиновые железы производят сок постоянно одинаковой кислотности и что наблюдаемая кислотность обусловливается имеющейся в большей или меньшей степени нейтрализацией кислоты щелочной желудочной слизью, которая к ней примешивается в то время, когда желудочный сок течет вдоль стенок то быстрее, то медленнее (Кетчер [197] ). Очень убедительное доказательство в пользу этого предположения можно видеть в том, что, как уже упоминалось выше, у голодной собаки, на которой повторно проделывается мнимое кормление, в конце концов, как только содержание хлора в организме в значительной степени уменьшится, совершенно останавливается секреция сока; при этом в выделяющемся желудочном соке до последних его порций кислотность колеблется в нормальных границах. Однако не исключена возможность, что существует более глубокая связь между кислотностью и скоростью секреции желудочного сока, совершенно так же, как это имеет место для слюны, в смысле содержания солей и скорости ее выделения. Если примкнуть к нашему толкованию, то, кроме того, можно ожидать колебаний кислотности и в зависимости от количества секретируемой слизи в желудке. Быть может, в некоторых особых случаях механизм колебаний кислотности состоит именно в этом. Легко усмотреть, что в случаях обиль ного выделения слизи этот факт может стать очень важным.
Совершенно иначе обстоит дело с содержанием ферментов в желудочном соке. Секреция кислого раствора секреция фермента (белковый фермент) являются двумя совершенно независимыми друг от друга функциями. Как уже видно было из приведенных раньше таблиц сокоотделения при различных сортах пищи, скорость секреции, т. е. секреция кислого раствора, и концентрация раствора, т. е. секреция фермента, образуют друг с другом чрезвычайно разнообразные комбинации. Как о неопровержимом факте можно говорить только о различных концентрациях фермента в желудочном соке и поэтому о разной скорости секреции фермента по сравнению с секрецией кислого раствора. Чтобы пойти дальше и говорить о процессе образования фермента в железе или о переходе фермента из латентного состояния в активный, как это делает Герцев, 1 [198] защищающий взгляд Шиффа и употребляющий обозначение пепсиногении, то для этого мы не располагаем в настоящее время достаточным материалом. Во всяком случае, на основании имеющегося материала можно утверждать, что в пепсиновых железах никогда не наблюдается то отсутствие пепсина (апепсия), из которого Герцен исходит в своих заключениях. Следует только вспомнить факт, как велика секреция желудочного сока при мнимом кормлении у животных, голодающих 14 дней. При этом сок обладает значительной переваривающей силой. Касательно секреции фермента известны два могущие служить мерилом факта. Добавка к введенным в желудок веществам (мясу, раствору либиховского экстракта) чистого крахмала ведет к увеличению количества фермента в выделенном желудочном соке. То же утверждает Герцен 2 2 [199] относительно некоторыгих углеводов. Добавка жира, наоборот, уменьшает содержание фермента в желудочном соке. Хотя жир вместе с тем тормозит и секрецию кислого раствора, часто оба влияния жира обнаруживаются не параллельно; слабая концентрация фермента наблюдается столь же часто как при скудной, так и при обильной секреции.
Приведенными фактами можно объяснить многое в колебаниях свойств желудочного сока при различной пище. Разная кислотность сортов желудочного сока, изливающихся на хлеб, мясо и молоко, зависит точным образом от средней скорости секреции за один час. В общем, быстрее всего сок выделяется в течение секреторного периода при кормлении мясом; соответственно с этим мясной сок содержит наивысшую кислотность. Хлебный сок как в первом, так и в последнем отношениях представляет прямую противоположность мясному соку. В сравнительно низкой кислотности хлебного соса частично повинна секреция слизи, которая при кормлении хлебом обильнее, чем при кормлении мясом и молоком. Значительная переваривающая сила хлебного сока объясняется тем, что в хлебе соединены вместе белок и углеводы; совершенно так же низкая переваривающая сила молочного сока объясняется тем, что в молоке соединены вместе белок и жиры. Переваривающая сила при кормлении сливками еще ниже, чем при кормлении молоком, очевидно потому, что здесь содержание жира еще значительнее. Переваривающая сила мясного сока стоит между переваривающей силой хлебного и молочного соков и скорее приближается к последней. С некоторым правом можно предположить. что и при кормлении мясом концентрация желудочного сока подавляется жиром, содержащимся в количестве нескольких процентов даже в тощем мясе. Раствор либиховского экстракта (экстрактивное вещество является той составной частью мяса, которая больше всего возбуждает секрецию мясногосока) дает желудочный сок с большей переваривающей силой, чем мясо. Меньшая переваривающая сила молочного сока, по сравнению c мясным соком, могла бы, возможно, зависеть от различного распределения жиров в массе обоих пищевых веществ.
Все выдвинутые до сих пор данные, при помощи которых должен быть объяснен механизм действия различных сортов пищи на секрецию пепсиновых желез, представляют, очевидно, лишь первую стадию анализа. Дальнейшая задача состоит в том, чтобы исследовать, каким образом определенные раздражения, действующие с определенных поверхностей пищеварительного канала, доходят до пепсиновых желез.
Возбуждение пепсиновых желез действующей на расстоянии пищей, так называемое психическое их возбуждение, может иметь место, конечно, только нервным путем. Само собою разумеется, что в настоящее время весь путь, который приходится проделать этим раздражениям, не может быть прослежен, ибо это явление принадлежит к наиболее сложным разыгрывающимся в нервной системе процессам. Объективный анализ этого психического раздражения, этого сложного нервного явления, еще почти не предпринят, и этот предмет до последнего времени трактовался с точки зрения субъективной психологии. Мы пытались связать это явление с тем, что мы знаем субъективно как аппетит, и назвали отделяющийся таким образом сок аппетитным соком. Мейсль [200] подвергнул анализу аппетит в его отношениях к секреции желудочного сока, исходя из психологической точки зрения. С чисто объективной физиологической точки зрения сделано для начала очень мало для объяснения этого явления. Некоторые авторы определенно установили, что никакое психическое возбуждение пепсиновых желез не имеет места, коль скоро у животного перерезаны nn. vagi. Гервер [201] утверждает, что по удалении некоторых участков мозговой коры пепсиновые железы не могут быть больше возбуждены психически. Искусственное возбуждение этих участков вызывает, по Герберу, секрецию желудочного сока; после перерезки nn. vagi это возбуждение остается недейственным.
Акт еды (опыт с мнимым кормлением) служит, как показано выше, очень сильным возбудителем пепсиновых желез. Уже из самого хода опыта явствует, что и здесь связь между актом еды и пепсиновыми железами чисто нервная. Пища так быстро проскальзывает по верхнему, к тому же обладающему очень малой всасывательной способностью, отрезку пищеварительного канала, что при этом вряд ли что-либо может попасть в кровь. Что связь здесь фактически исключительно нервная, следует из неопровержимо доказанного факта, чтомое кормление совершенно перестает действовать секреторновозбуждающим образом, как только перерезаны nn. vagi, будь то на шее или в грудной полости, или, наконец, под диафрагмой. Здесь, стало быть, наблюдаются те же отношения, как на слюнных железах после перерезки chorda tympani.
Результат опытов с мнимым кормлением до и после перерезки nn. vagi послужил вообще исходной точкой при изучении иннервационного аппарата пепсиновых желез. Из него неопровержимо следовало, что в блуждающих нервах проходят центробежные нервные волокна, через посредство которых раздражение переносится с центральной нервной системы на пепсиновые железы. Окончательным доказательством этого должны были послужить опыты, в которых раздражались периферические окончания этих нервов. Мы, совместно сумовой-Симановской, [202] поставили подобные опыты и смогли получить при этом абсолютно положительный результат. На собаках, через известные промежутки, в течение нескольких недель были проведены следующие операции: обычная желудочная фистула, эховаготомия, как она описана выше, перерезка правого n. vagus ниже ответвления n. laryngeus inferior. За день до опыта с раздражением на шее перерезался левый n. vagus, а затем его периферический конец, взятый на лигатуру, выводился непосредственно под кожу, которая зашивалась двумя-тремя швами. В день опыта у животного, поставленного в соответственный станок, снимались кожные швы и таким образом обнажался нерв. Раздражение нерва повторяющимися есекундно индукционными ударами вызывало значительную секрецию желудочного сока. Целью постановки опыта было исключить всякое постороннее чувствительное раздражение, ибо опыты Нечаева [203] показали, что чувствительные раздражения могут значительно уменьшить секрецию желудочного сока и даже приостановить ее на несколько часов. Позднейшие авторы (Аксенфельд, [204] Контежан, [205] Шнейер,3 [206] Ушаков [207] также смогли достигнуть положительных результатов в острых опытах на различных животных. Из некоторых других данных, состоявших в том, что между секрецией и усиленным кровообращением нельзя было установить постоянной и точной связи, что после введения атропина секреторное действие исчезало и что при усилении раздражения концентрация желудочного сока возрастала, следует заключить, что в n. vagus проходят специальные секреторные волокна к пепсиновым железам. На основании факта, что даже у собак, у которых было исключено всякое чувствительное раздражение, раздражение n. vagus вызывало секрецию желудочного сока лишь несколько десятков минут после начала раздражения, Ушаков считал себя вправе заключить, что в n. vages содержатся также особые волокна, тормозящие секрецию.
Если несомненен тот факт, что связь между актом еды и пепс новыми железами - связь исключительно нервная, то все же мнения различных авторов относительно механизма этого явления еще очень сильно расходятся. Что касается собак, то наша лаборатория в этом отношении стоит на той точке зрения, что в данном случае дело идет не о простом, исходящем из ротовой полости рефлексе, но о сложном нервном явлении (психическое возбуждение, оживленное внимание после еды, аппетит (Саноцкий) и др. [208] Этот взгляд основан на следующем. Раздражение того отдела пищеварительного аппарата, через который проходит пища при мнимом кормлении, различными раздражающими веществами - растворами кислот, солями, горькими веществами, эмульсией горчичного масла, камешками и т. д. - не дает ни одной капли желудочного сока при условии, чтобы избегалось всякое психическое возбуждение видом пищи и чтобы пепсиновые железы находились в состоянии покоя. Однако можно было бы предположить, что как раз химические свойства пищевых веществ действуют со рта как специфические раздражители. Но и это предположение опровергается следующим наблюдением. Если собака постоянно или в известное время предпочитает мясо хлебу или, наоборот, хлеб мясу, то только та еда действует как сильный возбудитель секреции, которую животное в данный момент охотно принимает; это наблюдается даже и в том случае, когда животное ест какой-нибудь другой сорт пищи, и экспериментатор в сравниваемых случаях заботится об одинаковой силе раздражения (например о величине и числе кусков). Наше предположение тем более оправдывается, что у некоторых особенно жадных животных секреция желудочного сока, вызванная действующей издали пищей, по своему объему абсолютно равна секреции, наблюдаемой после мнимого кормления. Вполне понятно, что при мнимом кормлении совершенно отпадают тормозящие психическое возбуждение моменты, играющие роль при возбуждении на расстоянии. Борисов, [209] наоборот, настаивает на том, что при акте еды дело идет о простом рефлекторном возбуждении с ротовой полости, и считает доказательством этого наблюдавшийся им факт, что при смазывании ротовой полости горькими веществами (которые и по Борисову сами по себе не возбуждают секрецию желудочного сока) мнимое кормление вызывает более значительную секрецию желудочного сока, чем до него. Но ясно, что здесь дело идет не о простом рефлексе, а о сложном воздействии; так называемое психическое возбуждение мы именно и рассматриваем как сложнонервное явление.
В наблюдениях и опытах, поставленных на людях, также подтверждается возбуждающее секрецию действие акта еды, но только и здесь мнения авторов о подробностях дела расходятся. Одни склонны признавать влияние акта жевания и веществ, чисто механически раздражающих ротовую полость; другие среди химических веществ наблюдали возбуждающее секрецию действие с ротовой полости только у пищевых веществ, и именно только тогда, когда данные вещества обладали нормальным, приятным для подопытного объекта вкусом и запахом. Новейшие, поставленные на людях опыть, наиболее безупречные в методическом отношении, говорят за взгляды, господствующие в нашей лаборатории (Горнборг 1). [210]
Анализ механизма действия пищи с желудка и с кишечника на секрецию желудочного сока сразу наталкивается на большие трудности. Так как в этих полостях возможен, и фактически наблюдается, переход пищевых веществ, продуктов их расщепления и переваривающих жидкостей в крови, то несомненно нужно считаться с двумя возможными родами связи между раздражением и секреторным элементом: с имеющим место через посредство нервной системы и через посредство крови. Из того открытия, что у пепсиновых желее, наверняка, имеется иннервационный аппарат, еще отнюдь не следует, что действие пищи в желудке и в кишечнике имеет место благодаря иннервационному аппарату. Возможно, что его участие ограничивается актом еды. Следовательно, этот вопрос должен быть разрешен специально для этой цели поставленными опытами. С одной стороны, надо было бы предпринять частичное или полное разрушение иннервационного аппарата; но оно не всегда выполнимо. С другой стороны, исследуемые вещества должны быть введены либо непосредственно в кровь, либо с других поверхностей пищеварительного канала, чем с нормальных. Введение пищевых веществ, а именно их смесей, непосредственно в кровь может так своеобразно и резко изменить состав крови, что на первый план выступает какая-нибудь другая физиологическая функция желез, а не их пищеварительная функция (например их способность освобождать кровь от вредных примесей, выравнивать состав крови). В этом случае, стало быть, действие было бы более значительным, чем при нормальном переходе этих веществ из кишечного канала в кровь; но результат может тоже быть и меньшим. Всасывание из различных отрезков пищеварительного канала определенными пищеварительными поверхностями может повести к тому, что в крове появятся вещества, которых не имеется во введенных смесях и которые происходят из самой кишечной стенки. Последнее обстоятельство недавно выдвинуто Бейлисом и Старлингом в их сообщении о поджелудочной железе, о котором мы еще сообщим в соответственном месте.
При существующих отношениях нельзя ожидать никаких окончательных результатов от уже произведенных аналитических исследований. Общий и несомненный результат всех как старых, так и новейших исследований сводится к тому, что секреция желудочного сока может иметь место и без nn. vagi. Особенно точное сравнительное исследование работы пепсиновых желез при нормальной еде до и после перерезки nn. vagi предпринято за последнее время Орбели [211] на изолированном желудочке. Сначала была исследована деятельность изолированного маленького желудочка, иннервационный аппарат которого оставался нетронутым, затем был перерезан серозно-мышечный мостик, соединяющий изолированный маленький желудочек с большим желудочком, т. е. ветви блуждающего нерва, и затем проведен ряд точно таких же исследований. Выполненные на двух собаках исследования дали абсолютно одинаковые результаты. Химические раздражители, как вода и раствор либиховского экстракта, действовавшие с большого желудка, вызывали в изолированном маленьком желудочке секрецию желудочного сока нормальной переваривающей силы, количество которого, однако, было меньше, чем в норме, самое большее вдвое. Возможно, что это количественное уменьшение основано не на устранении nn. vagi, принимающих в норме участие в процессе воздействия пищи с желудка и с кишечника, а на уменьшении работоспособности пепсиновых желез вообще, после перерезки nn. vagi. Тормозящее действие жира, проявляющееся в количестве сока, после отделения маленького желудочка от большого исчезает. Так как в этом случае при кормлении хлебом уменьшение пищеварительной силы было особенно резкое, то Орбели заключил из этого, что влияние крахмала или продуктов его переваривания на обогащение ферментом происходит также через посредство nn. vagi. Однако, согласно Герцену, 1 [212] углеводы увеличивают концентрацию фермента в желудочном соке и с rectum.
Чтобы наглядно показать в полной мере значение nn. vagi для нормальной работы пепсиновых желез, мы считаем нужным привести две относящиеся сюда таблицы (19 и 20) из сообщения Орбели.
Жидкий яичный белок, введенный прямо в кишечник, совсем не возбуждает секрецию желудочного сока, что видно уже из опытов с введением яичного белка в желудок, из которого он скоро переходит в кишечник.
Наконец все упомянутые вещества вводились per rectum в толстую кишку. Отсюда они не оказали абсолютно никакого действия на желудочные железы. В этом направлении доктор Лобасов продвинулся особенно энергично, применяя в клизмах огромные количества крепчайшего химического возбудителя пепсиновых желез, либиховского экстракта. Факт, что непосредственно введенная в желудок вода и таким же способом введенный раствор экстрактивных веществ действуют возбуждающим образом на работу пепсиновых желез, наблюдался многими клиницистами; совершенно так же они смогли доказать, что при ректальном введении тех же веществ этого действия не наблюдается.
В перечисленных до сих пор фактах, касающихся как акта еды, так и химического возбуждения пепсиновых желез, находит удовлетворительное объяснение нормальный ход желудочной секреции, наблюдаемой после кормления мясом. Возбужденное актом еды сокоотделение, наступающее через 5 минут после его начала, в первые четверти часа переходит в то, которое обусловливается значительным химическим действием мясной массы в жедудке. Этим обусловлено максимальное сокоотделение первых двух часов, из которых то один, то другой несколько перевешивает, смотря по тому, который из моментов одерживает верх. Как только пищевая кашица переходит в кишечник, с которого, как указано выше, она действует гораздо менее возбуждающим образом, секреция понемногу уменьшается и, наконец, падает до нуля, когда еще непереваренный и невсосавшийся остаток попадает в такие отрезки кишечного канала, с которых не проявляется никакого действия на пепсиновые железы.
Хлеб, как и вываренное мясо и вареный яичный белок, введенные непосредственно в желудок (и, конечно, при отсутствии всякого психического возбуждения), не оказывают влияния на пепсиновые железы. Значит, чтобы обусловить особенно длительный при кормлении хлебом секреторный период, необходимы химические раздражители, которые возникают в самом пищеварительном канале. Подобные раздражители могут быть даны либо в ротовой, либо в желудочной полости изливающимися в них соками; последние либо действуют сами секреторновозбуждающим образом, либо, благодаря им, содержащиеся в хлебной массе раздражающие вещества, - предполагая, что они там имеются, приобретают способность обнаруживать свое раздражающее действие. Непосредственно введенный в желудок хлеб не может раздражать пепсиновые железы, ибо он не содержит свободной воды, а содержащаяся в нем вода связана химически или физически. Так что можно ожидать, что слюна, которая, как упомянуто выше, в значительном количестве изливается на хлеб, может возбуждать деятельность пепсиновых желез самостоятельно (посредством своей воды) или также тем, что она растворяет содержашиеся, предположительно, в хлебе химически раздражающие вещества. Соколов поставил на своих собаках несколько опытов, в которых он вливал им непосредственно в желудок известное количество слюны; результат этих опытов был однообразный и точный: введенная в значительном количестве (от 100 до 200 куб. см) в желудок слюна обнаруживает секреторновозбуждающее действие, которое не только не уступает действию воды, но скорее несколько его превосходит.
Кроме воды, хлеб не содержит никаких готовых возбуждающих секрецию химических веществ: смесь накрошенного хлеба с водой действует с желудка не сильнее, а слабее, чем такой же объем воды.
Но на основании вышесказанного после того, как хлеб при нормальном акте еды вошел в соприкосновение с желудочным соком, в нем возникает некоторое количество продуктов переваривания белковых веществ, которые затем возбуждают дальнейшую секрецию желудочного сока. Ввиду того, что растительные белковые вещества очень трудно перевариваются и что к тому же их содержание в хлебе очень незначительно, количество накапливающихся в желудке продуктов переваривания не может быть никогда особенно большим. По Леконту, [192] введенный в кишечник 25% раствор глюкозы тормозит желудочную секрецию; раствор тростникового сахара действует так же, но слабее.
Только что обсужденные факты объясняют нам разницу в размере желудочной секреции после кормления хлебом между началом секреторного периода и его дальнейшим ходом. Вначале, после акта еды, секреторное раздражение обладает той же силой, как и при кормлении мясом, но позже, во время фазы химического действия, оно сразу становится явным образом слабее. Отсюда следует, что в течение второго часа обычно выделяется в два-три раза меньше желудочного сока, чем в течение первого. Чт Что вторая фаза обусловливается действием продуктов, получающихся при переваривании хлеба, может быть доказано еще тем, что у некоторых собак, которые вообще секретируют мало сока из изолированного маленького желудочка, часто наблюдается резкое падение, иногда даже остановка секреции после первого часа, после чего секреция затем опять начинается или нарастает. В уменьшении секреции по истечение первого часа, быть может, играет роль тормозящее влияние образующейся глюкозы.
Что касается химических раздражителей желудочной секреции, содержащихся в молоке, то здесь дело обстоит несколько сложнее. Влитое непосредственно в желудок молоко возбуждает секрецию желудочного сока, значит, оно содержит химически раздражающие вещества. Первым раздражителем является содержащаяся в нем вода. Затем действующие подобным образом вещества возникают из молочного белка, который особенно легко переваривается. Можно также предположить раздражающее действие некоторых других содержащихся в молоке веществ.
Однако к составным частям молока принадлежит еще одна, которая не только не возбуждает секреции желудочного сока, но, напротив, тормозит ее; это - жир. Первое лабораторное указание на то, что жир действует тормозящим образом на работу пепсиновых желез, мы находим у Хижина. Лобасов поставил значительное количество опытов, которые полностью подтверждают эти данные. Если влить собаке желудок 50-100 куб. см прованского масла и затем дать ей через час, например, мяса, то это последнее зачастую даже в течение часа не вызывает никакой желудочной секреции; начинающаяся после этого сегреция остается затем в течение долгого времени (в течение многих часов) очень скудной по сравнению с нормальной. Совершенно так же и в том случае, когда масло вливают в желудок после кормления мясом или хлебом, уже существующая секреция начинает вскоре уменьшаться и может даже совсем остановиться на некоторое время. Тормозящее действие жира на секрецию желудочного сока подтверждено также многочисленными наблюдениями на человеке.
Что это не является результатом механического действия масла, тормозящего доступ химически раздражающих веществ к желудочной стенке, но прямым, непосредственным воздействием на секреторный процесс, доказано в настоящее время многочисленными неопровержимыми опытами. Лобасов попробовал y эзофаготомированной собаки, у которой была желудочная фистула и изолированный маленький желудочек, посредством мнимого кормления, при прочих равных условиях, но один раз при пустом большом желудке, а в другой раз после введения известного количества жидкого масла, вызвать в них секрецию желудочного сока. Во втором ряде опытов из маленького желудочка либо вовсе не было секреции, либо она наступала позже и была скуднее, чем в первом ряде опытов. Эти опыты очень легко истолковать: секреторный процесс, наблюдаемый в железах изолированного желудочка и возбуждаемый актом еды, затормаживается маслом, действующим с остальной части пищеварительного канала.
Не менее убедительны были опыты Соколова, проделанные им на его сложным образом оперированных собаках. В качестве возбудителя сокоотделения в большой желудок вводилось мясо, где оно и оставалось лежать; масло вливалось через фистулу в duodenum, и только в этом случае наблюдалось значительное тормозящие секрецию действие. Из этого опыта ясно, что для возбуждения и торможения секреции желудочного сока служат две различные поверхности пищеварительного канала. И в этом случае, естественно, не может быть речи о косвенном тормозном действии жира. Дальнейшие наблюдения и опыты показали, что той ролью, которую жир играет в деятельности пепсиновых желез, не ограничивается его тормозящие секрецию действие. Нир, непосредственно введенный в желудок, в течение одного и даже двух часов не только не возбуждает деятельности желез, но тормозит, как мы видели, действие других возбуждающих секрецию веществ. Если же наблюдать собаку, получившую в желудок порядочную дозу жира, еще дальше, то можно видеть, что примерно на третьем часе начинает отделяться желудочный сок; эта секреция длится очень долго и дает немалое количество желудочного сока. Этот факт был тщательнейшим образом многократно проверен, дабы совершенно исключить какие бы то ни было психические воздействия. После всего этого должно было быть признано неопровержимым, что после введения жира в желудок очень поздно, не ранее, чем через два часа, возбуждается деятельность пепсиновых желез. Что же она теперь означает? Здесь можно сделать двоякое предположение: во-первых, не действуют ли секреторновозбуждающим образом соки, выбрасываемые из кишечника в желудок (подобное забрасывание пищеварительных соков из кишечника в желудок очень часто имеет место и, в частности, при богатов содержании жира в кишечном содержимом), и, во-вторых, не образуются ли действующие таким образом вещества при расщеплении жиров в кишечнике?
Была испытана правильность этих предположений. С одной стороны, Соколов наблюдал у своих собак значительное секреторновозбуждающее действие панкреатического сока и желчи, введенных в желудок. Однако вышеупомянутое действие жира не может быть во всем своем объеме объяснено, ибо обратное поступление вышеупомянутых соков далеко не так постоянно, как факт, который оно должно объяснить. В самом деле, Пионтковский [193] доказал, что глицерин вовсе не действует на желудочные железы, тогда как, наоборот, мыла, которые образуются из кислотных компонентов жира, являются очень энергичными возбудителями секреции желудочного сока. Они действуют с кишечника и обнаруживают свое действие, когда вводятся в него непосредственно. Если смешать жир с панкреатическим соком и желчью и оставить смесь на некоторое время в термостате, то после этого она возбуждает желудочную секрецию с кишечника столь же скоро. Олеиновая кислота и желчь, которые каждая сама по себе не действуют с кишечника на желудочную секрецию, делают это точнейшим образом, когда они смешаны вместе. Следует упомянуть, что если ввести в желудок или кишечник смесь жира и мыла, то тормозящее действие первого становится резко заметным и что при этом мыло обнаруживает свое действие позже и слабее. При этих обстоятельствах вполне понятно, что жир, который медленно поступает из желудка в кишечник и там окончательно расщепляется, так поздно начинает возбуждать желудочную секрецию. Стало быть, можно считать доказанным, что жир действует с кишечника на пепсиновые железы двояким образом: сам по себе он оказывает значительное тормозящее действие, а через мыла, возникающие из его продуктов расщепления, - сильное секреторновозбуждающее; оба действия проявляются с кишечника.
Упомянутые данные, касающиеся жира, способствуют удовлетворительному для нас объяснению хода секреции при некоторых сортах пищи, содержащих много жира, среди которых молоко, естественно, стоит на первом месте.
Когда узнали о тормозящем действии жира, то естественно пришли к предположению, что очень скудное вначале отделение желудочного сока при кормлении молоком должно быть объяснено действием жира. Это подтверждается разными вариантами опыта. Если дать животному сливок, то секреция еще скуднее и еще дольше остается таковой. Молоко, из которого фильтрацией удалены шарики жира, дает, безразлично - влито ли оно непосредственно в желудок или проглочено естественным образом, совершенно другую кривую секреции желудочного сока, причем в этом случае максимум секреции большею частью соответствует первому часу (Волкович [194] Наконец опыты Соколова, в которых молоко вливалось прямо в желудок оставалось там в течение всего времени, т. е. там, где оно не могло обнаружить своего тормозящего действия (которое ведь возникает только с кишечника), показали, что максимум секреции падал также на первый час.
Стало быть, при кормлении молоком возбуждение, вызванное актом еды и химическим действием других составных частей молока, подавляется жиром. Как же объяснить себе постепенное нарастание секреторного процесса почти до конца третьего часа? Есть ли это ослабление тормозящего действия или нарастание возбуждающего действия, или, наконец, и то и другое вместе? Первое могло бы обусловливаться расщеплением и постепенным исчезновением жира, последнее - постепенным накоплением продуктов переваривания молочного белка и образованием мыл. Более или менее значительное участие первых двух факторов после вышеуказанного ясно само собою. Ноо и последний фактор (образование мыл) играет большую роль в упомянутом явлении, об этом можно заключить из опытов, в которых животных кормили яичным желтком (Соборов 1 [195] и очень жирным мясом: различными натуральными сортами его (гусиным мясом, свининой), или постным, но с добавлением большого количества жира (Виршубский 2). 2 [196] Яичный желток дает огромный, но очень поздно наступающий максимум, падающий на четвертый-пятый час после кормления; совершенно так же ведет себя и жирное мясо. Но сначала секреция как в том, так и в другом случаях очень скудная, и только в течение часов она постепенно нарастает. Что здесь центр тяжести действительно лежит в тормозящем сначала действии жира и в позднейшем возбуждающем действии образующихся мыл, явствует из опытов Соколова. Скормленный собаке яичный желток в том случае, где желудок был связан с кишечником, дал после долго длящегося минимума значительный максимум только на четвертый-пятый час; в том же случае, где связь между желудком кишечником была нарушена, - гораздо меньший максимум и уже в течение первого часа секреции.
Описанные опыты, из которых обнаруживается двоякое действие жира, должны особенно интересовать клиницистов, и именно потому, что они, исходя из секреторнотормозящего действия, применяют жировую диету для лечения гиперсекреции. И здесь центр тяжести заключается в примененной дозе жира и в комбинации последнего с разными другими пищевыми веществами.
Собранного до сих пор материала достаточно, чтобы исчерпывающим образом объяснить касающийся количества желудочного сока ход его секреции при различных даваемых животному чистых сортах пищи.
Остается еще вопрос об изменениях состава желудочного сока в зависимости от различных сортов пищи и от времени секреторного периода.
В экспериментальном материале, касающемся чистого желудочного сока, мы до сегодняшнего дня не находим указаний на то, что секреция кислоты и воды протекает независимо друг от друга. Колебания кислотности желудочного сока наблюдаются постоянно и часто достигают значительной степени; но они всегда зависят от быстроты секреции, и именно следующим образом: чем значительнее быстрота, тем более высокую степень кислотности показывает желудочный сок, и наоборот. Ввиду этого факта можно предположить, что пепсиновые железы производят сок постоянно одинаковой кислотности и что наблюдаемая кислотность обусловливается имеющейся в большей или меньшей степени нейтрализацией кислоты щелочной желудочной слизью, которая к ней примешивается в то время, когда желудочный сок течет вдоль стенок то быстрее, то медленнее (Кетчер [197] ). Очень убедительное доказательство в пользу этого предположения можно видеть в том, что, как уже упоминалось выше, у голодной собаки, на которой повторно проделывается мнимое кормление, в конце концов, как только содержание хлора в организме в значительной степени уменьшится, совершенно останавливается секреция сока; при этом в выделяющемся желудочном соке до последних его порций кислотность колеблется в нормальных границах. Однако не исключена возможность, что существует более глубокая связь между кислотностью и скоростью секреции желудочного сока, совершенно так же, как это имеет место для слюны, в смысле содержания солей и скорости ее выделения. Если примкнуть к нашему толкованию, то, кроме того, можно ожидать колебаний кислотности и в зависимости от количества секретируемой слизи в желудке. Быть может, в некоторых особых случаях механизм колебаний кислотности состоит именно в этом. Легко усмотреть, что в случаях обиль ного выделения слизи этот факт может стать очень важным.
Совершенно иначе обстоит дело с содержанием ферментов в желудочном соке. Секреция кислого раствора секреция фермента (белковый фермент) являются двумя совершенно независимыми друг от друга функциями. Как уже видно было из приведенных раньше таблиц сокоотделения при различных сортах пищи, скорость секреции, т. е. секреция кислого раствора, и концентрация раствора, т. е. секреция фермента, образуют друг с другом чрезвычайно разнообразные комбинации. Как о неопровержимом факте можно говорить только о различных концентрациях фермента в желудочном соке и поэтому о разной скорости секреции фермента по сравнению с секрецией кислого раствора. Чтобы пойти дальше и говорить о процессе образования фермента в железе или о переходе фермента из латентного состояния в активный, как это делает Герцев, 1 [198] защищающий взгляд Шиффа и употребляющий обозначение пепсиногении, то для этого мы не располагаем в настоящее время достаточным материалом. Во всяком случае, на основании имеющегося материала можно утверждать, что в пепсиновых железах никогда не наблюдается то отсутствие пепсина (апепсия), из которого Герцен исходит в своих заключениях. Следует только вспомнить факт, как велика секреция желудочного сока при мнимом кормлении у животных, голодающих 14 дней. При этом сок обладает значительной переваривающей силой. Касательно секреции фермента известны два могущие служить мерилом факта. Добавка к введенным в желудок веществам (мясу, раствору либиховского экстракта) чистого крахмала ведет к увеличению количества фермента в выделенном желудочном соке. То же утверждает Герцен 2 2 [199] относительно некоторыгих углеводов. Добавка жира, наоборот, уменьшает содержание фермента в желудочном соке. Хотя жир вместе с тем тормозит и секрецию кислого раствора, часто оба влияния жира обнаруживаются не параллельно; слабая концентрация фермента наблюдается столь же часто как при скудной, так и при обильной секреции.
Приведенными фактами можно объяснить многое в колебаниях свойств желудочного сока при различной пище. Разная кислотность сортов желудочного сока, изливающихся на хлеб, мясо и молоко, зависит точным образом от средней скорости секреции за один час. В общем, быстрее всего сок выделяется в течение секреторного периода при кормлении мясом; соответственно с этим мясной сок содержит наивысшую кислотность. Хлебный сок как в первом, так и в последнем отношениях представляет прямую противоположность мясному соку. В сравнительно низкой кислотности хлебного соса частично повинна секреция слизи, которая при кормлении хлебом обильнее, чем при кормлении мясом и молоком. Значительная переваривающая сила хлебного сока объясняется тем, что в хлебе соединены вместе белок и углеводы; совершенно так же низкая переваривающая сила молочного сока объясняется тем, что в молоке соединены вместе белок и жиры. Переваривающая сила при кормлении сливками еще ниже, чем при кормлении молоком, очевидно потому, что здесь содержание жира еще значительнее. Переваривающая сила мясного сока стоит между переваривающей силой хлебного и молочного соков и скорее приближается к последней. С некоторым правом можно предположить. что и при кормлении мясом концентрация желудочного сока подавляется жиром, содержащимся в количестве нескольких процентов даже в тощем мясе. Раствор либиховского экстракта (экстрактивное вещество является той составной частью мяса, которая больше всего возбуждает секрецию мясногосока) дает желудочный сок с большей переваривающей силой, чем мясо. Меньшая переваривающая сила молочного сока, по сравнению c мясным соком, могла бы, возможно, зависеть от различного распределения жиров в массе обоих пищевых веществ.
Все выдвинутые до сих пор данные, при помощи которых должен быть объяснен механизм действия различных сортов пищи на секрецию пепсиновых желез, представляют, очевидно, лишь первую стадию анализа. Дальнейшая задача состоит в том, чтобы исследовать, каким образом определенные раздражения, действующие с определенных поверхностей пищеварительного канала, доходят до пепсиновых желез.
Возбуждение пепсиновых желез действующей на расстоянии пищей, так называемое психическое их возбуждение, может иметь место, конечно, только нервным путем. Само собою разумеется, что в настоящее время весь путь, который приходится проделать этим раздражениям, не может быть прослежен, ибо это явление принадлежит к наиболее сложным разыгрывающимся в нервной системе процессам. Объективный анализ этого психического раздражения, этого сложного нервного явления, еще почти не предпринят, и этот предмет до последнего времени трактовался с точки зрения субъективной психологии. Мы пытались связать это явление с тем, что мы знаем субъективно как аппетит, и назвали отделяющийся таким образом сок аппетитным соком. Мейсль [200] подвергнул анализу аппетит в его отношениях к секреции желудочного сока, исходя из психологической точки зрения. С чисто объективной физиологической точки зрения сделано для начала очень мало для объяснения этого явления. Некоторые авторы определенно установили, что никакое психическое возбуждение пепсиновых желез не имеет места, коль скоро у животного перерезаны nn. vagi. Гервер [201] утверждает, что по удалении некоторых участков мозговой коры пепсиновые железы не могут быть больше возбуждены психически. Искусственное возбуждение этих участков вызывает, по Герберу, секрецию желудочного сока; после перерезки nn. vagi это возбуждение остается недейственным.
Акт еды (опыт с мнимым кормлением) служит, как показано выше, очень сильным возбудителем пепсиновых желез. Уже из самого хода опыта явствует, что и здесь связь между актом еды и пепсиновыми железами чисто нервная. Пища так быстро проскальзывает по верхнему, к тому же обладающему очень малой всасывательной способностью, отрезку пищеварительного канала, что при этом вряд ли что-либо может попасть в кровь. Что связь здесь фактически исключительно нервная, следует из неопровержимо доказанного факта, чтомое кормление совершенно перестает действовать секреторновозбуждающим образом, как только перерезаны nn. vagi, будь то на шее или в грудной полости, или, наконец, под диафрагмой. Здесь, стало быть, наблюдаются те же отношения, как на слюнных железах после перерезки chorda tympani.
Результат опытов с мнимым кормлением до и после перерезки nn. vagi послужил вообще исходной точкой при изучении иннервационного аппарата пепсиновых желез. Из него неопровержимо следовало, что в блуждающих нервах проходят центробежные нервные волокна, через посредство которых раздражение переносится с центральной нервной системы на пепсиновые железы. Окончательным доказательством этого должны были послужить опыты, в которых раздражались периферические окончания этих нервов. Мы, совместно сумовой-Симановской, [202] поставили подобные опыты и смогли получить при этом абсолютно положительный результат. На собаках, через известные промежутки, в течение нескольких недель были проведены следующие операции: обычная желудочная фистула, эховаготомия, как она описана выше, перерезка правого n. vagus ниже ответвления n. laryngeus inferior. За день до опыта с раздражением на шее перерезался левый n. vagus, а затем его периферический конец, взятый на лигатуру, выводился непосредственно под кожу, которая зашивалась двумя-тремя швами. В день опыта у животного, поставленного в соответственный станок, снимались кожные швы и таким образом обнажался нерв. Раздражение нерва повторяющимися есекундно индукционными ударами вызывало значительную секрецию желудочного сока. Целью постановки опыта было исключить всякое постороннее чувствительное раздражение, ибо опыты Нечаева [203] показали, что чувствительные раздражения могут значительно уменьшить секрецию желудочного сока и даже приостановить ее на несколько часов. Позднейшие авторы (Аксенфельд, [204] Контежан, [205] Шнейер,3 [206] Ушаков [207] также смогли достигнуть положительных результатов в острых опытах на различных животных. Из некоторых других данных, состоявших в том, что между секрецией и усиленным кровообращением нельзя было установить постоянной и точной связи, что после введения атропина секреторное действие исчезало и что при усилении раздражения концентрация желудочного сока возрастала, следует заключить, что в n. vagus проходят специальные секреторные волокна к пепсиновым железам. На основании факта, что даже у собак, у которых было исключено всякое чувствительное раздражение, раздражение n. vagus вызывало секрецию желудочного сока лишь несколько десятков минут после начала раздражения, Ушаков считал себя вправе заключить, что в n. vages содержатся также особые волокна, тормозящие секрецию.
Если несомненен тот факт, что связь между актом еды и пепс новыми железами - связь исключительно нервная, то все же мнения различных авторов относительно механизма этого явления еще очень сильно расходятся. Что касается собак, то наша лаборатория в этом отношении стоит на той точке зрения, что в данном случае дело идет не о простом, исходящем из ротовой полости рефлексе, но о сложном нервном явлении (психическое возбуждение, оживленное внимание после еды, аппетит (Саноцкий) и др. [208] Этот взгляд основан на следующем. Раздражение того отдела пищеварительного аппарата, через который проходит пища при мнимом кормлении, различными раздражающими веществами - растворами кислот, солями, горькими веществами, эмульсией горчичного масла, камешками и т. д. - не дает ни одной капли желудочного сока при условии, чтобы избегалось всякое психическое возбуждение видом пищи и чтобы пепсиновые железы находились в состоянии покоя. Однако можно было бы предположить, что как раз химические свойства пищевых веществ действуют со рта как специфические раздражители. Но и это предположение опровергается следующим наблюдением. Если собака постоянно или в известное время предпочитает мясо хлебу или, наоборот, хлеб мясу, то только та еда действует как сильный возбудитель секреции, которую животное в данный момент охотно принимает; это наблюдается даже и в том случае, когда животное ест какой-нибудь другой сорт пищи, и экспериментатор в сравниваемых случаях заботится об одинаковой силе раздражения (например о величине и числе кусков). Наше предположение тем более оправдывается, что у некоторых особенно жадных животных секреция желудочного сока, вызванная действующей издали пищей, по своему объему абсолютно равна секреции, наблюдаемой после мнимого кормления. Вполне понятно, что при мнимом кормлении совершенно отпадают тормозящие психическое возбуждение моменты, играющие роль при возбуждении на расстоянии. Борисов, [209] наоборот, настаивает на том, что при акте еды дело идет о простом рефлекторном возбуждении с ротовой полости, и считает доказательством этого наблюдавшийся им факт, что при смазывании ротовой полости горькими веществами (которые и по Борисову сами по себе не возбуждают секрецию желудочного сока) мнимое кормление вызывает более значительную секрецию желудочного сока, чем до него. Но ясно, что здесь дело идет не о простом рефлексе, а о сложном воздействии; так называемое психическое возбуждение мы именно и рассматриваем как сложнонервное явление.
В наблюдениях и опытах, поставленных на людях, также подтверждается возбуждающее секрецию действие акта еды, но только и здесь мнения авторов о подробностях дела расходятся. Одни склонны признавать влияние акта жевания и веществ, чисто механически раздражающих ротовую полость; другие среди химических веществ наблюдали возбуждающее секрецию действие с ротовой полости только у пищевых веществ, и именно только тогда, когда данные вещества обладали нормальным, приятным для подопытного объекта вкусом и запахом. Новейшие, поставленные на людях опыть, наиболее безупречные в методическом отношении, говорят за взгляды, господствующие в нашей лаборатории (Горнборг 1). [210]
Анализ механизма действия пищи с желудка и с кишечника на секрецию желудочного сока сразу наталкивается на большие трудности. Так как в этих полостях возможен, и фактически наблюдается, переход пищевых веществ, продуктов их расщепления и переваривающих жидкостей в крови, то несомненно нужно считаться с двумя возможными родами связи между раздражением и секреторным элементом: с имеющим место через посредство нервной системы и через посредство крови. Из того открытия, что у пепсиновых желее, наверняка, имеется иннервационный аппарат, еще отнюдь не следует, что действие пищи в желудке и в кишечнике имеет место благодаря иннервационному аппарату. Возможно, что его участие ограничивается актом еды. Следовательно, этот вопрос должен быть разрешен специально для этой цели поставленными опытами. С одной стороны, надо было бы предпринять частичное или полное разрушение иннервационного аппарата; но оно не всегда выполнимо. С другой стороны, исследуемые вещества должны быть введены либо непосредственно в кровь, либо с других поверхностей пищеварительного канала, чем с нормальных. Введение пищевых веществ, а именно их смесей, непосредственно в кровь может так своеобразно и резко изменить состав крови, что на первый план выступает какая-нибудь другая физиологическая функция желез, а не их пищеварительная функция (например их способность освобождать кровь от вредных примесей, выравнивать состав крови). В этом случае, стало быть, действие было бы более значительным, чем при нормальном переходе этих веществ из кишечного канала в кровь; но результат может тоже быть и меньшим. Всасывание из различных отрезков пищеварительного канала определенными пищеварительными поверхностями может повести к тому, что в крове появятся вещества, которых не имеется во введенных смесях и которые происходят из самой кишечной стенки. Последнее обстоятельство недавно выдвинуто Бейлисом и Старлингом в их сообщении о поджелудочной железе, о котором мы еще сообщим в соответственном месте.
При существующих отношениях нельзя ожидать никаких окончательных результатов от уже произведенных аналитических исследований. Общий и несомненный результат всех как старых, так и новейших исследований сводится к тому, что секреция желудочного сока может иметь место и без nn. vagi. Особенно точное сравнительное исследование работы пепсиновых желез при нормальной еде до и после перерезки nn. vagi предпринято за последнее время Орбели [211] на изолированном желудочке. Сначала была исследована деятельность изолированного маленького желудочка, иннервационный аппарат которого оставался нетронутым, затем был перерезан серозно-мышечный мостик, соединяющий изолированный маленький желудочек с большим желудочком, т. е. ветви блуждающего нерва, и затем проведен ряд точно таких же исследований. Выполненные на двух собаках исследования дали абсолютно одинаковые результаты. Химические раздражители, как вода и раствор либиховского экстракта, действовавшие с большого желудка, вызывали в изолированном маленьком желудочке секрецию желудочного сока нормальной переваривающей силы, количество которого, однако, было меньше, чем в норме, самое большее вдвое. Возможно, что это количественное уменьшение основано не на устранении nn. vagi, принимающих в норме участие в процессе воздействия пищи с желудка и с кишечника, а на уменьшении работоспособности пепсиновых желез вообще, после перерезки nn. vagi. Тормозящее действие жира, проявляющееся в количестве сока, после отделения маленького желудочка от большого исчезает. Так как в этом случае при кормлении хлебом уменьшение пищеварительной силы было особенно резкое, то Орбели заключил из этого, что влияние крахмала или продуктов его переваривания на обогащение ферментом происходит также через посредство nn. vagi. Однако, согласно Герцену, 1 [212] углеводы увеличивают концентрацию фермента в желудочном соке и с rectum.
Чтобы наглядно показать в полной мере значение nn. vagi для нормальной работы пепсиновых желез, мы считаем нужным привести две относящиеся сюда таблицы (19 и 20) из сообщения Орбели.
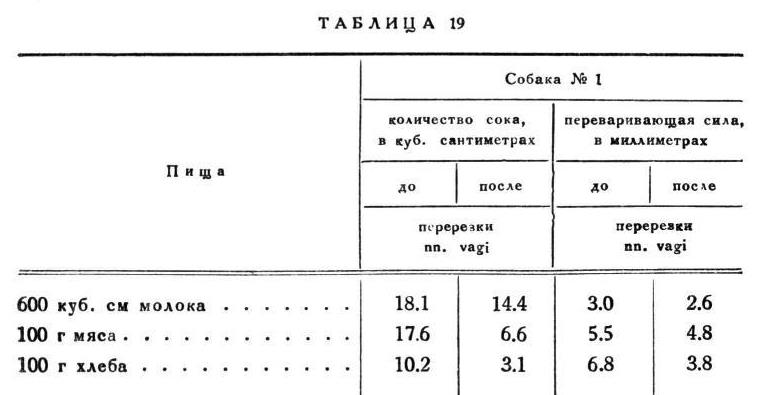 Быть может, химические раздражения действуют еще через посредство и других нервов, кроме nn.i. За участие n. sympathicus в секреции желудочного сока мог бы до известной степени говорить опыт Артуса,2 2 [213] состоящий в том, что вырезанный и вшитый в брюшную рану слизистой наружу кусок желудочной стенки никогда не дает кислой секреции. Попельский [214] разрушал все нервные связи желудка с центральной нервной системой, а также и ganglion solaris, и, несмотря на это, действие либиховского экстракта на пепсиновые железы сохранялось. Ввиду этого факта пришлось допустить либо наличие периферического рефлекторного аппарата в самой желудочной стенке (что предполагает Попельский), либо переданное через кровь действие. Так как рефлекторный аппарат удалить из желудочной стенки невозможно, то следует стремиться достигнуть конечного результата, исключая вторую возможность. Животным повторно вводились per rectum растворы экстрактивных веществ мяса, причем они не имели никакого возбуждающего секрецию действия на желудок. Лобасов [215] поставил подобные опыты с особой тщательностью, но всегда с отрицательным результатом. Прямое внутривенное введение либиховского экстракта вызывало - у собак Лобасова скудную секрецию желудочного сока, причем, однако, одновременно наблюдалась картина общего возбуждения, рвотные движения и словоотделение, так что этот опыт вряд ли может служить доказательством секреторновозбуждающего действия экстрактивных веществ через кровь при нормальных условиях. В опытах Соколова такой раствор действовал гораздо слабее, чем с желудка, хотя он вливался в duodenum. Подобное же поведение наблюдалось и при применении воды. Относительно последней пришлось признать как непреложный факт, что она возбуждает пепсиновые железы не тем, что она попадает в кровь. Наконец было доказано (Саноцкий [216] "), что атропин, парализующий секреторные нервы пепсиновых желез, останавливает всякую секрецию желудочного сока. Таким образом, значит, очень вероятно действие через посредство нервной системы. Однако нельзя совершенно исключить вторую возможность, ибо предположение, что, быть может, из резорбированных желудочной стенкой воды или раствора либиховского экстракта воспринимается что-либо раздражающее, не было проверено соответственными опытами.
Быть может, химические раздражения действуют еще через посредство и других нервов, кроме nn.i. За участие n. sympathicus в секреции желудочного сока мог бы до известной степени говорить опыт Артуса,2 2 [213] состоящий в том, что вырезанный и вшитый в брюшную рану слизистой наружу кусок желудочной стенки никогда не дает кислой секреции. Попельский [214] разрушал все нервные связи желудка с центральной нервной системой, а также и ganglion solaris, и, несмотря на это, действие либиховского экстракта на пепсиновые железы сохранялось. Ввиду этого факта пришлось допустить либо наличие периферического рефлекторного аппарата в самой желудочной стенке (что предполагает Попельский), либо переданное через кровь действие. Так как рефлекторный аппарат удалить из желудочной стенки невозможно, то следует стремиться достигнуть конечного результата, исключая вторую возможность. Животным повторно вводились per rectum растворы экстрактивных веществ мяса, причем они не имели никакого возбуждающего секрецию действия на желудок. Лобасов [215] поставил подобные опыты с особой тщательностью, но всегда с отрицательным результатом. Прямое внутривенное введение либиховского экстракта вызывало - у собак Лобасова скудную секрецию желудочного сока, причем, однако, одновременно наблюдалась картина общего возбуждения, рвотные движения и словоотделение, так что этот опыт вряд ли может служить доказательством секреторновозбуждающего действия экстрактивных веществ через кровь при нормальных условиях. В опытах Соколова такой раствор действовал гораздо слабее, чем с желудка, хотя он вливался в duodenum. Подобное же поведение наблюдалось и при применении воды. Относительно последней пришлось признать как непреложный факт, что она возбуждает пепсиновые железы не тем, что она попадает в кровь. Наконец было доказано (Саноцкий [216] "), что атропин, парализующий секреторные нервы пепсиновых желез, останавливает всякую секрецию желудочного сока. Таким образом, значит, очень вероятно действие через посредство нервной системы. Однако нельзя совершенно исключить вторую возможность, ибо предположение, что, быть может, из резорбированных желудочной стенкой воды или раствора либиховского экстракта воспринимается что-либо раздражающее, не было проверено соответственными опытами.
 Как уже упомянуто, работой пепсиновых желез при кормлении чистой пищей не исчерпывается вся жизнедеятельность этих желез; они должны еще, кроме того, так или иначе реагировать в интересах организма на раздражение, оказываемое другими пищевыми веществами, попадающими вместе с пищей в желудочно-кишечный канал.
Весь относящийся сюда материал еще не может быть приведен в настоящее время в систематический вид.
Ввиду значительного практического и теоретического интереса, представляемого этим предметом, мы кратко упомянем об одном веществе, представляющем нечто среднее между настоящими пищевыми веществами и другими случайными раздражителями желудочно-кишечного канала. Мы подразумеваем алкоголь, который за последнее время в значительной мере привлекает внимание как экспериментаторов, так и клиницистов. Все единогласно утверждают, что он действует на пепсиновые железы как энергичный возбудитель секреции. Также вряд ли может быть подвергнуто сомнению, что алкоголь действует непосредственно на секреторную клетку. Это, однако, не исключает возможности, что это действие передается также через нервную систему. За это последнее предположение как будто говорит более значительный секреторный эффект алкоголя с желудка, чем с rectum. Равным образом некоторые авторы утверждают, что изливающийся на алкоголь сок содержит мало фермента. Было бы, однако, преувеличением утверждать, что на алкоголь может изливаться совершенно бессерментный сок, т. е. чистый раствор соляной кислоты. Полученные в этом направлении данные должны быть отнесены за счет недостаточно совершенной методики.
Кроме желудочного сока и на той же поверхности, как этот последний, в желудке производится специальными секреторными элементами, покровным эпителием, еще другой секрет - слизь. К сожалению, нам известно очень мало относительно физиологии этого секрета. Естественно, и она меняется, что касается как ее количества, так и качества. Выделенная слизь то вязкая, то жидкая; иногда, как, например, при кормлении мясом, и именно в определенные часы, вообще не наблюдается никакой секреции слизи; при кормлении хлебом эта секреция, правда, скудная, но постоянная. Огромные количества слизи выделяются, когда в желудок попадают сильно местно раздражающие вещества. Это является, как следует предположить, главным приемом для обезвреживания этих веществ, совершенно так же, как это достигается при помощи слюны. Что в секреции слизи, по крайней мере частично, принимает участие нервная система, доказывается, с одной стороны, опытами Ушакова, [217] который наблюдал, как при раздражении n. vagus в начале этого раздражения изливалась совершенно чистая слизь в обильном количестве, с другой стороне, тем фактом, что при кормлении хлебом в изолированном маленьком желудочке наблюдается отделение слизи, как только хлеб подействует на поверхность большого желудка. Однако и здесь может быть предположено непосредственное раздражение эпителиальных клеток данным веществом.
Как уже упомянуто, работой пепсиновых желез при кормлении чистой пищей не исчерпывается вся жизнедеятельность этих желез; они должны еще, кроме того, так или иначе реагировать в интересах организма на раздражение, оказываемое другими пищевыми веществами, попадающими вместе с пищей в желудочно-кишечный канал.
Весь относящийся сюда материал еще не может быть приведен в настоящее время в систематический вид.
Ввиду значительного практического и теоретического интереса, представляемого этим предметом, мы кратко упомянем об одном веществе, представляющем нечто среднее между настоящими пищевыми веществами и другими случайными раздражителями желудочно-кишечного канала. Мы подразумеваем алкоголь, который за последнее время в значительной мере привлекает внимание как экспериментаторов, так и клиницистов. Все единогласно утверждают, что он действует на пепсиновые железы как энергичный возбудитель секреции. Также вряд ли может быть подвергнуто сомнению, что алкоголь действует непосредственно на секреторную клетку. Это, однако, не исключает возможности, что это действие передается также через нервную систему. За это последнее предположение как будто говорит более значительный секреторный эффект алкоголя с желудка, чем с rectum. Равным образом некоторые авторы утверждают, что изливающийся на алкоголь сок содержит мало фермента. Было бы, однако, преувеличением утверждать, что на алкоголь может изливаться совершенно бессерментный сок, т. е. чистый раствор соляной кислоты. Полученные в этом направлении данные должны быть отнесены за счет недостаточно совершенной методики.
Кроме желудочного сока и на той же поверхности, как этот последний, в желудке производится специальными секреторными элементами, покровным эпителием, еще другой секрет - слизь. К сожалению, нам известно очень мало относительно физиологии этого секрета. Естественно, и она меняется, что касается как ее количества, так и качества. Выделенная слизь то вязкая, то жидкая; иногда, как, например, при кормлении мясом, и именно в определенные часы, вообще не наблюдается никакой секреции слизи; при кормлении хлебом эта секреция, правда, скудная, но постоянная. Огромные количества слизи выделяются, когда в желудок попадают сильно местно раздражающие вещества. Это является, как следует предположить, главным приемом для обезвреживания этих веществ, совершенно так же, как это достигается при помощи слюны. Что в секреции слизи, по крайней мере частично, принимает участие нервная система, доказывается, с одной стороны, опытами Ушакова, [217] который наблюдал, как при раздражении n. vagus в начале этого раздражения изливалась совершенно чистая слизь в обильном количестве, с другой стороне, тем фактом, что при кормлении хлебом в изолированном маленьком желудочке наблюдается отделение слизи, как только хлеб подействует на поверхность большого желудка. Однако и здесь может быть предположено непосредственное раздражение эпителиальных клеток данным веществом.

 Наконец и разделенный по часам ход секреции является также особым для каждого сорта пищи. При кормлении хлебом и мясом энергичная секреция падает на первые два часа, причем во втором случае секреция быстро падает до нуля, тогда как при кормлении хлебом к первому периоду примыкает второй секреторный период, в течение которого секреция, правда, слабее, но все же падает до нуля только постепенно. При кормлении молоком, наоборот, в течение первых двух часов секреция очень вялая и часто даже в течение второго часа слабее, чем в первый, и лишь в течение третьего часа развивается бросающийся в глаза максимум секреции, после которого она быстро падает до нуля.
Оставив пока в стороне обсуждение физиологического значения секреции того или иного фермента, так как в нашем распоряжении не имеется относящихся сюда точных данных, мы рассмотрим ближе количество сока, т. е. количество раствора соды. Первая физиологическая задача кишечного пищеварения состоит, очевидно, в том, что необходимая для химических реакций в желудке сильнокислая среда должна быть превращена в нейтральную или даже в щелочную, как это требуется для действия кишечных ферментов и прежде всего для ферментов панкреатического сока. Так как при кормлении молоком желудочный сок, с одной стороны, нейтрализуется, а с другой стороны, очень сильно разбавляется, то соответственно с этим требуется гораздо меньшее количество щелочи, чтобы изменить реакцию. Это и на самом деле имеет место, ибо на молоко изливается особенно мало панкреатического сока, причем его щелочность, как и щелочность его золы, значительно меньше, чем при других родах пищи. При вышеприведенных порциях при кормлении хлебом в кишечник попадает в целом больше кислоты, чем при кормлении мясом, и соответственно с этим наибольшее количество панкреатического сока изливается на хлеб. Это имеет тем большее значение, что из всех панкреатических ферментов тот, который действует на крахмал, меньше всего может довольствоваться даже слабокислой реакцией. Даже если не считаться с настоящими определениями содержания фермента в панкреатическом соке, а руководствоваться только определениями органического вещества, содержание которого в общем параллельно содержанию фермента, то несомненно можно усмотреть некоторые бросающиеся в глаза закономерные отношения. На вышеприведенные количества мяса и молока железа выделяет почти одинаковые количества органических веществ, принимая, что количество сока берется в расчет наряду с концентрацией фермента. На хлеб же, наоборот, изливается почти вдвое больше сока. И это соответствует как меньшей перевариваемости хлебного белка по сравнению с мясным и молочным белком, так и громадному содержанию крахмала противовес содержанию жира в молоке и мясе.
То же самое подтверждают и проведенные определения ферментов (Вальтер).
Наконец и разделенный по часам ход секреции является также особым для каждого сорта пищи. При кормлении хлебом и мясом энергичная секреция падает на первые два часа, причем во втором случае секреция быстро падает до нуля, тогда как при кормлении хлебом к первому периоду примыкает второй секреторный период, в течение которого секреция, правда, слабее, но все же падает до нуля только постепенно. При кормлении молоком, наоборот, в течение первых двух часов секреция очень вялая и часто даже в течение второго часа слабее, чем в первый, и лишь в течение третьего часа развивается бросающийся в глаза максимум секреции, после которого она быстро падает до нуля.
Оставив пока в стороне обсуждение физиологического значения секреции того или иного фермента, так как в нашем распоряжении не имеется относящихся сюда точных данных, мы рассмотрим ближе количество сока, т. е. количество раствора соды. Первая физиологическая задача кишечного пищеварения состоит, очевидно, в том, что необходимая для химических реакций в желудке сильнокислая среда должна быть превращена в нейтральную или даже в щелочную, как это требуется для действия кишечных ферментов и прежде всего для ферментов панкреатического сока. Так как при кормлении молоком желудочный сок, с одной стороны, нейтрализуется, а с другой стороны, очень сильно разбавляется, то соответственно с этим требуется гораздо меньшее количество щелочи, чтобы изменить реакцию. Это и на самом деле имеет место, ибо на молоко изливается особенно мало панкреатического сока, причем его щелочность, как и щелочность его золы, значительно меньше, чем при других родах пищи. При вышеприведенных порциях при кормлении хлебом в кишечник попадает в целом больше кислоты, чем при кормлении мясом, и соответственно с этим наибольшее количество панкреатического сока изливается на хлеб. Это имеет тем большее значение, что из всех панкреатических ферментов тот, который действует на крахмал, меньше всего может довольствоваться даже слабокислой реакцией. Даже если не считаться с настоящими определениями содержания фермента в панкреатическом соке, а руководствоваться только определениями органического вещества, содержание которого в общем параллельно содержанию фермента, то несомненно можно усмотреть некоторые бросающиеся в глаза закономерные отношения. На вышеприведенные количества мяса и молока железа выделяет почти одинаковые количества органических веществ, принимая, что количество сока берется в расчет наряду с концентрацией фермента. На хлеб же, наоборот, изливается почти вдвое больше сока. И это соответствует как меньшей перевариваемости хлебного белка по сравнению с мясным и молочным белком, так и громадному содержанию крахмала противовес содержанию жира в молоке и мясе.
То же самое подтверждают и проведенные определения ферментов (Вальтер).
 Если дело идет о том, чтобы получить воронка, достаточно коротко подрезать ножницами шерсть; брить ее нельзя. Далее, чтобы не причинить животному боль и не вырвать у него все волосы на этом месте, следует немного нагреть замазку, что лучше всего сделать, проводя нагретой металлической палочкой по краю воронки. На нижнем конце воронки при помощи той же замазки укрепляются два проволочных крючочка, на которые при помощи проволочных петель подвешиваются точно градуированные цилиндрики с делениями на пятые или десятые доли кубического санслюну в абсолютно чистом виде, необходимо укрепить над отверстием цилиндра у нижнего конца воронки нечто вроде предохранительной крышки из мягкого прозрачного и водонепроницаемого материала, дабы в цилиндр не попало что-либо из вводимых в рот животного порошкасбразных или жидких веществ. Конечно, и здесь была бы желательной автоматическая регистрация, но до сих пор еще нет безукоризненной методики.
На первом месте среди вивисекций, которые служат для анализа секреторной функции, стоят операции на нервах, имеющих отношение к железам, с целью их перерезки или раздражения.
Чтобы обнажить chorda tympani, секреторный черепно-мозговой нерв подчелюстной и подъязычной желез, кожный разрез проводят так же, как при обнажении их выводных протоков, только разрез удлиняют кзади и перерезают m. mylohyoideus вплоть до его заднего края. Как только отпрепарована внешняя половина этой мышцы, становится виден идущий поперек раны n. lingualis. Если проследить этот нерв в сторону от пересекающего ero ductus whartonianus, то обнаруживается маленькая ответвляющаяся назад от нерва ветовка - chorda tympani. На месте ее ответвления от n. lingualis она может быть после некоторой препаровки или просто перерезана, или после перерезки взята на лигатуру и раздражаема либо простым электродом, либо электродом для глубоко лежащих нервов. Чтобы по возможности сохранить возбудимость нерва и чтобы раздражать все секреторные волокна слюнных желез, особенно подъязычной, рекомендуется как можно ниже отпрепаровать вместо chorda tympani самый n. lingualis и там его перерезать. На электрод берется его периферический конец и в нем также и chorda tympani.
Если дело идет о том, чтобы получить воронка, достаточно коротко подрезать ножницами шерсть; брить ее нельзя. Далее, чтобы не причинить животному боль и не вырвать у него все волосы на этом месте, следует немного нагреть замазку, что лучше всего сделать, проводя нагретой металлической палочкой по краю воронки. На нижнем конце воронки при помощи той же замазки укрепляются два проволочных крючочка, на которые при помощи проволочных петель подвешиваются точно градуированные цилиндрики с делениями на пятые или десятые доли кубического санслюну в абсолютно чистом виде, необходимо укрепить над отверстием цилиндра у нижнего конца воронки нечто вроде предохранительной крышки из мягкого прозрачного и водонепроницаемого материала, дабы в цилиндр не попало что-либо из вводимых в рот животного порошкасбразных или жидких веществ. Конечно, и здесь была бы желательной автоматическая регистрация, но до сих пор еще нет безукоризненной методики.
На первом месте среди вивисекций, которые служат для анализа секреторной функции, стоят операции на нервах, имеющих отношение к железам, с целью их перерезки или раздражения.
Чтобы обнажить chorda tympani, секреторный черепно-мозговой нерв подчелюстной и подъязычной желез, кожный разрез проводят так же, как при обнажении их выводных протоков, только разрез удлиняют кзади и перерезают m. mylohyoideus вплоть до его заднего края. Как только отпрепарована внешняя половина этой мышцы, становится виден идущий поперек раны n. lingualis. Если проследить этот нерв в сторону от пересекающего ero ductus whartonianus, то обнаруживается маленькая ответвляющаяся назад от нерва ветовка - chorda tympani. На месте ее ответвления от n. lingualis она может быть после некоторой препаровки или просто перерезана, или после перерезки взята на лигатуру и раздражаема либо простым электродом, либо электродом для глубоко лежащих нервов. Чтобы по возможности сохранить возбудимость нерва и чтобы раздражать все секреторные волокна слюнных желез, особенно подъязычной, рекомендуется как можно ниже отпрепаровать вместо chorda tympani самый n. lingualis и там его перерезать. На электрод берется его периферический конец и в нем также и chorda tympani.
 Для демонстрации во время лекции механизма рефлекторного раздражения слюнных желез проще всего пользоваться подчелюстной железой и именно следующим образом. Кончик языка слабо отравленного кураре животного окунают в чашечку c 0.5%-м раствором соляной кислоты. Вслед за этим следует истечение слюны через введенную в ductus whartonianus канюлю. Затем оба nervi lingualis перерезаются периферически от места перекреста их с выводными протоками. Погружение кончика языка в кислый раствор повторяется, но теперь остается безрезультатным. Затем раздражают конец соответственного n. lingualis. Снова следует секреция. Наконец перерезают chorda tympani на месте ее ответвления от n. lingsalis. Такое же раздражение n. lingualis остается теперь без результата. Под конец производится раздражение периферического конца chorda tympani.
Обнажение n. auriculotemporalis, содержащего секреторные волокна для околоушной железы, Навроцкий описывает следующим образом: «Сперва я обнажаю во всю ее длину m. digastricus maxillae, изолируя ее от остальных мышц, дважды перевязываю (чтобы предупредить возможность кровотечения), перерезаю ее между обеими лигатурами, отпрепаровываю и откидываю ее заднюю часть назад. Далее я проникаю (у корня языка) до головки сустава нижней челюсти; это достигается лучше всего без помощи режущих инструментов; иногда приходится перевязывать, чтобы легче было проникнуть в глубину, некоторые мелкие кровеносные сосуды; m. pterygoideus internus (именно его задний край) начисто отпрепаровывают, поднимая его кверху при помощи тупого крючка, и одно за другим осторожно перерезают его волокна, пока не обнажится n. alveolaris вместе с n. lingualis; почти под прямым углом к направлению этих нервов проходит n. auriculotemporalis, обычно прикрытый веной; осторожно удалив соединительную ткань, поднимают при помощи крючка n. auriculotemporalis. Как только под нерв подведена лигатура, дальнейшие изолирование и перерезку уже легко выполнить».
Секреторные черепно-мозговые нер трех больших слюнных желез можно с удобством одновременно раздражать в барабанной полости. Гейденгайн [0] описывает соответственную операцию следующим образом: «Вскрытие барабанной полости осуществляется на собаке без труда, если после кожного разреза идти в глубину по внутреннему краю заднего конца m. biventer, снаружи от корня языка. На этоместе можно с легкостью нащупать bulla ossea, нажав пальцем на мягкие части. Перерезав соединительную ткань между m. biventer и его внутренними соседями, оттягивают двумя сильными и широкими крючками первый кнаружи, а вторые кнутри и, медленно разрывая двумя пинцетами соединительную ткань, без единой капли крови достигают таким образом до bulla. По удалении надкостницы обращенную вперед и кнаружи плоскую поверхность ее просверливают маленьким трепаном и расширяют отверстие костными щипцами, пока не откроются promontorium и свободно лежащий на нем нерв. Для их раздражения я располагаю около нерва на кости округленные и близко друг к другу расположенные концевые пуговки вставленных в каучук двух жестких электродных проволочек и фиксирую их держалкой; таким образом нерв подвергается только действию петель тока. Если наполнить барабанную полость нейтральной жидкостью, например кровью, то легко удается придать электродам такое положение, при котором не только якобсонов нерв, но и chorda будут подвергаться. действию петель тока достаточной густоты, чтобы привести в деятельное состояние все три слюнных железы».
Методика препаровки ираздражения ramus buccinatorius nervi trigemini, секреторного нерва глазничной железы, разработана Лавдовским совместно с Гейденгайном и будет передана здесь в изложении Лавдовского: «После того как у кураризованного животного (у собаки возможно большего размера) налажено искусственное дыхание и его голове придано устойчивое положение, а именно такое, чтобы одна из двух сторон была обращена к эспериментатору, операция должна начинаться кожным разрезом, в 3-4 см, у переднего края m. masseter. Если мы затем осторожно освободим лежащую под ним фасцию, то, как правило, мы тотчас же наталкиваемся на довольно большую ветвь лицевой вены, которую нужно дважды перевязать и перерезать между лигатурами. В вилке между передним краем m. masseter и buccinatorius разрывают далее соединительную ткань, и тогда в ней находят на высоте зубов нижней челюсти n. buccinatorius. После этого на мышечную массу m. masseter накладываются две лигатуры, и мышца между ними перерезается. (Так как мы тут же натыкаемся на processus coronoideus и вынуждены проникнуть рукой глубоко за него, то лучше всего раскрыть пасть собаки и фиксировать ее в таком положении деревянной распоркой). Перерезав m. masseter, мы сперва очищаем processus coronoideus, затем осторожно разламываем его сразу или по кусочкам костными щипцами. Отыскав теперь в переднем углу раны ramus buccinatorius, мы прослеживаем его как можно дальше в глубину, причем мы как можно бережнее отделяем соединительную ткань, которая тянется к orbita. Как только удалось обнажить нерв до такой глубины, что становится доступным достаточно длинный его отрезок, мы тотчас же берем его на лигатуру и крепко его перевязываем; наконец, перерезав его, мы освобождаем вместе с лигатурой его периферический отрезок. Соединить с ним теперь влектрод дело одного мгновения.
«В течение каждого подобного опыта, особенно во время перерезки m. masseter, получаются очень сильные кровотечения. Останавливая кровь, надо быть очень осторожным, дабы избежать повреждения нерва, особенно я еще раз это подчеркиваю - при скалывании processus coronoideus».
Что касается выхода церебральных секреторных волокон слюнных желез из мозга, то относящиеся сюда операции должны быть описаны в отделе о мозговых нервах.
Симпатические секреторные нервы слюнных желез отыскиваются с целью раздражения или перерезки в двух местах. Чаще всего для этого препаруется на середине шеи n. vagosympathicus. Кожный разрез, длиною в 2-3 см, проводится между m. cleidomastoideus и мышцами, лежащими на трахее. В глубине раны нащупывают и находят art. carotis. N. vagosympathicus плотно сращен с артерией, так что его приходится отпрепаровывать. По отпрепарованному нерву проводится острым ножом продольный разрез, причем из надрезанноболочки выступает n. vagus в виде центрального тяжа, тогда как п. sympathicus остается в оболочке. Реже отпрепаровываются верхний симпатический ганглий и ответвляющиеся от него к железам ветви. У собаки проще всего добраться до ганглия следующим путем. Кожный разрез, длиною в 4-5 см, накладывается кнаружи от корня языка и щитовидного хряща вниз, начиная на высоте корня языка. Чтобы выгадать место в ране, можно удалить расположенные в верхней ее части лимфатические железы. Вскоре в ране бросается в глаза дугообразно идущий в глубину n. hypoglossus. Теперь нужно только, проникая в глубину в направлении n. hypoglossus, осторожно отпрепаровать ткани в разные стороны. Осторожность особенно нужна тогда, когда достигаешь до art. carotis externa, ибо очень легко, препаруя в глубине, перервать одну из ее веточек. Если соблюдать необходимую осторожность, можно без капли крови дойти до внутреннего края bulla ossea височной кости, на которой расположена группа черепных нервов, вместе с n. sympathicus; верхний ганглий последнего можно легко узнать по его форме и серой окраске. Идя от него, нетрудно проследить и нервную ветвь, идущую вместе с art. carotis.
Для опытов по изучению кровообращения слюнных желез пользуются почти исключительно подчелюстной железой, и именно ее венами. В большинстве случаев в целях наблюдения над кровообращением аппараты, измеряющие протекающее через железу количество крови, соединяются верхним концом v. jugularis externa (В. Фрей [10], Ленгли [1], Буртов-Опиц [12]). Эти аппараты будут описаны в отделе кровообращения. Операция обнажения вен производится следующим образом. Кожный разрез, длиною в 4 -5 см, проводится от угла нижней челюсти кзади, параллельно средней линии; он проходит как раз над серединой gl. submaxillaris. Отпрепаровав кожу и m. subcutaneus в обе стороны, ясно видишь vv. maxillaris interna et externa, огибающие железу снаружи и снутри и соединяющиеся сразу за железой в v. jugularis externa. V. maxillaris externa разделяется на половине высоты железы на v. facialis и v. lingualis. Одна или две вены подчелюстной слюнной железы впадают в одну или две из вышеназванных вен. Если соединить ствол v. jugularis externa вблизи ее образования из составляющих ее вен с тем или другим аппаратом, измеряющим кровяной ток, то нужно все вышеупомянутые вены перевязать дистально от тех мест, где в них впадают маленькие вены слюнной железы. При этом, конечно, должны быть приняты все меры для предотвращения свертывания крови.
Совершенно таким же образом для простого получения крови из железы, например для опытов над газообменом железы, пользуются преимущественно v. jugularis externa, причем в нее вставляется Т-образная канюля (Баркрофт [15]).
Д Для решения некоторых вопросов физиологии желез измеряют также объем железы при различных условиях. Подобные измерения были проведены на gl. submaxillaris. Плетизмограф состоял из резиновой капсулы со стеклянным кружком на одной стороне, так что было возможно точно определить на глаз до какой степени поверхностные сосуды наполнены кровью. Мелеза обнажалась у угла нижней челюсти и освобождалась от своей капсулы; ее выводной проток также отпрепаровывался на известном протяжении, для чего приходилось перерезать m. digastricus и отпрепаровывать оба его конца. При случае перевязывались одна-две маленькие вены, когда они выступали из железы в наиболее отдаленных от hilus пунктах. Большие же вены, наоборот, тщательно сохранялись. Сосуды и выводной проток железы проводились через отверстие капсулы; оставшаяся свободной часть отверстия затыкалась ватой и густым вазелином. Прежде чем помещать в него железу, плетизмограф подогревался; кроме того, в нем находилось - для сбережения железы - немного смоченной в теплом нормальном солевом растворе ваты. Капсула соединялась с барабанчиком или с регистрирующим рычагом посредством резиновой трубки, укрепленной на стеклянной трубочке, проведенной через стенку капсулы. Весь аппарат наполнялся воздухом.T-образная трубка, вставленная в резиновую трубку, позволяла в любой момент увеличивать или уменьшать давление воздуха в капсуле (Бэнч [14]). [14]
Для исследования лимфатической секреции подчелюстной слюнной железы пользуются тем лимфатическим сосудом, который лежит на правой стороне шеи рядом с art. carotis communis. Разрез тогда делается такой же, как при препаровке n. vagosympathicus. В надрезанный шейный сосуд вводится канюля, соединенная с градуированной трубкой; последняя позволяет определять количество выделившейся лимфы. Так как спонтанно лимфа течет через канюлю лишь в очень незначительных количествах, то через определенные промежутки времени массируют шею. Само собой разумеется, что массаж должен быть во всех отношениях равномерным. Так как при этих условиях количество выделившейся лимфы все же подвергается большим колебаниям, то, во избежание ошибок, нужно пользоваться средними числами из целой серии опытов (Бейнбридж [15]).
При исследовании gl. submaxillaris применялся также метод изолирования органа. Большим разрезом, проведенным несколько наискось прямо мимо угла нижней челюсти, обнажалась gl. submaxillaris c окружающими ее вышеописанными венами, также m. digastricus. Последний перерезался поперек, и задний конец его отпрепаровывался в глубину вплоть до места прикрепления. При этом в глубине раны открывалась art. carotis externa с отходящей от нее кверху art. maxillaris externa. Первою от art. maxillaris externa отделяется вниз веточка к железе. Ha art. maxillaris externa, ниже ответвления железистой веточки, накладывается лигатура. Две лигатуры подводятся под ту же артерию как раз на том месте, где она ответвляется от art. carotis externa. После этого вокруг железы перевязываются все вены, упомянутые выше по поводу операций, предназначенных для исследования кровообращения железы. В выводной проток железы вставляется канюля. Как только все это проделано, в art. maxillaris externa и v. jugularis externa вводятся канюли. Тогда велеза с частью окружающей ее ткани быстро вырезается и помещается в соответственный аппарат, где приступают к искусственному кровоснабжению железы. Если в случае необходимости при вышеописанной операции обнажается chorda tympani, она отпрепаровывается, перерезается, и нижний конец ее берется на лигатуру ру (Овсяницкий [16]).
Когда нужно определить вес железы и содержание в ней азота, то, чтобы полностью отпрепаровать железу и не спутать части gl. submaxillaris с частями непосредственно к ней прилегающей gl. sublingualis, в последнюю вспрыскивают какую-нибудь окрашенную жидкость; в виде красящего вещества можно употреблять индиго-сернокислый натрий, кармин и т. д.
Для демонстрации во время лекции механизма рефлекторного раздражения слюнных желез проще всего пользоваться подчелюстной железой и именно следующим образом. Кончик языка слабо отравленного кураре животного окунают в чашечку c 0.5%-м раствором соляной кислоты. Вслед за этим следует истечение слюны через введенную в ductus whartonianus канюлю. Затем оба nervi lingualis перерезаются периферически от места перекреста их с выводными протоками. Погружение кончика языка в кислый раствор повторяется, но теперь остается безрезультатным. Затем раздражают конец соответственного n. lingualis. Снова следует секреция. Наконец перерезают chorda tympani на месте ее ответвления от n. lingsalis. Такое же раздражение n. lingualis остается теперь без результата. Под конец производится раздражение периферического конца chorda tympani.
Обнажение n. auriculotemporalis, содержащего секреторные волокна для околоушной железы, Навроцкий описывает следующим образом: «Сперва я обнажаю во всю ее длину m. digastricus maxillae, изолируя ее от остальных мышц, дважды перевязываю (чтобы предупредить возможность кровотечения), перерезаю ее между обеими лигатурами, отпрепаровываю и откидываю ее заднюю часть назад. Далее я проникаю (у корня языка) до головки сустава нижней челюсти; это достигается лучше всего без помощи режущих инструментов; иногда приходится перевязывать, чтобы легче было проникнуть в глубину, некоторые мелкие кровеносные сосуды; m. pterygoideus internus (именно его задний край) начисто отпрепаровывают, поднимая его кверху при помощи тупого крючка, и одно за другим осторожно перерезают его волокна, пока не обнажится n. alveolaris вместе с n. lingualis; почти под прямым углом к направлению этих нервов проходит n. auriculotemporalis, обычно прикрытый веной; осторожно удалив соединительную ткань, поднимают при помощи крючка n. auriculotemporalis. Как только под нерв подведена лигатура, дальнейшие изолирование и перерезку уже легко выполнить».
Секреторные черепно-мозговые нер трех больших слюнных желез можно с удобством одновременно раздражать в барабанной полости. Гейденгайн [0] описывает соответственную операцию следующим образом: «Вскрытие барабанной полости осуществляется на собаке без труда, если после кожного разреза идти в глубину по внутреннему краю заднего конца m. biventer, снаружи от корня языка. На этоместе можно с легкостью нащупать bulla ossea, нажав пальцем на мягкие части. Перерезав соединительную ткань между m. biventer и его внутренними соседями, оттягивают двумя сильными и широкими крючками первый кнаружи, а вторые кнутри и, медленно разрывая двумя пинцетами соединительную ткань, без единой капли крови достигают таким образом до bulla. По удалении надкостницы обращенную вперед и кнаружи плоскую поверхность ее просверливают маленьким трепаном и расширяют отверстие костными щипцами, пока не откроются promontorium и свободно лежащий на нем нерв. Для их раздражения я располагаю около нерва на кости округленные и близко друг к другу расположенные концевые пуговки вставленных в каучук двух жестких электродных проволочек и фиксирую их держалкой; таким образом нерв подвергается только действию петель тока. Если наполнить барабанную полость нейтральной жидкостью, например кровью, то легко удается придать электродам такое положение, при котором не только якобсонов нерв, но и chorda будут подвергаться. действию петель тока достаточной густоты, чтобы привести в деятельное состояние все три слюнных железы».
Методика препаровки ираздражения ramus buccinatorius nervi trigemini, секреторного нерва глазничной железы, разработана Лавдовским совместно с Гейденгайном и будет передана здесь в изложении Лавдовского: «После того как у кураризованного животного (у собаки возможно большего размера) налажено искусственное дыхание и его голове придано устойчивое положение, а именно такое, чтобы одна из двух сторон была обращена к эспериментатору, операция должна начинаться кожным разрезом, в 3-4 см, у переднего края m. masseter. Если мы затем осторожно освободим лежащую под ним фасцию, то, как правило, мы тотчас же наталкиваемся на довольно большую ветвь лицевой вены, которую нужно дважды перевязать и перерезать между лигатурами. В вилке между передним краем m. masseter и buccinatorius разрывают далее соединительную ткань, и тогда в ней находят на высоте зубов нижней челюсти n. buccinatorius. После этого на мышечную массу m. masseter накладываются две лигатуры, и мышца между ними перерезается. (Так как мы тут же натыкаемся на processus coronoideus и вынуждены проникнуть рукой глубоко за него, то лучше всего раскрыть пасть собаки и фиксировать ее в таком положении деревянной распоркой). Перерезав m. masseter, мы сперва очищаем processus coronoideus, затем осторожно разламываем его сразу или по кусочкам костными щипцами. Отыскав теперь в переднем углу раны ramus buccinatorius, мы прослеживаем его как можно дальше в глубину, причем мы как можно бережнее отделяем соединительную ткань, которая тянется к orbita. Как только удалось обнажить нерв до такой глубины, что становится доступным достаточно длинный его отрезок, мы тотчас же берем его на лигатуру и крепко его перевязываем; наконец, перерезав его, мы освобождаем вместе с лигатурой его периферический отрезок. Соединить с ним теперь влектрод дело одного мгновения.
«В течение каждого подобного опыта, особенно во время перерезки m. masseter, получаются очень сильные кровотечения. Останавливая кровь, надо быть очень осторожным, дабы избежать повреждения нерва, особенно я еще раз это подчеркиваю - при скалывании processus coronoideus».
Что касается выхода церебральных секреторных волокон слюнных желез из мозга, то относящиеся сюда операции должны быть описаны в отделе о мозговых нервах.
Симпатические секреторные нервы слюнных желез отыскиваются с целью раздражения или перерезки в двух местах. Чаще всего для этого препаруется на середине шеи n. vagosympathicus. Кожный разрез, длиною в 2-3 см, проводится между m. cleidomastoideus и мышцами, лежащими на трахее. В глубине раны нащупывают и находят art. carotis. N. vagosympathicus плотно сращен с артерией, так что его приходится отпрепаровывать. По отпрепарованному нерву проводится острым ножом продольный разрез, причем из надрезанноболочки выступает n. vagus в виде центрального тяжа, тогда как п. sympathicus остается в оболочке. Реже отпрепаровываются верхний симпатический ганглий и ответвляющиеся от него к железам ветви. У собаки проще всего добраться до ганглия следующим путем. Кожный разрез, длиною в 4-5 см, накладывается кнаружи от корня языка и щитовидного хряща вниз, начиная на высоте корня языка. Чтобы выгадать место в ране, можно удалить расположенные в верхней ее части лимфатические железы. Вскоре в ране бросается в глаза дугообразно идущий в глубину n. hypoglossus. Теперь нужно только, проникая в глубину в направлении n. hypoglossus, осторожно отпрепаровать ткани в разные стороны. Осторожность особенно нужна тогда, когда достигаешь до art. carotis externa, ибо очень легко, препаруя в глубине, перервать одну из ее веточек. Если соблюдать необходимую осторожность, можно без капли крови дойти до внутреннего края bulla ossea височной кости, на которой расположена группа черепных нервов, вместе с n. sympathicus; верхний ганглий последнего можно легко узнать по его форме и серой окраске. Идя от него, нетрудно проследить и нервную ветвь, идущую вместе с art. carotis.
Для опытов по изучению кровообращения слюнных желез пользуются почти исключительно подчелюстной железой, и именно ее венами. В большинстве случаев в целях наблюдения над кровообращением аппараты, измеряющие протекающее через железу количество крови, соединяются верхним концом v. jugularis externa (В. Фрей [10], Ленгли [1], Буртов-Опиц [12]). Эти аппараты будут описаны в отделе кровообращения. Операция обнажения вен производится следующим образом. Кожный разрез, длиною в 4 -5 см, проводится от угла нижней челюсти кзади, параллельно средней линии; он проходит как раз над серединой gl. submaxillaris. Отпрепаровав кожу и m. subcutaneus в обе стороны, ясно видишь vv. maxillaris interna et externa, огибающие железу снаружи и снутри и соединяющиеся сразу за железой в v. jugularis externa. V. maxillaris externa разделяется на половине высоты железы на v. facialis и v. lingualis. Одна или две вены подчелюстной слюнной железы впадают в одну или две из вышеназванных вен. Если соединить ствол v. jugularis externa вблизи ее образования из составляющих ее вен с тем или другим аппаратом, измеряющим кровяной ток, то нужно все вышеупомянутые вены перевязать дистально от тех мест, где в них впадают маленькие вены слюнной железы. При этом, конечно, должны быть приняты все меры для предотвращения свертывания крови.
Совершенно таким же образом для простого получения крови из железы, например для опытов над газообменом железы, пользуются преимущественно v. jugularis externa, причем в нее вставляется Т-образная канюля (Баркрофт [15]).
Д Для решения некоторых вопросов физиологии желез измеряют также объем железы при различных условиях. Подобные измерения были проведены на gl. submaxillaris. Плетизмограф состоял из резиновой капсулы со стеклянным кружком на одной стороне, так что было возможно точно определить на глаз до какой степени поверхностные сосуды наполнены кровью. Мелеза обнажалась у угла нижней челюсти и освобождалась от своей капсулы; ее выводной проток также отпрепаровывался на известном протяжении, для чего приходилось перерезать m. digastricus и отпрепаровывать оба его конца. При случае перевязывались одна-две маленькие вены, когда они выступали из железы в наиболее отдаленных от hilus пунктах. Большие же вены, наоборот, тщательно сохранялись. Сосуды и выводной проток железы проводились через отверстие капсулы; оставшаяся свободной часть отверстия затыкалась ватой и густым вазелином. Прежде чем помещать в него железу, плетизмограф подогревался; кроме того, в нем находилось - для сбережения железы - немного смоченной в теплом нормальном солевом растворе ваты. Капсула соединялась с барабанчиком или с регистрирующим рычагом посредством резиновой трубки, укрепленной на стеклянной трубочке, проведенной через стенку капсулы. Весь аппарат наполнялся воздухом.T-образная трубка, вставленная в резиновую трубку, позволяла в любой момент увеличивать или уменьшать давление воздуха в капсуле (Бэнч [14]). [14]
Для исследования лимфатической секреции подчелюстной слюнной железы пользуются тем лимфатическим сосудом, который лежит на правой стороне шеи рядом с art. carotis communis. Разрез тогда делается такой же, как при препаровке n. vagosympathicus. В надрезанный шейный сосуд вводится канюля, соединенная с градуированной трубкой; последняя позволяет определять количество выделившейся лимфы. Так как спонтанно лимфа течет через канюлю лишь в очень незначительных количествах, то через определенные промежутки времени массируют шею. Само собой разумеется, что массаж должен быть во всех отношениях равномерным. Так как при этих условиях количество выделившейся лимфы все же подвергается большим колебаниям, то, во избежание ошибок, нужно пользоваться средними числами из целой серии опытов (Бейнбридж [15]).
При исследовании gl. submaxillaris применялся также метод изолирования органа. Большим разрезом, проведенным несколько наискось прямо мимо угла нижней челюсти, обнажалась gl. submaxillaris c окружающими ее вышеописанными венами, также m. digastricus. Последний перерезался поперек, и задний конец его отпрепаровывался в глубину вплоть до места прикрепления. При этом в глубине раны открывалась art. carotis externa с отходящей от нее кверху art. maxillaris externa. Первою от art. maxillaris externa отделяется вниз веточка к железе. Ha art. maxillaris externa, ниже ответвления железистой веточки, накладывается лигатура. Две лигатуры подводятся под ту же артерию как раз на том месте, где она ответвляется от art. carotis externa. После этого вокруг железы перевязываются все вены, упомянутые выше по поводу операций, предназначенных для исследования кровообращения железы. В выводной проток железы вставляется канюля. Как только все это проделано, в art. maxillaris externa и v. jugularis externa вводятся канюли. Тогда велеза с частью окружающей ее ткани быстро вырезается и помещается в соответственный аппарат, где приступают к искусственному кровоснабжению железы. Если в случае необходимости при вышеописанной операции обнажается chorda tympani, она отпрепаровывается, перерезается, и нижний конец ее берется на лигатуру ру (Овсяницкий [16]).
Когда нужно определить вес железы и содержание в ней азота, то, чтобы полностью отпрепаровать железу и не спутать части gl. submaxillaris с частями непосредственно к ней прилегающей gl. sublingualis, в последнюю вспрыскивают какую-нибудь окрашенную жидкость; в виде красящего вещества можно употреблять индиго-сернокислый натрий, кармин и т. д.
 Операция эзофаготомии также очень проста и при соблюдении известных условий удается всегда. Разрез, длиною в 10 см и больше, смотря по собаке, проводится по левой стороне шеи по внутреннему краю m. sternocleidomastoideus, начинаясь у внутреннего края щитовидного хряща разрезается, и пространство между m. sternocleidomastoideus и лежащими на трахее мышцами отпрепаровывается. Под трахеей, по середине раны, пищевод захватывается пэаном, отпрепаровывается по всей своей окружности на 2 3 см в длину и полностью перерезается поперек. Быстро вытянув концы закрыв их планами или тампонами, нужно теперь позаботиться том, чтобы слюна из пищевода совершенно не попадала в образующуюся глубокую рану. Концы пищевода подшиваются к раневым углам, причем иглой нужно обкалывать не только кожу, но мышцы, ограничивающие рану. Этим достигается то, что концы пищевода не втягиваются внутрь, но навсегда остаются на поверхности. Середина раны между вшитыми в углы раны концами пищевода тщательно зашивается как послойно в глубине, так на кожной поверхности. Таким образом между концами пищевода получается порядочный промежуток, образованный из сросшейся кожи, что исключает возможность того, чтобы даже самые незначительные количества слизи или составных частей пищи попадали из верхнего конца пищевода в нижний. Главное правило операции, гарантирующее правильное и быстрое заживление, заключается в том, чтобы операцию производили абсолютно чисто, избегая загрязнения глубокой раны, где нагноение, раз начавшись, распространяется быстро и интенсивно. По той же причине и после операции нужно ежедневно обмывать рану антисептической жидкостью; лучше даже повторять это несколько раз в день, так как рана постоянно очень сильно загрязняется снаружи слюной. Наложение желудочной фистулы должно предшествовать эзофаготомии на 3-4 недели, т. е. эзофаготомия проделывается после того, как образовалась рубцовая ткань, крепко обхватывающая канюлю.
Сперва, до полного заживления раны, собаку кормят через желудочную фистулу. Твердая пища вкладывается в желудок кусочками через открытую фистулу; жидкости вливаются, причем канюля закрывается простым вентилем, вставленным в просверленную пробку. В дальнейшем вся еда - жидкость вместе с размельченной твердой пищей - удобно и быстро вливается в нижний конец пищевода через воронку с толстой резиновой трубкой, при стоячем положении собаки.
Принимая во внимание потерю слюны и особенно часто повторяемое собирание желудочного сока, необходимо вводить собаке много жидкости. Если нужно, можно таким способом без вреда для животного ежедневно собирать желудочный сок в течение очень долгого времени. Получаемое каждый раз количество сока может достигать у больших собак 1.5 л. В то время как у нее описанным способом собирают сок, собаку можно оставлять без присмотра, если ставить перед ней на подставке довольно широкую чашку с кусочками какой-нибудь пищи так, чтобы собака могла сама до нее дотянуться. Проглоченный кусок каждый раз падает обратно в чашку. Под желудочную фистулу подвешивается довольно широкий, толстостенный суживающийся книзу в трубку цилиндр. В том месте, где цилиндр суживается, кладется стеклянная вата или лигнин, чтобы задерживать слизь. На суженный конец надевается довольно длинная резиновая трубка, другой конец которой соединяется несколько в стороне от животного, либо прямо, либо при помощи стеклянного или каучукового крючка с толстостенным, шарообразно расширяющимся книзу сосудом; в этом последнем и собирается весь чистый желудочный сок. У некоторых животных иногда в желудок забрасывается кишечное содержимое, что сразу можно узнать по окраске желудочного содержимого желчью. В подобном случае желудок очищается тщательным промыванием водой, и тогда собирание желудочного сока может быть возобновлено. Само собой разумеется, в день собирания желудочного сока собаку с утра не кормят. Несмотря на часто повторяемое обильное собирание сока, эти собаки живут в полном здоровии и остаются в течение многих лет совершенно точно работающими фабриками желудочного сока.
Для получения чистого желудочного сока служит также операция Фремона [2], которая состоит в полном изолировании всего желудка от остального пищеваритель поперечные разрезы как на границе между пищеводом и желудком, так и между желудком и двенадцатиперстной кишкой. Нижний конец пищевода сшивается с двенадцатиперстной кишкой, и таким образом восстанавливается непрерывность пищеварительного канала. Зашитый же с двух концов желудок опоражнивается через обыкновенную желудочную фистулу. Представляя решительное преимущество перед методикой одновременных желудочной и пищеводной фистул в том отношении, что забрасывание кишечного содержимого в желудок здесь совершенно исключено, эта методика все же сильно ей уступает; это касается как трудности самой операции (ввиду необходимости сохранять совершенно неповрежденными nn. vagi), трудно сохранить на более продолжительное время здоровыми и живыми оперированных таким образом собак.
Хотя обе описанные методики вполне пригодны для получения чистого желудочного сока, все же они не могут претендовать на то, чтобы годиться и для полного анализа нормальной работы пепсиновых желез. При обеих методиках не происходит соприкосновения пиши со стенками желудка, тогда как именно это соприкосновение, как выясняется, имеет важное значение для секреторной деятельности пепсиновых желез при нормальных условиях. Обе методики поэтому пригодны лишь для выяснения некоторых вопросов, относящихся работе пепсиновых желез. Так, например, первая методика особенно наглядно демонстрирует важное значение, которое акт еды имеет для работы пепсиновых желез, и делает возможным полный анализ этого факта. При разработке совершенной методики, которая позволяла бы исследовать всю работу пепсиновых желее, исходным пунктом служил принцип, впервые примененный Тири для изучения кишечной секреции, - принцип изолирования одной части сецернирующей поверхности. Впервые изолирование части фундального отдела желудка было осуществлено с полным успехом Гейденгайном [3].
На большой кривизне желудка собаки двумя соединяющимися друг с другом вверху разрезами, наложенными через переднюю и заднюю стенки желудка, вырезался ромбовидный кусок желудочной стенки, который питался сосудами сальника. Зашитый по линиям разреза кусок вместе с оставленным наверху отверстием вшивался в отверстие брюшной стенки. Таким путем получался маленький желудочек, совершенно изолированный от остального желудка. Из этого маленького желудочка поступал наружу чистый желудочный сок, когда в остальном пищеварительном канале продолжалось пищеварение.
Таким образом было Выполнено главное условие: точное наблюдение секреции происходило при соприкосновении пищи со всеми отрезками пищеварительного канала. Но вполне ли соответствовала наличная секреция маленького желудочка работе остального, оставшегося в нормальных условиях желудка? При гейденгайновском маленьком желудочке этого соответствия не существовало. И было ясно почему. При гейденгайновской операции поперечными разрезами перерезались все или почти все идущие к маленькому желудочку волокна блуждающего нерва. Ho n. vagus является бесспорно секреторным нервом пепсиновых желез. Стало быть, надо было изолировать маленький желудочек с сохранением всех его нервных связей. Это было достигнуто маленькой вариацией в операционной методике (Павлов [1]). Так как оперированный по этому способу маленький желудочек является в настоящее время наилучшим объектов изучения нормальной работы пепсиновых желез, то необходимо дать здесь подробное описание операции такого желудочка.
Вдоль белой линии, начиная от мечевидного отростка, проводят разрез длиною 8-10 см. Через него вытягивают почти весь желудок. На желудок накладывается временная лигатура из тонкой резиновой трубки, сначала на границе между фундальной и пилорической частью, причем на кривизнах трубка проводится между желудочной стенкой и сосудами сальника. На расстоянии 15 см в направлении к cardia таким же образом накладывается вторая, точно такая же лигатура на фундальную часть. На расстоянии 2-3 см от первой лигатуры на передней стенке фундальной части, начиная у линии прикрепления сальника к большой кривизне, проводят разрез, длиною в 7-8 см, который полого поднимается к малой кривизне, главным же образом в сторону cardia, т. е. в направлении продольной оси дна желудка. Разрез проводится лишь через серозный и мышечный слои, так что слизистая выпячивается через разрез. На ней выступают довольно крупные кровеносные сосуды. Каждый из этих сосудов перевязывается в двух местах по краям серозно-мышечного разреза. Такой же приблизительно симметричный разрез проводится по задней стенке желудка. Затем на передней стенке, между попарно надоженными лигатурами, накладывается небольшой разрез через слизистую. Через это отверстие лежащий между упомянутыми резиновыми лигатурами отрезок желудочной полости подвергается дезинфекции. Дезинфекция производится 0.5 %-м раствором соляной кислоты, которую 2-3 раза вливают внутрь желудка и вновь удаляют при помощи марлевых тампонов. Далее перерезается вся слизистая по всей длине разреза, сделанного ранее в серозной и мышечной оболочках, что происходит теперь почти без всякого кровотечения. Таким способом получают довольно большой удлиненный кусок желудочной стенки, имеющий форму треугольника, соединенного у своего основания с остальным желудком и у которого годаря направлению разреза - сохранены все волокна блуждающего нерва.
Теперь нужно этот кусок отделить от остального желудка. Этого достигают совершенно особым путем, образуя у основания нашего лоскута перегородку исключительно за счет слизистой. По концам линии основания на серозно-мышечный слой накладывают по плану. Натягивая несколько этот слой при помощи пэанов, быстрым и поверхностным движением перерезают лишь слизистую вдоль всего основания. Безошибочная удача в наложении разреза облегчается, с одной стороне, тем, что ассистент двумя пинцетами слегка приподнимает и натягивает слизистую, а с другой стороны, тем, что оператор нащупывает линию разреза по серозе. Накладывая небольшой марлевый жгутик на проведенный разрез, немного унимают кровотечение. Затем, приподнимая пинцетами оба края перерезанной слизистой и таким образом натягивая подслизистую, быстро еще раз проводят скальпелем по линии разреза. При этом, следовательно, перерезают подслизистую, причем палец оператора опять регулирует глубину разреза с поверхности серозы. Этим вторым разрезом слизистая оказывается сразу отпрепарованной на 0.75 см в обе стороны. Затем тщательно перевязывают почереди сосуды подслизистой, которые вызывают сильное кровотечение. Этим заканчивается выкраивание маленького желудочка.
Теперь начинается шитье. Здесь наибольшее внимание должно быть сосредоточено на швах в перегородке, ибо их расположение определяет судьбу маленького желудочка. Описанным далее распределением швов полностью гарантируется прочность перегородки, так как при этом из обоих краев слизистой образуются два полных свода: один - со стороны большого, другой - со стороны маленького желудка. Подслизистая каждого края слизистой сшивается с серозным слоем краев первого разреза и именно следующим образом. Первый шов накладывается на подслизистую в самом конце соответственного края и на серозный слой (вместе с мышечным) - у соответствующего из двух вышеупомянутых пэанов, и именно на той его стороне, на которой находится подлежащий пришитию край слизистой. Следующий шов на подслизистую накладывается ближе к середина кромки слизистой, на серозный слой - дальше от пэанов, вдоль края в направлении соответственной полости. Швы (в количестве пяти-семи) накладывают почти до середины края слизистой. Они занимают на краю серозно-мышечного слоя расстояние в 1-1.5 см. То же самое проделывается на каждой половине обоих краев перерезанной слизистой.
Два образованных таким образом из слизистой свода представляют собой перегородку между полостями большого и маленького желудка. Остается только закрыть обе полости. Выгоднее всего наложить первый шов на месте вышеупомянутых пэанов, т. е. соединить края серозно-мышечного моста между обеими полостями. Далее следуют два шва по обе стороны первого, причем в середине между серозно-мышечными полосами немного захватывается лигатурой также и куполообразно изогнутая подсерозная, чем она еще лучше фиксируется. Таких швов накладывается по 2-3 с каждой стороны первых. Наконец накладываются обыкновенные швы на большой желудок до тех пор, пока он не закроется; на маленьком их продолжают накладывать, пока не останется в конце небольшое отверстие, края которого подшиваются к отверстию, оставшемуся в брюшной стенке. Как только края этого отверстия совершенно заживут, приступают к собиранию сока, причем в полость маленького желудочка вводится резиновая трубка с боковыми отверстиями в той ее части, которая входит в маленький желудочек.
Теперь, раньше чем пользоваться маленьким желудочком для изучения нормальной работы пепсиновых желез, необходимо убедиться в его пригодности. С этой целью нужно сравнить его работу с работой большого желудка. Ввиду этого с операцией маленького желудка соединяют наложение обыкновенной желудочной фистулы на большом желудке. Сравнение состоит в разнообразных опытах. Сначала открывают фистулы большого желудка и дают собаке несколько кусочков мяса, которые скоро снова выпадают через фистулу. Начинается вызванная актом еды секреция. Если при энергичной секреции в большом желудке в маленьком не воспоследует никакой сексеции, то он должен считаться негодным. Значит, при операции повреждено очень много волокон блуждающего нерва. Если же, наоборот, секреция налицо, то нужно установить соотношение, существующее между количеством сока, выделенного из обеих полостей в течение определенной единицы времени или в течение всего хода секреции, и перейти к следующему испытанию. Оно заключается в том, что, без того чтобы собака это видела, в большой желудок вводится молотое мясо или раствор либиховского мясного экстракта. Когда секреция, определяемая по маленькому делудочку, достигнет значительной степени, открывают фистулу большого желудка и опоражнивают его. Выделяющийся несколько времени затем желудочный сок собирается, и его количество сравнивается с количеством сока, выделившегося из маленького желудочка за это же время. При абсолютно удавшимся маленьком желудочке соотношение количеств при обоих приведенных испытаниях должно быть одинаковым. Часть при первом испытании, т. е. испытании актом еды, отношение склоняется в пользу большого желудка; это обстоятельство такое указывает на значительное повреждение волокон блуждающего нерва маленького желудочка. В настоящее время вполне понятиен тот факт, что маленький желудочек может быть точным отражением большого, хотя в него не попадает никакой еды. Секреция пепсиновых желез возбуждается двоякого рода механизмами: во-первых, несомненным нервным механизмом при акте еды, без соприкосновения еды со слизистой желудка, и, во-вторых, химическим механизмом, в силу соприкосновения пищи с пилорической частью желудка.
Итак, если нервные связи маленького желудочка целы, то его пепсиновые железы находятся в точно таких же условиях возбуждения, как железы большого желудка.
Хотя маленький желудочек, как он только что описан, почти полностью отвечает идеалу, все же с чисто технической точки зрения ему присущ значительный недостаток: его отверстие имеет склонность быстро расширяться под действием вытекаюшего сока, который сильно разъедает окружность отверстия. Для избежания этого до сих пор применялось лишь одно верное средство: пока продолжается секреция из маленького желудочка, сок собирается при помощи вышеупомянутой трубочки.
Эпп [3] рекомендует для той же цели, для которой служит маленький желудочек, операцию, которую он проделывает над свиным желудком и которую он предпочитает маленькому желудочку, однако, как мы дальше увидим, без достаточного основания. Этот автор перерезает, при полном сохранении блуждающего нерва, пищевод перед cardia и соединяет его с двенадцатиперстной кишкой. Этим путем восстанавливается непрерывность пищеварительного канала. На другом конце желудок отрезается от привратника. Изолированный желудок зашивается с обоих концов; пилорическая часть также зашивается. На желудок накладывается обыкновенная фистула. Работа изолированного таким путем желудка совершеннее и ближе подходит к нормальным условиям, чем работа желудочка по Фремону, ибо у первого, кроме нервного механизма, остается до известной степени в силе и химический механизм. Но все же он не может совершенно безукоризненно служить целям изучения нормального типа секреции. Нормальный ход секреции определяется нормальным переходом пищевых масс вместе с желудочным соком из желудка; этот переход регулируется работой привратниковой части и пилорического сфинктера; при данной же методике Эппа эта работа исключена. Автор обещает, что он вошьет нижний конец пищевода в привратниковую часть; даже и тогда, когда пища будет переходить изо рта непосредственно в эту часть желудка, не приходится рассчитывать на нормальный переход ее в кишечник ни с механической, ни с химической точки зрения. Что же касается относительной трудности сравниваемых операций, то вряд ли преимущество окажется на стороне операции Эппа. При известном хирургическом навыке, - он нужен в одинаковой степени для обеих операции, - маленький желудочек удается всегда и при испытании почти всегда оказывается совершенно пригодным.
Ввиду анатомических и физиологических особенностей пиларической части желудка, его необходимо изолировать в целях точного исследования его секрета и его секреторной работы. Изолирование происходит двояким образом: с сохранением и без сохранения волокон блуждающего нерва. В последнем случае изолирование достигается просто двумя поперечными разрезами; один из них накладывается на место привратника, а другой отступя на 5 см вдоль малой кривизны и на 6 см вдоль большой кривизны влево от первого разреза. Fundus сшивается с duodenum, и таким образом восстанавливается непрерывность пищеварительного канала. Из пилорической же части образуют слепой мешок, который вшивается в отверстие брюшной раны (Гейденгайн [67 [6]). Акерман зашивает изолированную часть с обоих концов и, отпрепаровав сальник на большой кривизне, делает фистульное отверстие в середине этой кривизны.
Изолирование с сохранением волокон блуждающего нерва производится также двумя различными способами. Изолируется либо весь пилорический отдел, либо только его часть. По первой методике (Крестев [*j) пилорический отдел отделяется от фундального разрезами, которые проникают через всю толщу желудочной стенки; лишь наверху на малой кривизне оставляют состоящий из serosa и muscularis мост, шириной в 3 см, соединяющий пилорическую часть с фундальной. Перегородку на этом месте делают лишь за счет слизистой. Двенадцатиперстная кишка отрезается и вшивается в fundus, или накладывают гастроэнтеростомоз. По второй методике (Шемякин [9] [3]) с пилорическим отделом поступают так же, как это описано для изолирования участка фундальной части с сохранением волокон блуждающего нерва, причем изолируемый участок пилорического отдела берут либо из большой, либо из малой кривизны. Происходящее при этом сужение привратниковой части не влечет за собой каких-либо заметных нарушений перехода желудочного содержимого в кишечник. Впрочем, надо заметить, что проведен ные до сих пор опыты и наблюдения не установили какой-либо значительной разницы в работе пилорических желез, была ли пилорическая часть изолирована с сохранением или без сохранения волокон блуждающего нерва.
При решении некоторых вопросов оказывается необходимой обыкновенная фистула, т. е. боковое отверстие в привратниковой части. Она накладывается либо так же, как на fundus, либо по особому способу, который будет описан по поводу фистул желчного пузыря и кишечника.
При изучении желез вообще и специально пепсиновых желез всплывает вопрос: с какой части поверхности пищеварительного канала происходит возбуждение соответственной секреции? Для решения этого вопроса необходимы некоторые дополнительные операции. Чтобы дать обоснованный ответ на вопрос о том, с какой поверхности химические возбудители оказывают свое воздействие на пепсиновые железы, необходимо проделать довольно сложную комбинацию операций в одном и том же животном, которые нужно проводить в определенной последовагельности. Прежде всего делается обыкновенный маленький желудочек фундальной части (Соколов [107] [10]). При данной комбинации разрез этой операции проводится слева вдоль наружного края m. rectus abdominis, начиная от края ребра. Пользуясь тем же разрезом, на большом желудке накладывают обычную желудочную фистулу. Фистульная трубка укрепляется в верхнем конце раневого разреза, отверстие маленького желудочка в нижнем его конце. Нужно позаботиться, чтобы кожа между обоими отверстиями была тщательно зашита, чтобы между ними образовался прочный кожный мост. Затем на двенадцатиперстную кишку накладывается боковая фистула с металлической канюлей. Кожный разрез для этой операции проводится справа у внешнего края m. rectus abdominis. О методике, по которой осуществляется эта фистула, будет подробно сказано ниже. Как только оба фистульных отверстия с канюлями обрастут прочной рубцовой тканью, через отверстие вдоль средней линии вытягивают желудок в месте перехода его в двенадцатиперстную кишку и делают перегородку между желудком и двенадцатиперстной кишкой. Эту перегородку образуют, либо проводя сплошной поперечный разрез, - стало быть, из всех слоев, либо только из слизистой. В последнем случае делают сначала продольный разрез через серозный и мышечный слои, кругаобразно отпрепаровывают слизистую перерезают поперек только ее. Перерезанные концы, так же как продольный разрез, зашиваются. После этой операции возникает крайне трудная задача кормления оперированной таким образом собаки. Лучше всего эта задача разрешается наружным гастроэнтеростомозом, т. е. желудочная канюля соединяется с дуоденальной достаточно широкими трубками из стекла или металла и из резины. Пищу собака получает лишь в размельченном виде. Переход пищи из желудка в кишечник происходит в общем довольно легко. Лишь временами, пока пища еще недостаточно разжижена желудочным соком, возникает необходимость протолкнуть рукой застрявшие в резиновой трубке пищевые массы. Дабы избежать повреждения животным этого соединения желудка с кишечником, приходится во время пищеварения держать собаку в станке. Когда же собаку спускают, канюли либо разъединяются, и каждая из них затыкается пробкой, или их соединяют тонкой резиновой трубкой, которая почти прилегает к брюшной стенке и мало заметна для собаки. Если собака не трогает эту трубку, не вырывает ее, выгоднее оставить собаку с этой трубкой, чем совершенно разъединить канюли. По этой трубке накапливающаяся в желудке жидкость непрерывно переходит постепенно в кишечник; она состоит из выделяющегося после пищеварения желудочного сока, из слюны, психического сока и накапливается в течение ночи в количестве многих сотен кубических сантиметров.
На такой собаке можно совершенно точно сравнивать влияние химических возбудителей пепсиновых желез то со слизистой кишечника, то с желудка.
Идя далее, можно установить разницу в действии такого возбудителя с разных отрезков желудка, т. е. с поверхности пилорического или фундального отдела. этой целью поступают вначале точно так же, как в предыдущем случае, но при последней операции поперечный разрез на желудке накладывают не на его границе с двенадцатиперстной кишкой, а на границе между фундальной и пилорической частью. При этом особое внимание обращают на то, чтобы каждый из разделенных отрезков желудка не содержал каких-либо частей другого. Это достигается широким удалением промежуточной области (В. Гросс [1]).
Как вивисекции, так и хирургические операции производились не только для получения желудочного сока и для исследования нормальной работы желудочных желез, но и для исследования иннервации этих желез. Здесь дело идет главным образом перерезке или раздражении блуждающего нерва и n. splanchnicus. Так как операции на этих нервах производились для исследования кровообращения и будут описаны в соответственной главе, то я приведу здесь только те варианты, которые применялись или могли быть применены специально для исследования иннервации желудочных и других пищеварительных желез. Сюда относятся разнообразные операции перерезки блуждающего нерва на различных уровнях. Простую перерезку nn. vagi на середине шеи оказалось возможным применить в качестве убедительного опыта для доказательства секреторного значения nn. vagi для пепсиновых желез только тогда, когда был найден способ сохранять собак в совершенно здоровом состоянии на неопределенно долгое время. При первом опыте (Павлов и Шумова-Симановская), доказавшем участие сn. vagi в секреторном эффекте мнимого кормления, применялась операция Генцмера [12] [12]. Она состоит в том, что n. vagus перерезают слева, как обычно - по середине шеи, справа же - под местом ответвления n. laryngeus inferior в большинства сердечных ветвей блуждающего нерва. Эта последняя операция проделывается следующим образом. Кожный разрез накладывается внизу шеи от верхнего края грудной клетки на 5-6м кверху к внешнему краю m. sternocleidomastoideus. После кожи перерезается фасция, разрывается соединительная ткань; при этом обнажаются снаружи vv. jugularis и subclavia, а снутри от щитовидного хряща и n. vagosympathicus с прилежащими art. carotides. Далее изолируют в глубине n. vagus под art. subclavia dextra, исключительно при помощи препаровальной иглы, причем n. vagus, который загибается вниз, поднимается на крючок, при положении животного на спине, и перерезается на 1-1.5 5 см ниже art. subclavia. Выгоднее перерезать сначала n. vagus на правой стороне, как описано, затем через две недели слева, на середине шеи, после того как в промежутке все оставшиеся ветки правого блуждаюшего нерва оправились после неизбежной при операции травмы. Такой перерезкой в два темпа достигается то, что животное после двусторонней перерезки nn. vagi не впадает в депрессивное состояние, наступающее после простой перерезки по середине шеи, ибо таким путем избегаются важные функциональные нарушения со стороны гортани и сердца.
На оперированной таким образом собаке, на которой прежде проделаны операции гастро- и эзофаготомии, ставится также опыт с раздражением блуждающего нерва как секреторного нерва пепсиновых желез. Для этой цели при перерезке левого блуждающего нерва (при последней операции) его периферический конец слегка отпрепаровывается и перевязывается шелковой ниткой, конец которой проводится через зашитую рану наружу. На следующий день у животного, спокойно стоящего в станке, снимают 1--2 кожных шва и с помощью шелковой нитки легко вытягивают наружу нерв, что не вызывает никакой реакции со стороны животного. Нерв раздражается индукционными ударами, посылаемыми каждую секунду в нерв. Таким образом нерв раздражается без наркотизации животного и без того, чтобы оно испытывало боль перед опытом или во время него (Павлов и Шумова-Симановская[15]).
Общее самочувствие животного, разумеется, еще меньше нарушается перерезкой nn. vagi на более низких уровнях, т. е. в грудной полости, под местом ответвления легочных ветвей, и в брюшной полости, чем только что описанной перерезкой nn. vagi на шее, хотя первые операции технически гораздо сложнее.
Операция перерезки в грудной полости производится двумя способами: по способу Клод Бернара"] и по способу Креля [15] (лаборатория Людвига). По Клоду Бернару, на соответственном месте грудной стенки разрезается кожа; разрезанная кожа отодвигается, и здесь же перерезается фасция и mm. intercostales. За Затем проникают пальцем в грудную полость, но так, чтобы туда не попал воздух. По пальцу вводится изогнутый крючком нож, лезвие которого находится на вогнутой стороне. Под контролем пальца перерезают оба nn. vagi, так же осторожно, чтобы не впустить воздух, вынимают из раны палец и нож и затем сдвигают кожу на ее прежнее место. Подобная операция нелегка, ее трудно правильно выполнить, и она могла удаваться только такому превосходному оператору, каким был Клод Бернар.
В лаборатории Людвига эта операция производилась гораздо более сложным способом, но зато полным ручательством. Сперва изготовляются резиновые колпаки с двойными стенками, широкое отверстие которых надевается на морду собаки, а узкое отверстие соединяется резиновой трубкой с дыхательным аппаратом. Чтобы колпак герметически прилетал к морде собаки, пространство между двойными стенками сильно надувается воздухом. Во время вскрытия грудной полости делается искусственное дыхание. На левой стороне вблизи позвоночника между шестым и седьмым ребрами делается надрез, который проникает в грудную полость. Ребра раздвигают как можно шире, вытягивают пищевод и перерезают на нем nn. vagi. Затем сильным надуванием легких выдавливают воздух из грудной полости и тщательно зашивают рану. /Животные прекрасно переносят эту операцию. Эти обе последние операции применились собственно для решения вопроса о причинах смерти после перерезки nn. vagi, но они вполне годятся и для опытов над иннервацией пепсиновых желез и других пищеварительных желез блуждающими нервами.
Операция перерезки nn. vagi в грудной полости имеет также несколько вариантов. Делается довольно длинной разрез вдоль средней линии, начинающийся у processus xiphoideus (Юргенсно [16] желудка отодвигают кпозвоночнику и вниз и, следуя затем по легко заметным ветвям nn. vagi, достигают без особых трудностей до обоих нервных стволов. Если хотят их перерезать возможно выше, то их присутствие нужно определить нащупыванием, для чего cardia на несколько секунд оттягивают книзу. Нервы отпрепаровывают от окружающей их ткани и перерезают или резецируют. Главным условием успеха операции, кроме самой тщательной чистоты, является оперирование в глубине раны in situ, ибо притягивание cardia к краю брюшной раны вызывает часто смерть собаки через 2-3 дня, вероятно вследствие шока. По Шиффу [17], перерезка нервов производится следующим образом. B regio hypochondrica sinistra накладывается разрез, длиною в 2-3 см, через который добираются до субдиафрагмального отдела пищевода; тогда под него подводят зонд и вытягивают наружу. Затем перерезают все видимые на поверхности пищевода нервы или резецируют их, переворачивают пищевод так, чтобы видны были задняя и боковые поверхности, и перерезают серозную оболочку циркулярно, не повреждая мышечного слоя.
Из этих двух способов следует предпочесть первый, во-первых, потому, что он гарантирует полноту перерезки, т. е. при этом перерезаются все брюшные ветви nn. vagi, и верхние окончания нервов оказываются в грудной клетке на 1-2 см выше диагфрагмы. Во-вторых, этот способ дает возможность оперировать in situ, что, как уже указано выше, имеет то преимущество, что собаки не погибают от операции.
Фруэн и Позерский [187 [18] перерезают nn. vagi из брюшной полости над диафрагмой следующим образом. Вдоль linea mediana делают разрез, длиною в 10 см, начинающийся у грудины, захватывают между двумя браншамизогнутого кишечного пинцета lig. gastrophrenicum и диафрагму за пищеводом и перерезают соединительнотканные тяжи, образующие предназначенное для пищевода отверстие в грудобрюшной преграды. Таким путем устраняется опасность пнесимоторакса от прободения диаФрагмы при наложении пинцета, соединяющего обе стороны грудобрюшной преграды - правую и левую. Затем вытягивают пищевод как можно дальше под грудобрюшную преграду и берут его на крепкую шелковину. Хорошо видамые теперь nn. vagi перерезаются таким образом гораздо выше грудобрюшной преграды. Зашив отверстие в грудобрюшной преграды, снимают пинцет и закрывают брюшную рану обычным способом.
Все эти способы перерезки nn. vagi все же оставили нерешенной одну методическую задачу, а именно задачу точного сравнения, во всех подробностях, деятельности пепсиновых желез при наличии секреторных волокон блуждающего нерва и без них. Однако в этом последнем случае крайне важно, чтобы не наступали никакие побочные нарушения секреторной работы пепсиновых желез, вызывающие недостаточную обработку пищи в желудке и ослабление его моторной функции и наступающие при всех вышеприведенных способах. Подобное сравнение может быть сделано лишь при изолированных маленьких желудочках, которые оперируются по Гейденгайну (без ветвей блуждающего нерва) или по Павлову (с сохранением иннервации блуждающего нерва). Но и в этом случае сравнение не могло бы быть совершенно бесспорным и простым, так как относилось бы к разным собакам и к различным маленьким желудочком. Требуемое сравнение выполнено совершенно точно Орбели [18] следующим образом. Были изготовленасколько возможно, удачные маленькие желудочки с сохранением иннервации. После того как их деятельность была разносторонне обследована и оказалась закономерной и стационарной, вскрывалась брюшная полость и перерезался серозно-мышечный мост между большим и маленьким желудком. При этом без всякого затруднения отделялись друг от друга оба слоя слизистой, образующие перегородку между двумя желудками. При этой операции оба эти слоя могли оставаться совершенно неповрежденными и отделение маленького желудка от большого протекало почти без потери единой капли крови и без какой-либо перевязки кровеносных сосудов. Единственная невыгода только что описанного способа состоит в том, что некоторые, действительно едва заметные, волокна блуждающего нерва остаются неперерезанными. Очевидно, эти редкие и скудные волокна проникают в маленький желудочек со стороны мезентерия. Их наличие обнаруживается незначительным возбуждением пепсиновых желез при еде.
В заключение заслуживают краткого упоминания некоторые пункты операционной методики, которые имеют целью раздражение блуждающего нерва как секреторного нерва пепсиновых желез в опытах ex tempore. Так как при какдом чувствительном раздражении работа пепсиновых желез сильно затормаживается, а при сильном раздражении это торможение становится упорным и длительным, то в случае вивисекции нужно начинать с уничтожения чувствительного раздражения, т. е. с перерезки спинного мозга под продолговатым.
Это можно выполнить различными способами: 1) можно применять очень кратковременный хлороформный наркоз, которого как раз хватает для перерезки спинного мозга и для введения трахеотомической трубки; 2) можно начинать с перерезки спинного мозга (в течение нескольких секунд) и сразу же вводить канюлю в трахею без всякого предшествующего наркоза. В последнем случае необходимо, чтобы голова привязанной за ноги к столу собаки держалась крепко и правильно и чтобы оператор мог производить перерезку точно и быстро двумятремя разрезами.
Второй пункт методики затрагивает необходимость перевязывать пищевод и pylorus при получении желудочного сока, чтобы предупредить поступление в желудок другого рода жидкостей изо рта и из кишечника (Ушаков [201] 7).
Операция эзофаготомии также очень проста и при соблюдении известных условий удается всегда. Разрез, длиною в 10 см и больше, смотря по собаке, проводится по левой стороне шеи по внутреннему краю m. sternocleidomastoideus, начинаясь у внутреннего края щитовидного хряща разрезается, и пространство между m. sternocleidomastoideus и лежащими на трахее мышцами отпрепаровывается. Под трахеей, по середине раны, пищевод захватывается пэаном, отпрепаровывается по всей своей окружности на 2 3 см в длину и полностью перерезается поперек. Быстро вытянув концы закрыв их планами или тампонами, нужно теперь позаботиться том, чтобы слюна из пищевода совершенно не попадала в образующуюся глубокую рану. Концы пищевода подшиваются к раневым углам, причем иглой нужно обкалывать не только кожу, но мышцы, ограничивающие рану. Этим достигается то, что концы пищевода не втягиваются внутрь, но навсегда остаются на поверхности. Середина раны между вшитыми в углы раны концами пищевода тщательно зашивается как послойно в глубине, так на кожной поверхности. Таким образом между концами пищевода получается порядочный промежуток, образованный из сросшейся кожи, что исключает возможность того, чтобы даже самые незначительные количества слизи или составных частей пищи попадали из верхнего конца пищевода в нижний. Главное правило операции, гарантирующее правильное и быстрое заживление, заключается в том, чтобы операцию производили абсолютно чисто, избегая загрязнения глубокой раны, где нагноение, раз начавшись, распространяется быстро и интенсивно. По той же причине и после операции нужно ежедневно обмывать рану антисептической жидкостью; лучше даже повторять это несколько раз в день, так как рана постоянно очень сильно загрязняется снаружи слюной. Наложение желудочной фистулы должно предшествовать эзофаготомии на 3-4 недели, т. е. эзофаготомия проделывается после того, как образовалась рубцовая ткань, крепко обхватывающая канюлю.
Сперва, до полного заживления раны, собаку кормят через желудочную фистулу. Твердая пища вкладывается в желудок кусочками через открытую фистулу; жидкости вливаются, причем канюля закрывается простым вентилем, вставленным в просверленную пробку. В дальнейшем вся еда - жидкость вместе с размельченной твердой пищей - удобно и быстро вливается в нижний конец пищевода через воронку с толстой резиновой трубкой, при стоячем положении собаки.
Принимая во внимание потерю слюны и особенно часто повторяемое собирание желудочного сока, необходимо вводить собаке много жидкости. Если нужно, можно таким способом без вреда для животного ежедневно собирать желудочный сок в течение очень долгого времени. Получаемое каждый раз количество сока может достигать у больших собак 1.5 л. В то время как у нее описанным способом собирают сок, собаку можно оставлять без присмотра, если ставить перед ней на подставке довольно широкую чашку с кусочками какой-нибудь пищи так, чтобы собака могла сама до нее дотянуться. Проглоченный кусок каждый раз падает обратно в чашку. Под желудочную фистулу подвешивается довольно широкий, толстостенный суживающийся книзу в трубку цилиндр. В том месте, где цилиндр суживается, кладется стеклянная вата или лигнин, чтобы задерживать слизь. На суженный конец надевается довольно длинная резиновая трубка, другой конец которой соединяется несколько в стороне от животного, либо прямо, либо при помощи стеклянного или каучукового крючка с толстостенным, шарообразно расширяющимся книзу сосудом; в этом последнем и собирается весь чистый желудочный сок. У некоторых животных иногда в желудок забрасывается кишечное содержимое, что сразу можно узнать по окраске желудочного содержимого желчью. В подобном случае желудок очищается тщательным промыванием водой, и тогда собирание желудочного сока может быть возобновлено. Само собой разумеется, в день собирания желудочного сока собаку с утра не кормят. Несмотря на часто повторяемое обильное собирание сока, эти собаки живут в полном здоровии и остаются в течение многих лет совершенно точно работающими фабриками желудочного сока.
Для получения чистого желудочного сока служит также операция Фремона [2], которая состоит в полном изолировании всего желудка от остального пищеваритель поперечные разрезы как на границе между пищеводом и желудком, так и между желудком и двенадцатиперстной кишкой. Нижний конец пищевода сшивается с двенадцатиперстной кишкой, и таким образом восстанавливается непрерывность пищеварительного канала. Зашитый же с двух концов желудок опоражнивается через обыкновенную желудочную фистулу. Представляя решительное преимущество перед методикой одновременных желудочной и пищеводной фистул в том отношении, что забрасывание кишечного содержимого в желудок здесь совершенно исключено, эта методика все же сильно ей уступает; это касается как трудности самой операции (ввиду необходимости сохранять совершенно неповрежденными nn. vagi), трудно сохранить на более продолжительное время здоровыми и живыми оперированных таким образом собак.
Хотя обе описанные методики вполне пригодны для получения чистого желудочного сока, все же они не могут претендовать на то, чтобы годиться и для полного анализа нормальной работы пепсиновых желез. При обеих методиках не происходит соприкосновения пиши со стенками желудка, тогда как именно это соприкосновение, как выясняется, имеет важное значение для секреторной деятельности пепсиновых желез при нормальных условиях. Обе методики поэтому пригодны лишь для выяснения некоторых вопросов, относящихся работе пепсиновых желез. Так, например, первая методика особенно наглядно демонстрирует важное значение, которое акт еды имеет для работы пепсиновых желез, и делает возможным полный анализ этого факта. При разработке совершенной методики, которая позволяла бы исследовать всю работу пепсиновых желее, исходным пунктом служил принцип, впервые примененный Тири для изучения кишечной секреции, - принцип изолирования одной части сецернирующей поверхности. Впервые изолирование части фундального отдела желудка было осуществлено с полным успехом Гейденгайном [3].
На большой кривизне желудка собаки двумя соединяющимися друг с другом вверху разрезами, наложенными через переднюю и заднюю стенки желудка, вырезался ромбовидный кусок желудочной стенки, который питался сосудами сальника. Зашитый по линиям разреза кусок вместе с оставленным наверху отверстием вшивался в отверстие брюшной стенки. Таким путем получался маленький желудочек, совершенно изолированный от остального желудка. Из этого маленького желудочка поступал наружу чистый желудочный сок, когда в остальном пищеварительном канале продолжалось пищеварение.
Таким образом было Выполнено главное условие: точное наблюдение секреции происходило при соприкосновении пищи со всеми отрезками пищеварительного канала. Но вполне ли соответствовала наличная секреция маленького желудочка работе остального, оставшегося в нормальных условиях желудка? При гейденгайновском маленьком желудочке этого соответствия не существовало. И было ясно почему. При гейденгайновской операции поперечными разрезами перерезались все или почти все идущие к маленькому желудочку волокна блуждающего нерва. Ho n. vagus является бесспорно секреторным нервом пепсиновых желез. Стало быть, надо было изолировать маленький желудочек с сохранением всех его нервных связей. Это было достигнуто маленькой вариацией в операционной методике (Павлов [1]). Так как оперированный по этому способу маленький желудочек является в настоящее время наилучшим объектов изучения нормальной работы пепсиновых желез, то необходимо дать здесь подробное описание операции такого желудочка.
Вдоль белой линии, начиная от мечевидного отростка, проводят разрез длиною 8-10 см. Через него вытягивают почти весь желудок. На желудок накладывается временная лигатура из тонкой резиновой трубки, сначала на границе между фундальной и пилорической частью, причем на кривизнах трубка проводится между желудочной стенкой и сосудами сальника. На расстоянии 15 см в направлении к cardia таким же образом накладывается вторая, точно такая же лигатура на фундальную часть. На расстоянии 2-3 см от первой лигатуры на передней стенке фундальной части, начиная у линии прикрепления сальника к большой кривизне, проводят разрез, длиною в 7-8 см, который полого поднимается к малой кривизне, главным же образом в сторону cardia, т. е. в направлении продольной оси дна желудка. Разрез проводится лишь через серозный и мышечный слои, так что слизистая выпячивается через разрез. На ней выступают довольно крупные кровеносные сосуды. Каждый из этих сосудов перевязывается в двух местах по краям серозно-мышечного разреза. Такой же приблизительно симметричный разрез проводится по задней стенке желудка. Затем на передней стенке, между попарно надоженными лигатурами, накладывается небольшой разрез через слизистую. Через это отверстие лежащий между упомянутыми резиновыми лигатурами отрезок желудочной полости подвергается дезинфекции. Дезинфекция производится 0.5 %-м раствором соляной кислоты, которую 2-3 раза вливают внутрь желудка и вновь удаляют при помощи марлевых тампонов. Далее перерезается вся слизистая по всей длине разреза, сделанного ранее в серозной и мышечной оболочках, что происходит теперь почти без всякого кровотечения. Таким способом получают довольно большой удлиненный кусок желудочной стенки, имеющий форму треугольника, соединенного у своего основания с остальным желудком и у которого годаря направлению разреза - сохранены все волокна блуждающего нерва.
Теперь нужно этот кусок отделить от остального желудка. Этого достигают совершенно особым путем, образуя у основания нашего лоскута перегородку исключительно за счет слизистой. По концам линии основания на серозно-мышечный слой накладывают по плану. Натягивая несколько этот слой при помощи пэанов, быстрым и поверхностным движением перерезают лишь слизистую вдоль всего основания. Безошибочная удача в наложении разреза облегчается, с одной стороне, тем, что ассистент двумя пинцетами слегка приподнимает и натягивает слизистую, а с другой стороны, тем, что оператор нащупывает линию разреза по серозе. Накладывая небольшой марлевый жгутик на проведенный разрез, немного унимают кровотечение. Затем, приподнимая пинцетами оба края перерезанной слизистой и таким образом натягивая подслизистую, быстро еще раз проводят скальпелем по линии разреза. При этом, следовательно, перерезают подслизистую, причем палец оператора опять регулирует глубину разреза с поверхности серозы. Этим вторым разрезом слизистая оказывается сразу отпрепарованной на 0.75 см в обе стороны. Затем тщательно перевязывают почереди сосуды подслизистой, которые вызывают сильное кровотечение. Этим заканчивается выкраивание маленького желудочка.
Теперь начинается шитье. Здесь наибольшее внимание должно быть сосредоточено на швах в перегородке, ибо их расположение определяет судьбу маленького желудочка. Описанным далее распределением швов полностью гарантируется прочность перегородки, так как при этом из обоих краев слизистой образуются два полных свода: один - со стороны большого, другой - со стороны маленького желудка. Подслизистая каждого края слизистой сшивается с серозным слоем краев первого разреза и именно следующим образом. Первый шов накладывается на подслизистую в самом конце соответственного края и на серозный слой (вместе с мышечным) - у соответствующего из двух вышеупомянутых пэанов, и именно на той его стороне, на которой находится подлежащий пришитию край слизистой. Следующий шов на подслизистую накладывается ближе к середина кромки слизистой, на серозный слой - дальше от пэанов, вдоль края в направлении соответственной полости. Швы (в количестве пяти-семи) накладывают почти до середины края слизистой. Они занимают на краю серозно-мышечного слоя расстояние в 1-1.5 см. То же самое проделывается на каждой половине обоих краев перерезанной слизистой.
Два образованных таким образом из слизистой свода представляют собой перегородку между полостями большого и маленького желудка. Остается только закрыть обе полости. Выгоднее всего наложить первый шов на месте вышеупомянутых пэанов, т. е. соединить края серозно-мышечного моста между обеими полостями. Далее следуют два шва по обе стороны первого, причем в середине между серозно-мышечными полосами немного захватывается лигатурой также и куполообразно изогнутая подсерозная, чем она еще лучше фиксируется. Таких швов накладывается по 2-3 с каждой стороны первых. Наконец накладываются обыкновенные швы на большой желудок до тех пор, пока он не закроется; на маленьком их продолжают накладывать, пока не останется в конце небольшое отверстие, края которого подшиваются к отверстию, оставшемуся в брюшной стенке. Как только края этого отверстия совершенно заживут, приступают к собиранию сока, причем в полость маленького желудочка вводится резиновая трубка с боковыми отверстиями в той ее части, которая входит в маленький желудочек.
Теперь, раньше чем пользоваться маленьким желудочком для изучения нормальной работы пепсиновых желез, необходимо убедиться в его пригодности. С этой целью нужно сравнить его работу с работой большого желудка. Ввиду этого с операцией маленького желудка соединяют наложение обыкновенной желудочной фистулы на большом желудке. Сравнение состоит в разнообразных опытах. Сначала открывают фистулы большого желудка и дают собаке несколько кусочков мяса, которые скоро снова выпадают через фистулу. Начинается вызванная актом еды секреция. Если при энергичной секреции в большом желудке в маленьком не воспоследует никакой сексеции, то он должен считаться негодным. Значит, при операции повреждено очень много волокон блуждающего нерва. Если же, наоборот, секреция налицо, то нужно установить соотношение, существующее между количеством сока, выделенного из обеих полостей в течение определенной единицы времени или в течение всего хода секреции, и перейти к следующему испытанию. Оно заключается в том, что, без того чтобы собака это видела, в большой желудок вводится молотое мясо или раствор либиховского мясного экстракта. Когда секреция, определяемая по маленькому делудочку, достигнет значительной степени, открывают фистулу большого желудка и опоражнивают его. Выделяющийся несколько времени затем желудочный сок собирается, и его количество сравнивается с количеством сока, выделившегося из маленького желудочка за это же время. При абсолютно удавшимся маленьком желудочке соотношение количеств при обоих приведенных испытаниях должно быть одинаковым. Часть при первом испытании, т. е. испытании актом еды, отношение склоняется в пользу большого желудка; это обстоятельство такое указывает на значительное повреждение волокон блуждающего нерва маленького желудочка. В настоящее время вполне понятиен тот факт, что маленький желудочек может быть точным отражением большого, хотя в него не попадает никакой еды. Секреция пепсиновых желез возбуждается двоякого рода механизмами: во-первых, несомненным нервным механизмом при акте еды, без соприкосновения еды со слизистой желудка, и, во-вторых, химическим механизмом, в силу соприкосновения пищи с пилорической частью желудка.
Итак, если нервные связи маленького желудочка целы, то его пепсиновые железы находятся в точно таких же условиях возбуждения, как железы большого желудка.
Хотя маленький желудочек, как он только что описан, почти полностью отвечает идеалу, все же с чисто технической точки зрения ему присущ значительный недостаток: его отверстие имеет склонность быстро расширяться под действием вытекаюшего сока, который сильно разъедает окружность отверстия. Для избежания этого до сих пор применялось лишь одно верное средство: пока продолжается секреция из маленького желудочка, сок собирается при помощи вышеупомянутой трубочки.
Эпп [3] рекомендует для той же цели, для которой служит маленький желудочек, операцию, которую он проделывает над свиным желудком и которую он предпочитает маленькому желудочку, однако, как мы дальше увидим, без достаточного основания. Этот автор перерезает, при полном сохранении блуждающего нерва, пищевод перед cardia и соединяет его с двенадцатиперстной кишкой. Этим путем восстанавливается непрерывность пищеварительного канала. На другом конце желудок отрезается от привратника. Изолированный желудок зашивается с обоих концов; пилорическая часть также зашивается. На желудок накладывается обыкновенная фистула. Работа изолированного таким путем желудка совершеннее и ближе подходит к нормальным условиям, чем работа желудочка по Фремону, ибо у первого, кроме нервного механизма, остается до известной степени в силе и химический механизм. Но все же он не может совершенно безукоризненно служить целям изучения нормального типа секреции. Нормальный ход секреции определяется нормальным переходом пищевых масс вместе с желудочным соком из желудка; этот переход регулируется работой привратниковой части и пилорического сфинктера; при данной же методике Эппа эта работа исключена. Автор обещает, что он вошьет нижний конец пищевода в привратниковую часть; даже и тогда, когда пища будет переходить изо рта непосредственно в эту часть желудка, не приходится рассчитывать на нормальный переход ее в кишечник ни с механической, ни с химической точки зрения. Что же касается относительной трудности сравниваемых операций, то вряд ли преимущество окажется на стороне операции Эппа. При известном хирургическом навыке, - он нужен в одинаковой степени для обеих операции, - маленький желудочек удается всегда и при испытании почти всегда оказывается совершенно пригодным.
Ввиду анатомических и физиологических особенностей пиларической части желудка, его необходимо изолировать в целях точного исследования его секрета и его секреторной работы. Изолирование происходит двояким образом: с сохранением и без сохранения волокон блуждающего нерва. В последнем случае изолирование достигается просто двумя поперечными разрезами; один из них накладывается на место привратника, а другой отступя на 5 см вдоль малой кривизны и на 6 см вдоль большой кривизны влево от первого разреза. Fundus сшивается с duodenum, и таким образом восстанавливается непрерывность пищеварительного канала. Из пилорической же части образуют слепой мешок, который вшивается в отверстие брюшной раны (Гейденгайн [67 [6]). Акерман зашивает изолированную часть с обоих концов и, отпрепаровав сальник на большой кривизне, делает фистульное отверстие в середине этой кривизны.
Изолирование с сохранением волокон блуждающего нерва производится также двумя различными способами. Изолируется либо весь пилорический отдел, либо только его часть. По первой методике (Крестев [*j) пилорический отдел отделяется от фундального разрезами, которые проникают через всю толщу желудочной стенки; лишь наверху на малой кривизне оставляют состоящий из serosa и muscularis мост, шириной в 3 см, соединяющий пилорическую часть с фундальной. Перегородку на этом месте делают лишь за счет слизистой. Двенадцатиперстная кишка отрезается и вшивается в fundus, или накладывают гастроэнтеростомоз. По второй методике (Шемякин [9] [3]) с пилорическим отделом поступают так же, как это описано для изолирования участка фундальной части с сохранением волокон блуждающего нерва, причем изолируемый участок пилорического отдела берут либо из большой, либо из малой кривизны. Происходящее при этом сужение привратниковой части не влечет за собой каких-либо заметных нарушений перехода желудочного содержимого в кишечник. Впрочем, надо заметить, что проведен ные до сих пор опыты и наблюдения не установили какой-либо значительной разницы в работе пилорических желез, была ли пилорическая часть изолирована с сохранением или без сохранения волокон блуждающего нерва.
При решении некоторых вопросов оказывается необходимой обыкновенная фистула, т. е. боковое отверстие в привратниковой части. Она накладывается либо так же, как на fundus, либо по особому способу, который будет описан по поводу фистул желчного пузыря и кишечника.
При изучении желез вообще и специально пепсиновых желез всплывает вопрос: с какой части поверхности пищеварительного канала происходит возбуждение соответственной секреции? Для решения этого вопроса необходимы некоторые дополнительные операции. Чтобы дать обоснованный ответ на вопрос о том, с какой поверхности химические возбудители оказывают свое воздействие на пепсиновые железы, необходимо проделать довольно сложную комбинацию операций в одном и том же животном, которые нужно проводить в определенной последовагельности. Прежде всего делается обыкновенный маленький желудочек фундальной части (Соколов [107] [10]). При данной комбинации разрез этой операции проводится слева вдоль наружного края m. rectus abdominis, начиная от края ребра. Пользуясь тем же разрезом, на большом желудке накладывают обычную желудочную фистулу. Фистульная трубка укрепляется в верхнем конце раневого разреза, отверстие маленького желудочка в нижнем его конце. Нужно позаботиться, чтобы кожа между обоими отверстиями была тщательно зашита, чтобы между ними образовался прочный кожный мост. Затем на двенадцатиперстную кишку накладывается боковая фистула с металлической канюлей. Кожный разрез для этой операции проводится справа у внешнего края m. rectus abdominis. О методике, по которой осуществляется эта фистула, будет подробно сказано ниже. Как только оба фистульных отверстия с канюлями обрастут прочной рубцовой тканью, через отверстие вдоль средней линии вытягивают желудок в месте перехода его в двенадцатиперстную кишку и делают перегородку между желудком и двенадцатиперстной кишкой. Эту перегородку образуют, либо проводя сплошной поперечный разрез, - стало быть, из всех слоев, либо только из слизистой. В последнем случае делают сначала продольный разрез через серозный и мышечный слои, кругаобразно отпрепаровывают слизистую перерезают поперек только ее. Перерезанные концы, так же как продольный разрез, зашиваются. После этой операции возникает крайне трудная задача кормления оперированной таким образом собаки. Лучше всего эта задача разрешается наружным гастроэнтеростомозом, т. е. желудочная канюля соединяется с дуоденальной достаточно широкими трубками из стекла или металла и из резины. Пищу собака получает лишь в размельченном виде. Переход пищи из желудка в кишечник происходит в общем довольно легко. Лишь временами, пока пища еще недостаточно разжижена желудочным соком, возникает необходимость протолкнуть рукой застрявшие в резиновой трубке пищевые массы. Дабы избежать повреждения животным этого соединения желудка с кишечником, приходится во время пищеварения держать собаку в станке. Когда же собаку спускают, канюли либо разъединяются, и каждая из них затыкается пробкой, или их соединяют тонкой резиновой трубкой, которая почти прилегает к брюшной стенке и мало заметна для собаки. Если собака не трогает эту трубку, не вырывает ее, выгоднее оставить собаку с этой трубкой, чем совершенно разъединить канюли. По этой трубке накапливающаяся в желудке жидкость непрерывно переходит постепенно в кишечник; она состоит из выделяющегося после пищеварения желудочного сока, из слюны, психического сока и накапливается в течение ночи в количестве многих сотен кубических сантиметров.
На такой собаке можно совершенно точно сравнивать влияние химических возбудителей пепсиновых желез то со слизистой кишечника, то с желудка.
Идя далее, можно установить разницу в действии такого возбудителя с разных отрезков желудка, т. е. с поверхности пилорического или фундального отдела. этой целью поступают вначале точно так же, как в предыдущем случае, но при последней операции поперечный разрез на желудке накладывают не на его границе с двенадцатиперстной кишкой, а на границе между фундальной и пилорической частью. При этом особое внимание обращают на то, чтобы каждый из разделенных отрезков желудка не содержал каких-либо частей другого. Это достигается широким удалением промежуточной области (В. Гросс [1]).
Как вивисекции, так и хирургические операции производились не только для получения желудочного сока и для исследования нормальной работы желудочных желез, но и для исследования иннервации этих желез. Здесь дело идет главным образом перерезке или раздражении блуждающего нерва и n. splanchnicus. Так как операции на этих нервах производились для исследования кровообращения и будут описаны в соответственной главе, то я приведу здесь только те варианты, которые применялись или могли быть применены специально для исследования иннервации желудочных и других пищеварительных желез. Сюда относятся разнообразные операции перерезки блуждающего нерва на различных уровнях. Простую перерезку nn. vagi на середине шеи оказалось возможным применить в качестве убедительного опыта для доказательства секреторного значения nn. vagi для пепсиновых желез только тогда, когда был найден способ сохранять собак в совершенно здоровом состоянии на неопределенно долгое время. При первом опыте (Павлов и Шумова-Симановская), доказавшем участие сn. vagi в секреторном эффекте мнимого кормления, применялась операция Генцмера [12] [12]. Она состоит в том, что n. vagus перерезают слева, как обычно - по середине шеи, справа же - под местом ответвления n. laryngeus inferior в большинства сердечных ветвей блуждающего нерва. Эта последняя операция проделывается следующим образом. Кожный разрез накладывается внизу шеи от верхнего края грудной клетки на 5-6м кверху к внешнему краю m. sternocleidomastoideus. После кожи перерезается фасция, разрывается соединительная ткань; при этом обнажаются снаружи vv. jugularis и subclavia, а снутри от щитовидного хряща и n. vagosympathicus с прилежащими art. carotides. Далее изолируют в глубине n. vagus под art. subclavia dextra, исключительно при помощи препаровальной иглы, причем n. vagus, который загибается вниз, поднимается на крючок, при положении животного на спине, и перерезается на 1-1.5 5 см ниже art. subclavia. Выгоднее перерезать сначала n. vagus на правой стороне, как описано, затем через две недели слева, на середине шеи, после того как в промежутке все оставшиеся ветки правого блуждаюшего нерва оправились после неизбежной при операции травмы. Такой перерезкой в два темпа достигается то, что животное после двусторонней перерезки nn. vagi не впадает в депрессивное состояние, наступающее после простой перерезки по середине шеи, ибо таким путем избегаются важные функциональные нарушения со стороны гортани и сердца.
На оперированной таким образом собаке, на которой прежде проделаны операции гастро- и эзофаготомии, ставится также опыт с раздражением блуждающего нерва как секреторного нерва пепсиновых желез. Для этой цели при перерезке левого блуждающего нерва (при последней операции) его периферический конец слегка отпрепаровывается и перевязывается шелковой ниткой, конец которой проводится через зашитую рану наружу. На следующий день у животного, спокойно стоящего в станке, снимают 1--2 кожных шва и с помощью шелковой нитки легко вытягивают наружу нерв, что не вызывает никакой реакции со стороны животного. Нерв раздражается индукционными ударами, посылаемыми каждую секунду в нерв. Таким образом нерв раздражается без наркотизации животного и без того, чтобы оно испытывало боль перед опытом или во время него (Павлов и Шумова-Симановская[15]).
Общее самочувствие животного, разумеется, еще меньше нарушается перерезкой nn. vagi на более низких уровнях, т. е. в грудной полости, под местом ответвления легочных ветвей, и в брюшной полости, чем только что описанной перерезкой nn. vagi на шее, хотя первые операции технически гораздо сложнее.
Операция перерезки в грудной полости производится двумя способами: по способу Клод Бернара"] и по способу Креля [15] (лаборатория Людвига). По Клоду Бернару, на соответственном месте грудной стенки разрезается кожа; разрезанная кожа отодвигается, и здесь же перерезается фасция и mm. intercostales. За Затем проникают пальцем в грудную полость, но так, чтобы туда не попал воздух. По пальцу вводится изогнутый крючком нож, лезвие которого находится на вогнутой стороне. Под контролем пальца перерезают оба nn. vagi, так же осторожно, чтобы не впустить воздух, вынимают из раны палец и нож и затем сдвигают кожу на ее прежнее место. Подобная операция нелегка, ее трудно правильно выполнить, и она могла удаваться только такому превосходному оператору, каким был Клод Бернар.
В лаборатории Людвига эта операция производилась гораздо более сложным способом, но зато полным ручательством. Сперва изготовляются резиновые колпаки с двойными стенками, широкое отверстие которых надевается на морду собаки, а узкое отверстие соединяется резиновой трубкой с дыхательным аппаратом. Чтобы колпак герметически прилетал к морде собаки, пространство между двойными стенками сильно надувается воздухом. Во время вскрытия грудной полости делается искусственное дыхание. На левой стороне вблизи позвоночника между шестым и седьмым ребрами делается надрез, который проникает в грудную полость. Ребра раздвигают как можно шире, вытягивают пищевод и перерезают на нем nn. vagi. Затем сильным надуванием легких выдавливают воздух из грудной полости и тщательно зашивают рану. /Животные прекрасно переносят эту операцию. Эти обе последние операции применились собственно для решения вопроса о причинах смерти после перерезки nn. vagi, но они вполне годятся и для опытов над иннервацией пепсиновых желез и других пищеварительных желез блуждающими нервами.
Операция перерезки nn. vagi в грудной полости имеет также несколько вариантов. Делается довольно длинной разрез вдоль средней линии, начинающийся у processus xiphoideus (Юргенсно [16] желудка отодвигают кпозвоночнику и вниз и, следуя затем по легко заметным ветвям nn. vagi, достигают без особых трудностей до обоих нервных стволов. Если хотят их перерезать возможно выше, то их присутствие нужно определить нащупыванием, для чего cardia на несколько секунд оттягивают книзу. Нервы отпрепаровывают от окружающей их ткани и перерезают или резецируют. Главным условием успеха операции, кроме самой тщательной чистоты, является оперирование в глубине раны in situ, ибо притягивание cardia к краю брюшной раны вызывает часто смерть собаки через 2-3 дня, вероятно вследствие шока. По Шиффу [17], перерезка нервов производится следующим образом. B regio hypochondrica sinistra накладывается разрез, длиною в 2-3 см, через который добираются до субдиафрагмального отдела пищевода; тогда под него подводят зонд и вытягивают наружу. Затем перерезают все видимые на поверхности пищевода нервы или резецируют их, переворачивают пищевод так, чтобы видны были задняя и боковые поверхности, и перерезают серозную оболочку циркулярно, не повреждая мышечного слоя.
Из этих двух способов следует предпочесть первый, во-первых, потому, что он гарантирует полноту перерезки, т. е. при этом перерезаются все брюшные ветви nn. vagi, и верхние окончания нервов оказываются в грудной клетке на 1-2 см выше диагфрагмы. Во-вторых, этот способ дает возможность оперировать in situ, что, как уже указано выше, имеет то преимущество, что собаки не погибают от операции.
Фруэн и Позерский [187 [18] перерезают nn. vagi из брюшной полости над диафрагмой следующим образом. Вдоль linea mediana делают разрез, длиною в 10 см, начинающийся у грудины, захватывают между двумя браншамизогнутого кишечного пинцета lig. gastrophrenicum и диафрагму за пищеводом и перерезают соединительнотканные тяжи, образующие предназначенное для пищевода отверстие в грудобрюшной преграды. Таким путем устраняется опасность пнесимоторакса от прободения диаФрагмы при наложении пинцета, соединяющего обе стороны грудобрюшной преграды - правую и левую. Затем вытягивают пищевод как можно дальше под грудобрюшную преграду и берут его на крепкую шелковину. Хорошо видамые теперь nn. vagi перерезаются таким образом гораздо выше грудобрюшной преграды. Зашив отверстие в грудобрюшной преграды, снимают пинцет и закрывают брюшную рану обычным способом.
Все эти способы перерезки nn. vagi все же оставили нерешенной одну методическую задачу, а именно задачу точного сравнения, во всех подробностях, деятельности пепсиновых желез при наличии секреторных волокон блуждающего нерва и без них. Однако в этом последнем случае крайне важно, чтобы не наступали никакие побочные нарушения секреторной работы пепсиновых желез, вызывающие недостаточную обработку пищи в желудке и ослабление его моторной функции и наступающие при всех вышеприведенных способах. Подобное сравнение может быть сделано лишь при изолированных маленьких желудочках, которые оперируются по Гейденгайну (без ветвей блуждающего нерва) или по Павлову (с сохранением иннервации блуждающего нерва). Но и в этом случае сравнение не могло бы быть совершенно бесспорным и простым, так как относилось бы к разным собакам и к различным маленьким желудочком. Требуемое сравнение выполнено совершенно точно Орбели [18] следующим образом. Были изготовленасколько возможно, удачные маленькие желудочки с сохранением иннервации. После того как их деятельность была разносторонне обследована и оказалась закономерной и стационарной, вскрывалась брюшная полость и перерезался серозно-мышечный мост между большим и маленьким желудком. При этом без всякого затруднения отделялись друг от друга оба слоя слизистой, образующие перегородку между двумя желудками. При этой операции оба эти слоя могли оставаться совершенно неповрежденными и отделение маленького желудка от большого протекало почти без потери единой капли крови и без какой-либо перевязки кровеносных сосудов. Единственная невыгода только что описанного способа состоит в том, что некоторые, действительно едва заметные, волокна блуждающего нерва остаются неперерезанными. Очевидно, эти редкие и скудные волокна проникают в маленький желудочек со стороны мезентерия. Их наличие обнаруживается незначительным возбуждением пепсиновых желез при еде.
В заключение заслуживают краткого упоминания некоторые пункты операционной методики, которые имеют целью раздражение блуждающего нерва как секреторного нерва пепсиновых желез в опытах ex tempore. Так как при какдом чувствительном раздражении работа пепсиновых желез сильно затормаживается, а при сильном раздражении это торможение становится упорным и длительным, то в случае вивисекции нужно начинать с уничтожения чувствительного раздражения, т. е. с перерезки спинного мозга под продолговатым.
Это можно выполнить различными способами: 1) можно применять очень кратковременный хлороформный наркоз, которого как раз хватает для перерезки спинного мозга и для введения трахеотомической трубки; 2) можно начинать с перерезки спинного мозга (в течение нескольких секунд) и сразу же вводить канюлю в трахею без всякого предшествующего наркоза. В последнем случае необходимо, чтобы голова привязанной за ноги к столу собаки держалась крепко и правильно и чтобы оператор мог производить перерезку точно и быстро двумятремя разрезами.
Второй пункт методики затрагивает необходимость перевязывать пищевод и pylorus при получении желудочного сока, чтобы предупредить поступление в желудок другого рода жидкостей изо рта и из кишечника (Ушаков [201] 7).
 Фистулы, наложенные на различных местах ductus choledochus, имеют то же физиологическое значение, как фистулы желчного пузыря. Изготовление подобной фистулы ex tempore не представляет никаких трудностей. Вскрыв ductus, в него вставляют канюлю по направлению к центру и фиксируют ее лигатурой. Постоянную же фистулу протока сделать очень трудно.
Явно целесообразный вариант фистулы ductus choledochus, предложенный С. Левашевым [3] [°], еще до сих пор не применялся и не подвергался никакой проверке. В искусственное боковое отверстие ductus choledochus вводят прямую, открытую с обоих концов и снабженную боковым отверстием трубочку и, поворачивая ее. вставляют так, чтобы это боковое отверстие приходилось против отверстия ductus choledochus.
Через отверстие протока в боковерстие трубочки ввинчивается под прямым углом другая трубочка; таким путем в протоке фиксируется Т-образная трубка. Ввинченная трубка выводится через брюшную рану приживляется в ней.
Фистула нормального окончания ductus choleodochus должна иметь иное значение, чем описанные фистулы, потому что условия перехода желчи в пищеварительный канал и их отношение к пищеварительному процессу могут быть изучены только таким способом. Простая пересадка куска дуоденальной стенки с нормальным отверстием ductus choledochus отверстие брюшной раны, как это делается в случае pancreas, большею частью не удается, так как обычно ductus choledochus разрывается вследствие сильного напряжения. Цель достигается следующей операцией (Брюно [1]). Из duodenum вырезается через всю его толщу языкообразный кусок стенки, длиною приблизительно в 1 см; примерно в его середине находится нормальное отверстие ductus choledochus. Свободный конец лоскута лежит книзу от этого отверстия; основание же, которым лоскут остается соединенным с дуоденальной стенкой, лежит над этим отверстием в направлении к желудку. Языкообразный кусок стенки перегибается у своего основания кверху и крепко пришивается своей слизистой к слизистой кишечной стенки. Оставшееся в кишечнике отверстие тщательно зашивается, причем отделение слизистой в верхней части отверстия на сгибе языкообразного лоскута заслуживает особого внимания, так как иначе не может иметь места хорошее заживление отверстия. Таким путем отверстие ductus choledochus переносится с внутренней стороны кишечной стенки на наружную. Затем двенадцатиперстная кишка вшивается тем местом, где находится перегнутый лоскут, в отверстие брюшной раны.
Чтобы все хорошо зажило, за раной нужно тщательно ухаживать. Если бы рана в кишечнике снова открылась, можно новым отделением краев отверстия и новыми швами снова добиться заживления. Сомнительный пункт этого способа заключается в том, что перегибание протока может оказать препятствие для свободного оттока желчи; поэтому была испробована модиФикация приведенного способа. После того как кишечное отверстие зашито и сама кишка фиксирована в брюшной ране, языкообразный лоскут с отверстием протока подшивается в самой ране из глубины раны к кожной поверхности. Мелчь собирается при помощи воронки, которая прикладывается к брюшной стенке.
Фистулы, наложенные на различных местах ductus choledochus, имеют то же физиологическое значение, как фистулы желчного пузыря. Изготовление подобной фистулы ex tempore не представляет никаких трудностей. Вскрыв ductus, в него вставляют канюлю по направлению к центру и фиксируют ее лигатурой. Постоянную же фистулу протока сделать очень трудно.
Явно целесообразный вариант фистулы ductus choledochus, предложенный С. Левашевым [3] [°], еще до сих пор не применялся и не подвергался никакой проверке. В искусственное боковое отверстие ductus choledochus вводят прямую, открытую с обоих концов и снабженную боковым отверстием трубочку и, поворачивая ее. вставляют так, чтобы это боковое отверстие приходилось против отверстия ductus choledochus.
Через отверстие протока в боковерстие трубочки ввинчивается под прямым углом другая трубочка; таким путем в протоке фиксируется Т-образная трубка. Ввинченная трубка выводится через брюшную рану приживляется в ней.
Фистула нормального окончания ductus choleodochus должна иметь иное значение, чем описанные фистулы, потому что условия перехода желчи в пищеварительный канал и их отношение к пищеварительному процессу могут быть изучены только таким способом. Простая пересадка куска дуоденальной стенки с нормальным отверстием ductus choledochus отверстие брюшной раны, как это делается в случае pancreas, большею частью не удается, так как обычно ductus choledochus разрывается вследствие сильного напряжения. Цель достигается следующей операцией (Брюно [1]). Из duodenum вырезается через всю его толщу языкообразный кусок стенки, длиною приблизительно в 1 см; примерно в его середине находится нормальное отверстие ductus choledochus. Свободный конец лоскута лежит книзу от этого отверстия; основание же, которым лоскут остается соединенным с дуоденальной стенкой, лежит над этим отверстием в направлении к желудку. Языкообразный кусок стенки перегибается у своего основания кверху и крепко пришивается своей слизистой к слизистой кишечной стенки. Оставшееся в кишечнике отверстие тщательно зашивается, причем отделение слизистой в верхней части отверстия на сгибе языкообразного лоскута заслуживает особого внимания, так как иначе не может иметь места хорошее заживление отверстия. Таким путем отверстие ductus choledochus переносится с внутренней стороны кишечной стенки на наружную. Затем двенадцатиперстная кишка вшивается тем местом, где находится перегнутый лоскут, в отверстие брюшной раны.
Чтобы все хорошо зажило, за раной нужно тщательно ухаживать. Если бы рана в кишечнике снова открылась, можно новым отделением краев отверстия и новыми швами снова добиться заживления. Сомнительный пункт этого способа заключается в том, что перегибание протока может оказать препятствие для свободного оттока желчи; поэтому была испробована модиФикация приведенного способа. После того как кишечное отверстие зашито и сама кишка фиксирована в брюшной ране, языкообразный лоскут с отверстием протока подшивается в самой ране из глубины раны к кожной поверхности. Мелчь собирается при помощи воронки, которая прикладывается к брюшной стенке.
 Для некоторых особых целей фистульная трубка для обычной боковой кишечной фистулы выбирается различной величины и различной формы (то круглая, то овальная); к ней также присоединяют дополнительные части. Так, к внутренней стенке припаивают тонкую трубочку, просвета фистульной трубки. Далее просвет трубки (который в этом случае должен быть овальныделяют продольнон перегородкой, чтобы отделить один отрезок кишечника от другого (Лондон [*]).
Пользуясь способом гастротомии, который применяется на человеке, Розенберг сделал у собаки бовую самозакрывающуюся кишечную фистулу, без канюли, с пробкой. Вот описание этого способа самим автором: «Я накладываю разрез длиною примерно в 8 см либо вдоль изгиба правого края ребер, либо несколько кнутри от правой мамиллярной линии, разъединяю брюшные слои и окаймляю кожу пристеночной брюшиной. Затем я вытягиваю пилорическую часть желудка и duodenum в область раны так, чтобы в нижнем углу раны появилась верхняя часть двенадцатиперстной кишки, длиною примерно в 1.5 см, с тем, чтобы остальная часть раны была заполнена желудком. В этом положении я пришиваю желудок и кишку к окаймленной брюшиной коже. Теперь я накладываю разрез, длиною в 5 см, на пилорическую часть желудка, кончающийся как раз на месте перехода желудка в кишку, так, чтобы serosa и muscularis расщепились и чтобы обнажилась слизистая. Исходя из этого разреза, в обе стороны на протяжение по крайней мере 1.5 см от слизистой отпрепаровывается серозно-мышечный лоскут. На месте перехода желудочной слизистой в кишечную последняя вскрывается, и в отверстие вводится заткнутая в направлении желудка толстостенная резиновая трубочка. Пришивая серозно-мышечный лоскут одной стороны через трубочку к такому же лоскуту другой стороны, причем каждый шов захватывает также стенку канюли, последнюю фиксируют в этом положении между серозномышечной и слизистой желудка.
«Рана посыпается каким-нибудь хирургическим порошком и покрывается складками нормальной кожи, которые сшиваются над раной. Через 5-8 дней швы прорезаются, и канюля выпадает. Чтобы предупредить зарастание слоев желудочной стенки, достаточно ежедневно на 15-30 минут вводить в отверстие стеклянную или каучуковую палочку. Через 4-6 недель фистула готова. До шестого послеоперационного дня собака не получает никакой пищи; со второго дня ей только впрыскивают под кожу 400 700 куб. см изотонического раствора поваренной соли. Начиная с шестого дня, собака получает небольшими порциями молоко и т. д.».
Из других операций, имеющих отношение к физиологии кишечных желез, остается упомянуть лишь о перерезке мезентереальных нервов. По Моро [6] и Ганау [7], она делается следующим образом. Главное требование при операции - это чтобы все нервы без исключения были перерезаны и чтобы ни один кровеносный сосуд не был поврежден. При отыскивании нервов пользуются стилетообразным полым зондом, несколько изогнутым на конце (который не должен быть острым); жолоб зонда находится на вогнутой поверхности изгиба. Инструмент подводится под один из листков брыжейки в направлении сосудов, предпочтительно над веной. Приподнятый на зонде лист брыжейки перерезается ножницами на протяжении одного сантиметра.
Лежащий на вене нерв берется на зонд и перерезается. Затем концом инструмента отделяется вся adventitia - и сосуд обнажен. Затем совершенно таким же способом изолируется артерия, причем перерезаются встречающиеся нервы. Если не окажется других сосудов, то изолированные сосуды приподнимаются и все остальное перерезается en masse.
Для некоторых особых целей фистульная трубка для обычной боковой кишечной фистулы выбирается различной величины и различной формы (то круглая, то овальная); к ней также присоединяют дополнительные части. Так, к внутренней стенке припаивают тонкую трубочку, просвета фистульной трубки. Далее просвет трубки (который в этом случае должен быть овальныделяют продольнон перегородкой, чтобы отделить один отрезок кишечника от другого (Лондон [*]).
Пользуясь способом гастротомии, который применяется на человеке, Розенберг сделал у собаки бовую самозакрывающуюся кишечную фистулу, без канюли, с пробкой. Вот описание этого способа самим автором: «Я накладываю разрез длиною примерно в 8 см либо вдоль изгиба правого края ребер, либо несколько кнутри от правой мамиллярной линии, разъединяю брюшные слои и окаймляю кожу пристеночной брюшиной. Затем я вытягиваю пилорическую часть желудка и duodenum в область раны так, чтобы в нижнем углу раны появилась верхняя часть двенадцатиперстной кишки, длиною примерно в 1.5 см, с тем, чтобы остальная часть раны была заполнена желудком. В этом положении я пришиваю желудок и кишку к окаймленной брюшиной коже. Теперь я накладываю разрез, длиною в 5 см, на пилорическую часть желудка, кончающийся как раз на месте перехода желудка в кишку, так, чтобы serosa и muscularis расщепились и чтобы обнажилась слизистая. Исходя из этого разреза, в обе стороны на протяжение по крайней мере 1.5 см от слизистой отпрепаровывается серозно-мышечный лоскут. На месте перехода желудочной слизистой в кишечную последняя вскрывается, и в отверстие вводится заткнутая в направлении желудка толстостенная резиновая трубочка. Пришивая серозно-мышечный лоскут одной стороны через трубочку к такому же лоскуту другой стороны, причем каждый шов захватывает также стенку канюли, последнюю фиксируют в этом положении между серозномышечной и слизистой желудка.
«Рана посыпается каким-нибудь хирургическим порошком и покрывается складками нормальной кожи, которые сшиваются над раной. Через 5-8 дней швы прорезаются, и канюля выпадает. Чтобы предупредить зарастание слоев желудочной стенки, достаточно ежедневно на 15-30 минут вводить в отверстие стеклянную или каучуковую палочку. Через 4-6 недель фистула готова. До шестого послеоперационного дня собака не получает никакой пищи; со второго дня ей только впрыскивают под кожу 400 700 куб. см изотонического раствора поваренной соли. Начиная с шестого дня, собака получает небольшими порциями молоко и т. д.».
Из других операций, имеющих отношение к физиологии кишечных желез, остается упомянуть лишь о перерезке мезентереальных нервов. По Моро [6] и Ганау [7], она делается следующим образом. Главное требование при операции - это чтобы все нервы без исключения были перерезаны и чтобы ни один кровеносный сосуд не был поврежден. При отыскивании нервов пользуются стилетообразным полым зондом, несколько изогнутым на конце (который не должен быть острым); жолоб зонда находится на вогнутой поверхности изгиба. Инструмент подводится под один из листков брыжейки в направлении сосудов, предпочтительно над веной. Приподнятый на зонде лист брыжейки перерезается ножницами на протяжении одного сантиметра.
Лежащий на вене нерв берется на зонд и перерезается. Затем концом инструмента отделяется вся adventitia - и сосуд обнажен. Затем совершенно таким же способом изолируется артерия, причем перерезаются встречающиеся нервы. Если не окажется других сосудов, то изолированные сосуды приподнимаются и все остальное перерезается en masse.

 [1] 1 См. эту книгу, стр. 247. - Ред.
[2] 1 2 Тр. Общ. русск. врачей в СПб., 1894 1895. (См. эту книгу, стр. 247. - Ред. ).
[3] 1Heidenhain, 3 in:] Hermann. Handbuch der Physiologie. [1880, S. 106 -115]. (Здесь и далее дополнения к библиографическим данным, стоящие в прямых скобках, принадлежат Редакции. - Ред. ).
[4] 4 влияние азотнокислого стрихнина отделение панкреатического сока у собаки. Диссертация, СПб., 1893.
[5] Bd. 5 1 XVI, [F o d 1896, e r a], [S. Moleschott's 79-89]. Untersuch. z. Naturlehre d. Mensch. u. d. Tiere,
[6] 2 6 Bull. de la Soc. des natur. de Moscou, t. 16, [1843].
[7] 3 7 [B l o n d l o t]. Traite analytique de la digestion. 1843.
[8] 8 [K l e m e n s i e w i c z), Sitzungsber. d. Wiener Akad., [Bd. LXXI, 1875, S. 249-296].
[9] 2 9 [He i d e n h a i n], Pflugers Archiv f. d. gen. Physiol., Bd. XVIII, 1878, [S. 169-171].
[10] 3 10 Ibidem, Bd. XIX, 1879, [S. 148 -166].
[11] 1 11 Кислотность определялась титрованием сока щелочью и выражалась в процентах HCI на всю массу сока.
[12] 1 12 [L u d w i gl, Zeitschr. f. rat. Med. , N. F., I, 1851, [S. 255-277].
[13] 1 13 R. H e i d e n h a i n, Stud. d. Physiol. Inst. zu Breslau, Bd. 4, 1868. Pfliger's Archiv f. d. ges. Physiol., Bd. XVII, 1878, [S. 1--67].
[14] 1 14 Schiff. Lecons sur la physiologie de la digestion. 1867.
[15] 15 R Regnard et Loye. Experiences sur un supplies, Progres medical, 1885, [t. II, № 29, p. 33 341.
[16] Bidder 16 u. C. Schmidt. Die Verdauungssafte u. s. w. 1852.
[17] 2 17 [R i c h e t Journ. de Tanat. et de la physiol... 1878, [p. 170--333].
[18] 1 18 Собака. которая служила для приведенного опыта, осталась жить многие месяцы. Впоследствии ей перерезан был и правый вагус на шее. Собака пользовалась отличным здоровьем, без преувеличения можно сказать - наслаждалась жизнью; за все это время многократные пробы с мнимым кормлением неизменно давали безусловно отрицательный результат относительно выделения желудочного сока. То же повторялось и на другой собаке, также много месяцев пережившей полную перерезку блуждающих нервов на шее.
[19] 19 [Bernstein], Berichte d. Sachsisch. Ges. d. Wiss. zu Leipzig, 1869, [S. 96 -131].
[20] 2 20 Pfluger's Archiv f. d. ges. Physiol., Bd. XVI. [1878, S. 173]. (См. первую книгу этого тома, стр. 49. -- Ред. ).
[21] 21 Об угнетающем влиянии на отделительные железы сока. . . Диссертация, СПб., 1882.
[22] 22 [He i d e n h a i n] Pfluger's Archiv f. d. gen. Physiol., Bd. X, 1875.
[23] 1 23 L a n d a u. Zur Physiologie der Bauchspeichelabsonderung. Dissertation, Breslau, 1873.
[24] 24 Под внешним деятелем я понимаю безразлично как агента внешней природы, так и всякого агента внутри организма; следовательно, слово внешний обозначает все, кроме самой нервной системы.
[25] 25 (H e i d e n h a i n). Uber die Absonderung der Fundusdrusen des Magens. Pfluger's Archiv f. d. ges. Physiol... 1879, [Bd. XIX, S. 148 166].
[26] 26 Как в этом, так и в последующих списках работ данные, стоящие в прямых скобках, принадлежат Редакции. - Ред.
[27] 27 Тр. Общ. русск. врачей в СПб., т. 64, май, 1897, стр. 581-589. [См. также: 1) Больн. газ., № 22, 1897, стр. 809-814; 2) Uber einen pathologisch-therapeutischen Versuch uber Magensaftabsonderung des Hundes. Arch. f. Verdauungskr., Bd. 3, 1898, S. 488-489].
[28] 28 Больничн. газета Боткина, 1897, стр. 1569. - См. также: Тр. Общ. русск. врачей в СПб., т. 65, сентябрь, 1897, стр. 25--30.
[29] 29 Архив биол. наук, т. V. 1897, стр. 213-224. - [См. также: Zur Frage uber den Ort der Harnstoffbildung bei den Saugetieren]. Archiv f. exper. Pathol. u. Pharmacol., Bd. XXXVIII, 1897, S. 215-222.
[30] 1 30 Schondorff, Pfluger's Archiv f. d. ges. Physiol., Bd. LIV, S. 423.
[31] 2 31 Архив биол. наук, т. IV, 1896, стр. 241.
[32] 32 Киевск. университетск. изв. , 1894, и I. B. M a l y s, 1896, стр. 315.
[33] 33 Ka u f m a n n. Nouvelles recherches sur le lieu de formation de Turee dans l'organisme animal. Role preponderant du fole dans cette formation. Archive de physiol., v. XXVI, p. 531-546; Jahresber. f. Tierchemie, 1895, S. 172.
[34] 2 34 Судя по предварительному сообщению Шендорфа, мышцы содержат мочевину в количествах, не допускающих отнесения ее на счет крови, пропитывающей мышечную ткань (Pfluger's Archiv, 1895).
[35] 35 Ср.: Mintzer, Archiv f. exper. Pathol. S. 197; Richter, Berl. klin. Wochenschr., Bd. XXXIII, 1896, S. 454.
[36] 2 36 Архив биол. наук, т. IV, 1896, стр. 212. (См. первую книгу этого тома, стр. 287. - Ред. ).
[37] 3 37 L. cit. , S. 193.
[38] 1 38 Архив биол. наук, т. I, 1892, стр. 400.
[39] 39 Magnamini. Le modification del recombrio azotato dopo Timnesto della vena porta colla vena cava inferiore. II Policlinico, v. III, 1896, p. 11.
[40] 2 40 2 Queirolo, Untersuch. z. Naturlehre d. Mensch, u. d. Tiere, Bd. XV, 1895.
[41] 41 Lieblein, Archiv f. exper. Pathol. u. Pharmakol., Bd. XXXIII, S. 332.
[42] 2 42 Архив биол. наук, т. III, [1894].
[43] 3 43 Там же, т. II, [1893].
[44] 44 Там же, т. I, 1892, стр. 400. (См. первую книгу этого тома, стр. 210. - Ред. ).
[45] 45 Mi n t z e r, Archiv f. exper. Pathol. u. Pharmacol., Bd. XXXIII, S. 182.
[46] 46 Речь, читанная в торжественном заседании Общества русских врачей в память С. П. Боткина, 1899 г. [Больн. газ., 1900. № 14--15, стр. 617630; № 16, стр. 703-709; № 17, стр. 752-756; № 18, стр. 804-811.См. также: Труды Общ. русск. врачей, т. 67, 1900, ноябрь-декабрь, стр. 197-242].
[47] 47 Эти опыты были начаты доктором Д. Л. Глинским и продолжались доктором С. Г. Вульфсоном на собаках, у которых нормальные концы слюнных протоков были выведены предварительной операцией из полости рта на кожу. Во время опытов на коже около отверстия протока приклеивалась воронка с подвешенным цилиндриком. Слюна подчелюстной и подъязычной (слизистых) желез собиралась вместе, слюна околоушной - отдельно.
[48] 48 Операция эта не очень легка. После нескольких попыток мы остановились на следующем приеме: отверстие желчного протока с куском слизистой оболочки выводится из полости кишки на ее поверхность и здесь пришивается, а затем эта кишка соответственным местом укрепляется и приживляется в отверстии брюшной раны.
[49] 49 Автор несколько раз ранее делал сообщения о ниже упоминаемом исследовании.
[50] 1 50 См. том 1, стр. 536. -- Ред.
[51] 51 [Die physiologische Chirurgie des Verdauungskanals]. Ergebn. d. Physiol... I. Jarhrg., 1. Abteil., 1902, [S. 246 284].
[52] 1 52 Данные, стоящие в прямых скобках, принадлежат Редакции. - Ред.
[53] Тр. 53 Общ. русск. врачей в СПб., т. 69, март- май, 1902, стр. 405 417. - Бо - Больничн, газета Боткина, № 33, 1902, [стр. 1481-1487].
[54] 54 Тр. Общ. русск. врачей в СПб., т. 70, сентябрь октябрь, 1902, стр. 139-143. - Больничн. газета Боткина, 1902, [№ 47, стр. 2153 2155].
[55] 55 Les prix Nobel en 1904. Stockholm, 1906.
[56] 56 Тр. Общ. русск. врачей в СПб., т. 71, март-май, 1904, стр. 39-54.
[57] 57 Изв. Военно-медиц. акад. , т. IX, 1904, сентябрь, стр. 3. [См. также: Uber die ein und demselben Eiweissfermente zukommende proteolytische und milchkoagulierende Wirkung verschiedener Verdauungssafte Zeitschr. f. physiol. Chemie, Bd. XLII, 1904, S. 415 452.
[58] 58 [Glassner], Beitrage zur chemischen Physiologie und Pathologie, Bd. I, 1901.
[59] 59 [Loevenhart], Zeitschr. f. physiol. Chemie, Bd. XLI.
[60] 2 60 Диссертация, СПб., 1901.
[61] 61 Zeitschr. f. physiol. Chemie, Bd. XXXII, [1901, S. 291-319]. 19).
[62] 2 62 [Pekelharing], ibidem, Bd. XXXV, [1902, S. 8 30].
[63] 63 3 Тр. Общ. русск. врачей в СПб. за 1900-1901 гг. и протокол заседания 21 января 1901 г.
[64] 1 64 [Die aussere Arbeit der Verdauungsdrusen und ihr Mechanismus). Nagel's Handbuch d. Physiol. d. Mensch. , Bd. II, 1. H., 1906 -1907, [S. 666 -743]. Классическое описание секреторных процессов дал Гейденгайн в «L. Hermann's Handbuch der Physiologie" (1880). В «Schaftr's Textbook of Physiology» (1893) имеется в высшей степени продуманное и обширное описание Ленгли физиологии слюнных желез. Как первая, так и вторая статья, особенно последняя, снабжены богатыми литературными указаниями. Результаты нашей лаборатории о работе пищеварительных желез собраны в моей книге «Работа пищеварительных желее» (1898), французский перевод которой появился в 1901 г., а английский - в 1902 г.
[65] Тр. 65 Общ. русск. врачей в СПб., 1895.
[66] 2 66 Диссертация, СПб., 1898.
[67] 67 3 Диссертация, СПб., 1901.
[68] 68 [M a l l o i z e l], Journ. de physiol. et pathol. gener., 1902, [t. 4, p. 547, 749, 926].
[69] 69 5 Диссертация, СПб., 1904.
[70] 6 70 Диссертация, СПб., 1904.
[71] 71 [Billard et Dieulafe], Compt, rend. de lac. de biol. a Paris, 1902.
[72] Русский. 72 врач. 1903, [№ 23, стр. 869--872].
[73] 73 Forhandlingar vid Nord. Naturforsekareoch Lakaremotet. Helsingfors, 1909.
[74] 74 [L u d w i gl, Zeitschr. f. rat. Med. , N. F., L., 1851, [S. a55-277].
[75] 75 2 [S c h i f f], Archiv f. physiol. Heilkunde, 1851.
[76] 3 76 [Cl. Bernard]. Lecons sur la physiologie et la pathologie du systeme nerveux, t. II, 1858.
[77] 77 [E c k h a r d], Eckhard's Beitrage... 2, 1860, [S. 81-98].
[78] 5 78 L. cit.
[79] 6 79 [S c h i f f]. Lehrbuch der Muskel- und Nervenphysiologie. 1858-1859.
[80] 7 80 [N a v r o t z k yl, Studien d. Physiol. Inst. zu Breslau, Bd. 4, 1868, [H. 4, S. 125-145].
[81] 8 81 [Loeb], Eckhard's Beitrage, V, 1869, [S. 1-26].
[82] 9 82 [He i d e n h a i n], Pfleger's Archiv f. d. ges. Physiol., Be. XVII, 1878. [S. 1-67].
[83] 83 [L a n g l e yl, Philosophic. Transact. of the Royal Soc. (в дальнейшем: Phil. Trans. ), London, 1892, [p. 85--124].
[84] 84 [H e i d e n h a i n], Stud. d. Physiol. Inst. zu Breslau, 1868, [Bd. 4, Н. 4, S. 1-124], и Pfluger's Archiv f. d. ges. Physiol., Bd. XVII, 1878, [S. 1-67].
[85] 2 85 [Werther], Prliger's Archiv f. d. ges. Physiol., Bd. XXXVIII, 1886, S. 293 -311].
[86] 3 86 [Fletcher], Phil. Trans. , 1889, [vol. 180, p. 109--154].
[87] 4 87 [He i d e n h a i n], l. cit.
[88] 88 [He i d e n h a i n], l. cit.
[89] 1 89 [Becher und Ludwigl, Zeitschr. f. rat. Med. , N. F., I, 1851, [S. 278 284].
[90] 90 [H e i d e n h a i n], Pflugers Archiv f. d. gen. Physiol., Bd. XVII, 1878, [S. 1-67].
[91] 2 91 [L a n g l e y], Phil. Trans. , 1892, [р. 85--124].
[92] 92 [Zerner], Mediz. Jahrbucher, 1887, [S. 530-538].
[93] 1 93 [L u d w i gl, Sitzungsber. d. Wien. Akad., mathem.-nature. Kl., XXV, [S. 584 590], и Wien. med. Wochenschr., 1860, [№ 28 -29, стлб. 433-437, 449-454].
[94] 2 94 [B a y l i s s and Hill, Journ. of Physiol., vol. 16, 1904, [p. 351-359].
[95] 95 [B a y l i s s and B r a d f o r dj, Proceed. of the Royal Soc. , London, 1886, [vol. XL, p. 203-206].
[96] 2 96 [Bradford], Journ. of Physiol., vol. 8, 1887, [р. 86-98].
[97] 3 97 [H e i d e n h a i n], Stud. d. Physiol. Inst. zu Breslau, Bd. 4, 1868, [H. 4, S. 1-124].
[98] 98 [L a n g l e y], Journ, of Physiol., vol. 2, [1879-1880, р. 261-280j.
[99] 1 99 [He i d e n h a i n], 1. cit.
[100] 2 100 2 Врач, 1890, [№ 7, стр. 153-156; № 9, стр. 210-212; № 10, стр. 231--234]. (См. первую книгу этого тома, сто. 142. -- Ред. ).
[101] 3 3 101 Диссертация, СПб., 1890.
[102] 102 [Henderson], Americ. Journ. of Physiol., vol. 3, 1900, [р. 19-25].
[103] 103 a r c r o f t ), Journ. of Physiol., vol. 27, 1901, [p. 31-47].
[104] 6 104 [Ke u c h e l], Диссертация, Юрьев, 1878.
[105] 7 105 [L a n g l e y]. Journ, of Physiol., vol. 9, 1888.
[106] 1 106 [B a r c r o f t], 1. cit.
[107] 2 107 [Cl. Bernardl, Compt. rend. de la Soc. de biol. a Paris, 1858.
[108] 108 3 [H e i d e n h a i n], Pfliger's Archiv f. d. ges. Physiol., Bd. V, 1872, ip. 309 -318].
[109] 4 109 [B e i n b r i d g el, Journ. of Physiol., vol. 26, 1900--1901, [р. 79-91].
[110] 5 110 [L a n g l e y], ibidem, vol. 11, 1890, [p. 123 158].
[111] 6 111 [Cl. Bernard], Journ. de Tanat. et physiol., t. I, 1864, [p. 507-513].
[112] 112 7 [L a n g l e yl, Journ. of Physiol., vol. 6, 1885, [p. 71-92].
[113] 113 [H e i d e n h a i n), Stud. d. Physiol. Inst. zu Breslau, Bd. 4, 1868, [H. 4, S. 1-124].
[114] 2 114 [E c k h a r d], Eckhard's Beitrage. . . Bd. 2, 1860, [S. 205--216].
[115] 115 3 [L a n g l e y], Journ. of Physiol., vol. 1, 1878, [p. 96--103].
[116] 4 116 [Heidenhain], l. cit.
[117] 117 [L a n g l e y], Schafer's Textbook of Physiol., 1898, [vol. I, p. 475-530'
[118] 6 118 [Bradford], Journ. of Physiol., vol. 8, 1887, [p. 86 -98].
[119] 1 119 [H e i d e n h a i nl, Pfluger's Archiv f. d. gen. Physiol., Bd. V, 1872, [S. 143-153].
[120] 2 120 [B a i n b r i d g el, Journ. of Physiol., vol. 26, (1900-1901), [р. 79-91].
[121] 2 121 [L a n g l e yl, ibidem, vol. 11, 1890.
[122] 122 [B r a d f o r d], ibidem, vol. 9, 1888, [p. 287--316].
[123] 1 123 [He i d e n h a i n], l. cit.
[124] 124 [L a n g l e y], Journ. of Physiol., vol. 1, 1878, [p. 96-103].
[125] 3 125 [L a n g l e yl. Journ. of Physiol., vol. 10, 1889, [p. 291-328].
[126] 3 126 [M a t h e w s], Americ. Journ. of
[127] 1 127 [B u n c h], ibidem, vol. 26 (1900 Physiol., vol. 4, 1900, ip. 482-489].
[128] 1 128 [He i d e n h a i n), Pfluger's Archiv f. d. gen. Physiol., Bd. XVII, 1878, [S. 1-67].
[129] 2 129 [Langley], Journ, of Physiol., vol. 10, 1889, [p. 291-328].
[130] 130 3 [M a t h e w s], Annal. of the New-York Acad. of Sci. , vol. II, 1898, и Americ. Journ. of Physiol., vol. 4, 1900, [p. 482 499].
[131] 131 [He i d e n h a i n], Pflugers Archiv f. d. gen. Physiol., Bd. XVII, 1878, 1-67].
[132] 132 [M a l l o i z e l], 1. cit.
[133] 2 133 [C z e r m a k], Sitzungsber. d. Wien. Acad., mathem.-naturw. Kl., XXV, 1857, [S. 3-18].
[134] 134 [Eckhard], Eckhard's Beitrage... 2, 1860, [S. 81-98].
[135] 2 135 [Heidenhain], Pfliger's Archiv f. d. ges. Physiol., Bd. XVII, 1878.
[136] 3 136 [L a n g l e y], Journ. of Physiol., vol. 1, 1878, [p. 339. --369].
[137] 137 [Bradford], ibidem, vol. 9, 1888, [p. 287-316].
[138] 1 138 [Heidenhain, in:] Hermann. Handbuch der Physiologie. 1880.
[139] 2 139 Врач, 1890, [№ 7, стр. 153--156; 9, стр. 210-212; № 10, стр. 231-234].
[140] 3 140 [B r a d f o r dl, l. cit.
[141] 141 [Langley], Journ. of Physiol., vol. 6, 1885, [p. 71-92].
[142] 5 142 Диссертация, СПб., 1891.
[143] 1 143 [Cl. Bernard), цитировано по Langley.
[144] 2 2 144 Melang. biol. Acad. imp. d. sc. de St.-Petersbourg, 8, 1872, [стр. 651665].
[145] 3 145 [B u f f], Eckhard's Beitrage. . . . 12, 1888, [S. 3 -39].
[146] 4 146 Диссертация, СПб., 1894.
[147] 147 [Heidenhain, in:] Hermann. Handbuch der Physiologie. 1880.
[148] 148 2 Pfluger's Archiv f. d. ges. Physiol., Bd. XVI, 1878. (См. первую книгу этого тома, стр. 9. -- Ред. ).
[149] 1 149 Диссертация, СПб., 1904.
[150] 1 150 L. cit.
[151] 1 151 Диссертация, СПб., 1901.
[152] 2 152 [Cl. Bernard], Compt. rend. , 1852,
[153] 1 153 [Schiff]. Lecons sur la physiologie de la digestion. 1867.
[154] 2 154 [L a n g l e y and Anderson l, Journ. of Physics., vol. 16, 1894, [p. 410 440].
[155] 155 [Cl. B e r n a r d]. Lecons sur la physiologie et la pathologie du systeme nerveux. 1858.
[156] 156 [L o e b], Eckhard's Beitrage. . 5, 5, 1870, [S. 73-145].
[157] 5 157 [Grutzner und Chlapowski], Pfliger's Archiv f. d. ges. Physiol., Bd. VII, 1873, [S. 522-529].
[158] 6 158 [Lepine], Gaz, med. de Paris, 1875, [t. IV, № 27, р. 332-333; № 35, р. 436-437].
[159] 1 159 [B o c h e f o n t a i n e], Archives de physiol. norm. et pathol., 1876, [2 ser., t. 3, p. 140 -172].
[160] 2 160 Neurolog. Centralbl., Bd. 7, 1888, [S. 505-509].
[161] 3 161 [E c k h a r d], ibidem, Bd. 8, 1889.
[162] 4 162 [F l e c k], Dissertation, Giessen, 1889.
[163] Невролог. 163 вестн., 1899, [т. 7, № 4, стр. 1--6].
[164] 6 164 Диссертация, СПб., 1900.
[165] 165 Verhandl. d. Kongr. in Helsingfors, 1902.
[166] 2 166 Диссертация, СПб., 1904.
[167] 167 Изв. Общ. естествоиспыт. в Москве, т. 16, [1843, стр. 315].
[168] 2 168 [B o n d l o t]. Traite analytique de la digestion. 1843.
[169] 169 [H e i d e n h a i nj, Pfluger's Archiv f. d. ges. Physicl., Bd. XIX, 1879, [S. 169-171].
[170] 170 Тр. Общ. русск. врачей в СПб., 1894, [март, сто. 24--28].
[171] 1 171 [Fremont], Bull. de I'Acad. de med. , Paris, 1895, [t. XXXIV, p. 509].
[172] 2 172 Archiv f. Anat. u. Physiol., 1895. (Предварительное сообщение см.: Centralbl. f. Physiol., 1889, [Bd. III, № 6, S. 113-114]. (См. первую книгу этого тома, стр. 138 и 175. --Ред.).
[173] 1 173 Тр. Общ. русск. врачей в СПб., 1900, [сентябрь октябрь, стр. 197--242].
[174] Archiv 174 f. Anat. u. Physiol., 1894, [S. 58-82]. - Диссертация, СПб., 1889.
[175] 1 175 Диссертация, СПб., 1891.
[176] 2 176 [Sc h z], Zeitschr. f. physiol. Chemie, Bd. IX, 1890, [S. 577-590].
[177] 3 177 Диссертация, СПб., 1890.
[178] 178 Zeitschr. f. physiol. Chemie, Bd. XXXV, [1902, S. 8-30].
[179] 179 Диссертация, СПб., 1902.
[180] 180 Архив биол. наук, т. III. 1894, [стр. 453-515].
[181] 181 L. cit.
[182] 182 [Bidder und Schmidt]. Die Verdauungssafte. 1852.
[183] 183 Verhandl, d. Kongr. in Helsingfors, 1902.
[184] 2 184 Диссертация, СПб., 1903.
[185] 185 Еженед. клинич. газета Боткина, 1897, [№ 41, стр. 1569--1573]. (См. эту книгу, стр. 226. - Ред. ).
[186] 186 Архив биол. наук, т. V, 1896, [стр. 417--519].
[187] 1 187 Диссертация, СПб., 1904.
[188] 2 188 L. cit.
[189] 1 189 [Schiff]. Lecons sur la digestion. 1867.
[190] 2 190 Еженед. клинич. газета Боткина, 1897, [№ 22, стр. 809-814].
[191] 3 191 [L e c o n t e], La Cellule, 17, 1900, [p. 284--318].
[192] 192 [Leconte], l. cit.
[193] Тр. 193 Общ. русск. врачей в СПб., 1904, [год 71-й, март- май. стр. 5-111.
[194] 1 194 Диссертация, СПб., 1898.
[195] 195 Диссертация, СПб., 1899.
[196] 2 196 Диссертация, СПб., 1900.
[197] 197 cit.
[198] Pfliger's 198 Archiv f. d. ges. Physiol., Bd. LXXXIV, 1901, [S. 101-114].
[199] 2 199 L. cit.
[200] 200 [Meisl], Wien. klin. Rundschau (1903 и. 1904).
[201] 2 201 Обозрение психиатрии, 1899.
[202] 202 L. cit.
[203] 203 Диссертация, СПб., 1882.
[204] 204 [Axenfeld], Atti e rendic. della Acad. med. -chirurg. di Perugia, 1890, [vol. 2, parte 1, p. 142-176].
[205] 2 205 2 [C o n t e j e a n]. These de Paris. 1892.
[206] 206 [S c h n e y e r], Deutsche med. Wochenschr., 1896, [№ 12, S. 173].
[207] 207 Архив биол. наук, 1896, [т. IV, стр. 425-448].
[208] 208 Архив биол наук, т. I, 1892, [стр. 586-709].
[209] 209 Русск. врач, 1903, [№ 32, стр. 1121-1123].
[210] 210 [Hornborg] Skand. Archiv. f. Physiol., Bd. 15, 1904, [S. 209-258].
[211] 108). 211 Тр. Общ. русск. врачей в СПб. (1903-1904), [год 71-й, стр. 95
[212] L. 212 cit.
[213] 2 213 [Arthusj, Compt. rend. de la Soc. de biol. a Paris, 1903, [t. 55, p. 475-476].
[214] 3 214 Русск. врач, 1903.
[215] 215 L. cit.
[216] 216 L. cit.
[217] L. 217 ait.
[218] 1 218 Тр. С.-Петербургск, общ. естествоиспыт., т. XI, 1880. (См. первую книгу этого тома, стр. 88. - Ред. ).
[219] 2 219 [Heidenhain, in:] Hermann. Handbuch der Physiologie. 1880.
[220] 220 [Fodera, in:] Moleschott, Untersuch. z. Naturlehre d. Mensch. u. d. Thiere, Bd. XVI, 1896, [S. 79-89].
[221] 221 [Frouin], Compt. rend. de I'Acad. d. Sc., 1902, [p. 890-893].
[222] 2 222 Диссертация, СПб., 1899.
[223] [Bernstein]. 223 Berichte d. Sachsisch. Ges. d. Wiss. zu Leipzig, 1869.
[224] 2 224 [Heidenhain, in:] Hermann. Handbuch der Physiologie. 1880.
[225] 225 3 Диссертация, СПб., 1888.
[226] 4 226 Архив биол. наук, 1899, [т. VII, стр. 1-86].
[227] 1 227 Диссертация, СПб., 1901.
[228] 2 228 Archiv f. exper. Pathol. u. Pharmacol., Bd. XX, [1886, S. 367--384].
[229] 1 229 [Delezennel Compt. rend. de la Soc. de biol. a Paris (1902 et 1903), [t. 54 et t. 55].
[230] 230 Диссертация. СПб., 1899.
[231] 231 Архив биол. наук, 1893, [т. II, стр. 432-460].
[232] 2 232 [Gottlieb], Archiv f. exper. Pathol. u. Pharmacol... Bd. XXXIII, 1894, [S. 261--285].
[233] 3 233 Архив биол. наук, 1894, [т. III, стр. 395-421].
[234] 1 234 [Gottlieb], I. cit.
[235] 2 235 Диссертация, СПб., 1896.
[236] 236 [Wertheimer et Lepage], Journ. de physiol. et de pathol. gener., 1901, [t. III, р. 821].
[237] 4 Тр. 237 Общ. русск. врачей в СПб., 1896, [год 63-й, февраль, стр. 7--14].
[238] 5 238 Тр. Конгресса в Гельсингфорсе, 1902. 02.
[239] 239 [CI. Bernard]. Memoire sur le pancreas. 1856.
[240] 2 240 [B ernstein], Berichte d. konigl. Sachsisch. Ges. d. Wiss. zu Leipzig, 1869, [Bd. 21, S. 96-131].
[241] 3 241 Pfliger's Archiv f. d. ges. Physiol., Bd. XVI, [1878, S. 173-189]. (См. первую книгу этого тома, стр. 49. - Ред. ).
[242] 4 242 [H e i d e n h a i n], Pfluger's Archiv f. d. ges. Physiol., Bd. X. [1875, S. 557-632].
[243] 5 243 Archiv f. Anat. u. Physiol., 1893, [Suppl. Bd., S. 176 -200]. (См. первую книгу этого тома, стр. 96. - Ред. ).
[244] 1 244 Диссертация, СПб., 1896.
[245] 2 245 [Wertheimeret Lepage], Compt. rend. de la Soc. de biol. a Paris, 1901, [t. 53, p. 879-880].
[246] 246 Протоколы Конгресса в Гельсингфорсе, 1902.
[247] 2 247 Archiv f. Anat. u. Physiol., 1894, [Physiol. Abt., S. 83-116].
[248] 1 248 Диссертация, СПб., 1896.
[249] 2 249 [Bernstein], l. cit.
[250] 3 250 [CI. Bernard]. Lecons sur les proprietes physiologiques des liquides de lorganisme. 1859.
[251] 251 Клинич. газета Боткина, 1900, [Больн. газета Боткина, № 28, стр. 1272-1288].
[252] 2 2 252 [Wertheimer et Lepage], Journ. de physiol. et de pathol. gener... 1901, [t. III, p. 335-348, 363-374].
[253] 3 253 [B a y l i s s and Starling Journ. of Physiol., vol. 28, 1902, Ip. 325--353].
[254] 254 [E n r i q u e s et H a l l i o n], Compt. rend. de la Soc. de biol. a Paris, 1903.
[255] 255 [Wertheimer], Journ. de physiol. et pathol. gener., 1901, [t. III, p. 689-702, 708-718].
[256] 2 256 [Fleigl, Compt. rend. de la Soc. de biol. a Paris, 1903, [t. 55, p. 462 463].
[257] 3 257 Тр. Общ. русск. врачей в СПб., 1903, [год 71-й, ноябрь-декабрь, стр. 99-103].
[258] 1 258 [Fleig], Journ. de physiol. et pathol. gener., 1904, [t. VI, р. 148].
[259] 259 [Wertheimeret Lepage], l. cit.
[260] 260 Тр. Общ. русск. врачей в СПб., т. 75, март-май, 1908, стр. 456-458.
[261] 261 [Die operative Methodik des Studiums der Verdauungsdrisen]. Tigerstedt's Hande. d. physiol. Methodik, Bd. II, I. Halfte, 2. Abt., 1911, S. 150 -188.
[262] 262 Данные, стоящие в прямых скобках, принадлежат Редакции. - Ред.
[1] 1 См. эту книгу, стр. 247. - Ред.
[2] 1 2 Тр. Общ. русск. врачей в СПб., 1894 1895. (См. эту книгу, стр. 247. - Ред. ).
[3] 1Heidenhain, 3 in:] Hermann. Handbuch der Physiologie. [1880, S. 106 -115]. (Здесь и далее дополнения к библиографическим данным, стоящие в прямых скобках, принадлежат Редакции. - Ред. ).
[4] 4 влияние азотнокислого стрихнина отделение панкреатического сока у собаки. Диссертация, СПб., 1893.
[5] Bd. 5 1 XVI, [F o d 1896, e r a], [S. Moleschott's 79-89]. Untersuch. z. Naturlehre d. Mensch. u. d. Tiere,
[6] 2 6 Bull. de la Soc. des natur. de Moscou, t. 16, [1843].
[7] 3 7 [B l o n d l o t]. Traite analytique de la digestion. 1843.
[8] 8 [K l e m e n s i e w i c z), Sitzungsber. d. Wiener Akad., [Bd. LXXI, 1875, S. 249-296].
[9] 2 9 [He i d e n h a i n], Pflugers Archiv f. d. gen. Physiol., Bd. XVIII, 1878, [S. 169-171].
[10] 3 10 Ibidem, Bd. XIX, 1879, [S. 148 -166].
[11] 1 11 Кислотность определялась титрованием сока щелочью и выражалась в процентах HCI на всю массу сока.
[12] 1 12 [L u d w i gl, Zeitschr. f. rat. Med. , N. F., I, 1851, [S. 255-277].
[13] 1 13 R. H e i d e n h a i n, Stud. d. Physiol. Inst. zu Breslau, Bd. 4, 1868. Pfliger's Archiv f. d. ges. Physiol., Bd. XVII, 1878, [S. 1--67].
[14] 1 14 Schiff. Lecons sur la physiologie de la digestion. 1867.
[15] 15 R Regnard et Loye. Experiences sur un supplies, Progres medical, 1885, [t. II, № 29, p. 33 341.
[16] Bidder 16 u. C. Schmidt. Die Verdauungssafte u. s. w. 1852.
[17] 2 17 [R i c h e t Journ. de Tanat. et de la physiol... 1878, [p. 170--333].
[18] 1 18 Собака. которая служила для приведенного опыта, осталась жить многие месяцы. Впоследствии ей перерезан был и правый вагус на шее. Собака пользовалась отличным здоровьем, без преувеличения можно сказать - наслаждалась жизнью; за все это время многократные пробы с мнимым кормлением неизменно давали безусловно отрицательный результат относительно выделения желудочного сока. То же повторялось и на другой собаке, также много месяцев пережившей полную перерезку блуждающих нервов на шее.
[19] 19 [Bernstein], Berichte d. Sachsisch. Ges. d. Wiss. zu Leipzig, 1869, [S. 96 -131].
[20] 2 20 Pfluger's Archiv f. d. ges. Physiol., Bd. XVI. [1878, S. 173]. (См. первую книгу этого тома, стр. 49. -- Ред. ).
[21] 21 Об угнетающем влиянии на отделительные железы сока. . . Диссертация, СПб., 1882.
[22] 22 [He i d e n h a i n] Pfluger's Archiv f. d. gen. Physiol., Bd. X, 1875.
[23] 1 23 L a n d a u. Zur Physiologie der Bauchspeichelabsonderung. Dissertation, Breslau, 1873.
[24] 24 Под внешним деятелем я понимаю безразлично как агента внешней природы, так и всякого агента внутри организма; следовательно, слово внешний обозначает все, кроме самой нервной системы.
[25] 25 (H e i d e n h a i n). Uber die Absonderung der Fundusdrusen des Magens. Pfluger's Archiv f. d. ges. Physiol... 1879, [Bd. XIX, S. 148 166].
[26] 26 Как в этом, так и в последующих списках работ данные, стоящие в прямых скобках, принадлежат Редакции. - Ред.
[27] 27 Тр. Общ. русск. врачей в СПб., т. 64, май, 1897, стр. 581-589. [См. также: 1) Больн. газ., № 22, 1897, стр. 809-814; 2) Uber einen pathologisch-therapeutischen Versuch uber Magensaftabsonderung des Hundes. Arch. f. Verdauungskr., Bd. 3, 1898, S. 488-489].
[28] 28 Больничн. газета Боткина, 1897, стр. 1569. - См. также: Тр. Общ. русск. врачей в СПб., т. 65, сентябрь, 1897, стр. 25--30.
[29] 29 Архив биол. наук, т. V. 1897, стр. 213-224. - [См. также: Zur Frage uber den Ort der Harnstoffbildung bei den Saugetieren]. Archiv f. exper. Pathol. u. Pharmacol., Bd. XXXVIII, 1897, S. 215-222.
[30] 1 30 Schondorff, Pfluger's Archiv f. d. ges. Physiol., Bd. LIV, S. 423.
[31] 2 31 Архив биол. наук, т. IV, 1896, стр. 241.
[32] 32 Киевск. университетск. изв. , 1894, и I. B. M a l y s, 1896, стр. 315.
[33] 33 Ka u f m a n n. Nouvelles recherches sur le lieu de formation de Turee dans l'organisme animal. Role preponderant du fole dans cette formation. Archive de physiol., v. XXVI, p. 531-546; Jahresber. f. Tierchemie, 1895, S. 172.
[34] 2 34 Судя по предварительному сообщению Шендорфа, мышцы содержат мочевину в количествах, не допускающих отнесения ее на счет крови, пропитывающей мышечную ткань (Pfluger's Archiv, 1895).
[35] 35 Ср.: Mintzer, Archiv f. exper. Pathol. S. 197; Richter, Berl. klin. Wochenschr., Bd. XXXIII, 1896, S. 454.
[36] 2 36 Архив биол. наук, т. IV, 1896, стр. 212. (См. первую книгу этого тома, стр. 287. - Ред. ).
[37] 3 37 L. cit. , S. 193.
[38] 1 38 Архив биол. наук, т. I, 1892, стр. 400.
[39] 39 Magnamini. Le modification del recombrio azotato dopo Timnesto della vena porta colla vena cava inferiore. II Policlinico, v. III, 1896, p. 11.
[40] 2 40 2 Queirolo, Untersuch. z. Naturlehre d. Mensch, u. d. Tiere, Bd. XV, 1895.
[41] 41 Lieblein, Archiv f. exper. Pathol. u. Pharmakol., Bd. XXXIII, S. 332.
[42] 2 42 Архив биол. наук, т. III, [1894].
[43] 3 43 Там же, т. II, [1893].
[44] 44 Там же, т. I, 1892, стр. 400. (См. первую книгу этого тома, стр. 210. - Ред. ).
[45] 45 Mi n t z e r, Archiv f. exper. Pathol. u. Pharmacol., Bd. XXXIII, S. 182.
[46] 46 Речь, читанная в торжественном заседании Общества русских врачей в память С. П. Боткина, 1899 г. [Больн. газ., 1900. № 14--15, стр. 617630; № 16, стр. 703-709; № 17, стр. 752-756; № 18, стр. 804-811.См. также: Труды Общ. русск. врачей, т. 67, 1900, ноябрь-декабрь, стр. 197-242].
[47] 47 Эти опыты были начаты доктором Д. Л. Глинским и продолжались доктором С. Г. Вульфсоном на собаках, у которых нормальные концы слюнных протоков были выведены предварительной операцией из полости рта на кожу. Во время опытов на коже около отверстия протока приклеивалась воронка с подвешенным цилиндриком. Слюна подчелюстной и подъязычной (слизистых) желез собиралась вместе, слюна околоушной - отдельно.
[48] 48 Операция эта не очень легка. После нескольких попыток мы остановились на следующем приеме: отверстие желчного протока с куском слизистой оболочки выводится из полости кишки на ее поверхность и здесь пришивается, а затем эта кишка соответственным местом укрепляется и приживляется в отверстии брюшной раны.
[49] 49 Автор несколько раз ранее делал сообщения о ниже упоминаемом исследовании.
[50] 1 50 См. том 1, стр. 536. -- Ред.
[51] 51 [Die physiologische Chirurgie des Verdauungskanals]. Ergebn. d. Physiol... I. Jarhrg., 1. Abteil., 1902, [S. 246 284].
[52] 1 52 Данные, стоящие в прямых скобках, принадлежат Редакции. - Ред.
[53] Тр. 53 Общ. русск. врачей в СПб., т. 69, март- май, 1902, стр. 405 417. - Бо - Больничн, газета Боткина, № 33, 1902, [стр. 1481-1487].
[54] 54 Тр. Общ. русск. врачей в СПб., т. 70, сентябрь октябрь, 1902, стр. 139-143. - Больничн. газета Боткина, 1902, [№ 47, стр. 2153 2155].
[55] 55 Les prix Nobel en 1904. Stockholm, 1906.
[56] 56 Тр. Общ. русск. врачей в СПб., т. 71, март-май, 1904, стр. 39-54.
[57] 57 Изв. Военно-медиц. акад. , т. IX, 1904, сентябрь, стр. 3. [См. также: Uber die ein und demselben Eiweissfermente zukommende proteolytische und milchkoagulierende Wirkung verschiedener Verdauungssafte Zeitschr. f. physiol. Chemie, Bd. XLII, 1904, S. 415 452.
[58] 58 [Glassner], Beitrage zur chemischen Physiologie und Pathologie, Bd. I, 1901.
[59] 59 [Loevenhart], Zeitschr. f. physiol. Chemie, Bd. XLI.
[60] 2 60 Диссертация, СПб., 1901.
[61] 61 Zeitschr. f. physiol. Chemie, Bd. XXXII, [1901, S. 291-319]. 19).
[62] 2 62 [Pekelharing], ibidem, Bd. XXXV, [1902, S. 8 30].
[63] 63 3 Тр. Общ. русск. врачей в СПб. за 1900-1901 гг. и протокол заседания 21 января 1901 г.
[64] 1 64 [Die aussere Arbeit der Verdauungsdrusen und ihr Mechanismus). Nagel's Handbuch d. Physiol. d. Mensch. , Bd. II, 1. H., 1906 -1907, [S. 666 -743]. Классическое описание секреторных процессов дал Гейденгайн в «L. Hermann's Handbuch der Physiologie" (1880). В «Schaftr's Textbook of Physiology» (1893) имеется в высшей степени продуманное и обширное описание Ленгли физиологии слюнных желез. Как первая, так и вторая статья, особенно последняя, снабжены богатыми литературными указаниями. Результаты нашей лаборатории о работе пищеварительных желез собраны в моей книге «Работа пищеварительных желее» (1898), французский перевод которой появился в 1901 г., а английский - в 1902 г.
[65] Тр. 65 Общ. русск. врачей в СПб., 1895.
[66] 2 66 Диссертация, СПб., 1898.
[67] 67 3 Диссертация, СПб., 1901.
[68] 68 [M a l l o i z e l], Journ. de physiol. et pathol. gener., 1902, [t. 4, p. 547, 749, 926].
[69] 69 5 Диссертация, СПб., 1904.
[70] 6 70 Диссертация, СПб., 1904.
[71] 71 [Billard et Dieulafe], Compt, rend. de lac. de biol. a Paris, 1902.
[72] Русский. 72 врач. 1903, [№ 23, стр. 869--872].
[73] 73 Forhandlingar vid Nord. Naturforsekareoch Lakaremotet. Helsingfors, 1909.
[74] 74 [L u d w i gl, Zeitschr. f. rat. Med. , N. F., L., 1851, [S. a55-277].
[75] 75 2 [S c h i f f], Archiv f. physiol. Heilkunde, 1851.
[76] 3 76 [Cl. Bernard]. Lecons sur la physiologie et la pathologie du systeme nerveux, t. II, 1858.
[77] 77 [E c k h a r d], Eckhard's Beitrage... 2, 1860, [S. 81-98].
[78] 5 78 L. cit.
[79] 6 79 [S c h i f f]. Lehrbuch der Muskel- und Nervenphysiologie. 1858-1859.
[80] 7 80 [N a v r o t z k yl, Studien d. Physiol. Inst. zu Breslau, Bd. 4, 1868, [H. 4, S. 125-145].
[81] 8 81 [Loeb], Eckhard's Beitrage, V, 1869, [S. 1-26].
[82] 9 82 [He i d e n h a i n], Pfleger's Archiv f. d. ges. Physiol., Be. XVII, 1878. [S. 1-67].
[83] 83 [L a n g l e yl, Philosophic. Transact. of the Royal Soc. (в дальнейшем: Phil. Trans. ), London, 1892, [p. 85--124].
[84] 84 [H e i d e n h a i n], Stud. d. Physiol. Inst. zu Breslau, 1868, [Bd. 4, Н. 4, S. 1-124], и Pfluger's Archiv f. d. ges. Physiol., Bd. XVII, 1878, [S. 1-67].
[85] 2 85 [Werther], Prliger's Archiv f. d. ges. Physiol., Bd. XXXVIII, 1886, S. 293 -311].
[86] 3 86 [Fletcher], Phil. Trans. , 1889, [vol. 180, p. 109--154].
[87] 4 87 [He i d e n h a i n], l. cit.
[88] 88 [He i d e n h a i n], l. cit.
[89] 1 89 [Becher und Ludwigl, Zeitschr. f. rat. Med. , N. F., I, 1851, [S. 278 284].
[90] 90 [H e i d e n h a i n], Pflugers Archiv f. d. gen. Physiol., Bd. XVII, 1878, [S. 1-67].
[91] 2 91 [L a n g l e y], Phil. Trans. , 1892, [р. 85--124].
[92] 92 [Zerner], Mediz. Jahrbucher, 1887, [S. 530-538].
[93] 1 93 [L u d w i gl, Sitzungsber. d. Wien. Akad., mathem.-nature. Kl., XXV, [S. 584 590], и Wien. med. Wochenschr., 1860, [№ 28 -29, стлб. 433-437, 449-454].
[94] 2 94 [B a y l i s s and Hill, Journ. of Physiol., vol. 16, 1904, [p. 351-359].
[95] 95 [B a y l i s s and B r a d f o r dj, Proceed. of the Royal Soc. , London, 1886, [vol. XL, p. 203-206].
[96] 2 96 [Bradford], Journ. of Physiol., vol. 8, 1887, [р. 86-98].
[97] 3 97 [H e i d e n h a i n], Stud. d. Physiol. Inst. zu Breslau, Bd. 4, 1868, [H. 4, S. 1-124].
[98] 98 [L a n g l e y], Journ, of Physiol., vol. 2, [1879-1880, р. 261-280j.
[99] 1 99 [He i d e n h a i n], 1. cit.
[100] 2 100 2 Врач, 1890, [№ 7, стр. 153-156; № 9, стр. 210-212; № 10, стр. 231--234]. (См. первую книгу этого тома, сто. 142. -- Ред. ).
[101] 3 3 101 Диссертация, СПб., 1890.
[102] 102 [Henderson], Americ. Journ. of Physiol., vol. 3, 1900, [р. 19-25].
[103] 103 a r c r o f t ), Journ. of Physiol., vol. 27, 1901, [p. 31-47].
[104] 6 104 [Ke u c h e l], Диссертация, Юрьев, 1878.
[105] 7 105 [L a n g l e y]. Journ, of Physiol., vol. 9, 1888.
[106] 1 106 [B a r c r o f t], 1. cit.
[107] 2 107 [Cl. Bernardl, Compt. rend. de la Soc. de biol. a Paris, 1858.
[108] 108 3 [H e i d e n h a i n], Pfliger's Archiv f. d. ges. Physiol., Bd. V, 1872, ip. 309 -318].
[109] 4 109 [B e i n b r i d g el, Journ. of Physiol., vol. 26, 1900--1901, [р. 79-91].
[110] 5 110 [L a n g l e y], ibidem, vol. 11, 1890, [p. 123 158].
[111] 6 111 [Cl. Bernard], Journ. de Tanat. et physiol., t. I, 1864, [p. 507-513].
[112] 112 7 [L a n g l e yl, Journ. of Physiol., vol. 6, 1885, [p. 71-92].
[113] 113 [H e i d e n h a i n), Stud. d. Physiol. Inst. zu Breslau, Bd. 4, 1868, [H. 4, S. 1-124].
[114] 2 114 [E c k h a r d], Eckhard's Beitrage. . . Bd. 2, 1860, [S. 205--216].
[115] 115 3 [L a n g l e y], Journ. of Physiol., vol. 1, 1878, [p. 96--103].
[116] 4 116 [Heidenhain], l. cit.
[117] 117 [L a n g l e y], Schafer's Textbook of Physiol., 1898, [vol. I, p. 475-530'
[118] 6 118 [Bradford], Journ. of Physiol., vol. 8, 1887, [p. 86 -98].
[119] 1 119 [H e i d e n h a i nl, Pfluger's Archiv f. d. gen. Physiol., Bd. V, 1872, [S. 143-153].
[120] 2 120 [B a i n b r i d g el, Journ. of Physiol., vol. 26, (1900-1901), [р. 79-91].
[121] 2 121 [L a n g l e yl, ibidem, vol. 11, 1890.
[122] 122 [B r a d f o r d], ibidem, vol. 9, 1888, [p. 287--316].
[123] 1 123 [He i d e n h a i n], l. cit.
[124] 124 [L a n g l e y], Journ. of Physiol., vol. 1, 1878, [p. 96-103].
[125] 3 125 [L a n g l e yl. Journ. of Physiol., vol. 10, 1889, [p. 291-328].
[126] 3 126 [M a t h e w s], Americ. Journ. of
[127] 1 127 [B u n c h], ibidem, vol. 26 (1900 Physiol., vol. 4, 1900, ip. 482-489].
[128] 1 128 [He i d e n h a i n), Pfluger's Archiv f. d. gen. Physiol., Bd. XVII, 1878, [S. 1-67].
[129] 2 129 [Langley], Journ, of Physiol., vol. 10, 1889, [p. 291-328].
[130] 130 3 [M a t h e w s], Annal. of the New-York Acad. of Sci. , vol. II, 1898, и Americ. Journ. of Physiol., vol. 4, 1900, [p. 482 499].
[131] 131 [He i d e n h a i n], Pflugers Archiv f. d. gen. Physiol., Bd. XVII, 1878, 1-67].
[132] 132 [M a l l o i z e l], 1. cit.
[133] 2 133 [C z e r m a k], Sitzungsber. d. Wien. Acad., mathem.-naturw. Kl., XXV, 1857, [S. 3-18].
[134] 134 [Eckhard], Eckhard's Beitrage... 2, 1860, [S. 81-98].
[135] 2 135 [Heidenhain], Pfliger's Archiv f. d. ges. Physiol., Bd. XVII, 1878.
[136] 3 136 [L a n g l e y], Journ. of Physiol., vol. 1, 1878, [p. 339. --369].
[137] 137 [Bradford], ibidem, vol. 9, 1888, [p. 287-316].
[138] 1 138 [Heidenhain, in:] Hermann. Handbuch der Physiologie. 1880.
[139] 2 139 Врач, 1890, [№ 7, стр. 153--156; 9, стр. 210-212; № 10, стр. 231-234].
[140] 3 140 [B r a d f o r dl, l. cit.
[141] 141 [Langley], Journ. of Physiol., vol. 6, 1885, [p. 71-92].
[142] 5 142 Диссертация, СПб., 1891.
[143] 1 143 [Cl. Bernard), цитировано по Langley.
[144] 2 2 144 Melang. biol. Acad. imp. d. sc. de St.-Petersbourg, 8, 1872, [стр. 651665].
[145] 3 145 [B u f f], Eckhard's Beitrage. . . . 12, 1888, [S. 3 -39].
[146] 4 146 Диссертация, СПб., 1894.
[147] 147 [Heidenhain, in:] Hermann. Handbuch der Physiologie. 1880.
[148] 148 2 Pfluger's Archiv f. d. ges. Physiol., Bd. XVI, 1878. (См. первую книгу этого тома, стр. 9. -- Ред. ).
[149] 1 149 Диссертация, СПб., 1904.
[150] 1 150 L. cit.
[151] 1 151 Диссертация, СПб., 1901.
[152] 2 152 [Cl. Bernard], Compt. rend. , 1852,
[153] 1 153 [Schiff]. Lecons sur la physiologie de la digestion. 1867.
[154] 2 154 [L a n g l e y and Anderson l, Journ. of Physics., vol. 16, 1894, [p. 410 440].
[155] 155 [Cl. B e r n a r d]. Lecons sur la physiologie et la pathologie du systeme nerveux. 1858.
[156] 156 [L o e b], Eckhard's Beitrage. . 5, 5, 1870, [S. 73-145].
[157] 5 157 [Grutzner und Chlapowski], Pfliger's Archiv f. d. ges. Physiol., Bd. VII, 1873, [S. 522-529].
[158] 6 158 [Lepine], Gaz, med. de Paris, 1875, [t. IV, № 27, р. 332-333; № 35, р. 436-437].
[159] 1 159 [B o c h e f o n t a i n e], Archives de physiol. norm. et pathol., 1876, [2 ser., t. 3, p. 140 -172].
[160] 2 160 Neurolog. Centralbl., Bd. 7, 1888, [S. 505-509].
[161] 3 161 [E c k h a r d], ibidem, Bd. 8, 1889.
[162] 4 162 [F l e c k], Dissertation, Giessen, 1889.
[163] Невролог. 163 вестн., 1899, [т. 7, № 4, стр. 1--6].
[164] 6 164 Диссертация, СПб., 1900.
[165] 165 Verhandl. d. Kongr. in Helsingfors, 1902.
[166] 2 166 Диссертация, СПб., 1904.
[167] 167 Изв. Общ. естествоиспыт. в Москве, т. 16, [1843, стр. 315].
[168] 2 168 [B o n d l o t]. Traite analytique de la digestion. 1843.
[169] 169 [H e i d e n h a i nj, Pfluger's Archiv f. d. ges. Physicl., Bd. XIX, 1879, [S. 169-171].
[170] 170 Тр. Общ. русск. врачей в СПб., 1894, [март, сто. 24--28].
[171] 1 171 [Fremont], Bull. de I'Acad. de med. , Paris, 1895, [t. XXXIV, p. 509].
[172] 2 172 Archiv f. Anat. u. Physiol., 1895. (Предварительное сообщение см.: Centralbl. f. Physiol., 1889, [Bd. III, № 6, S. 113-114]. (См. первую книгу этого тома, стр. 138 и 175. --Ред.).
[173] 1 173 Тр. Общ. русск. врачей в СПб., 1900, [сентябрь октябрь, стр. 197--242].
[174] Archiv 174 f. Anat. u. Physiol., 1894, [S. 58-82]. - Диссертация, СПб., 1889.
[175] 1 175 Диссертация, СПб., 1891.
[176] 2 176 [Sc h z], Zeitschr. f. physiol. Chemie, Bd. IX, 1890, [S. 577-590].
[177] 3 177 Диссертация, СПб., 1890.
[178] 178 Zeitschr. f. physiol. Chemie, Bd. XXXV, [1902, S. 8-30].
[179] 179 Диссертация, СПб., 1902.
[180] 180 Архив биол. наук, т. III. 1894, [стр. 453-515].
[181] 181 L. cit.
[182] 182 [Bidder und Schmidt]. Die Verdauungssafte. 1852.
[183] 183 Verhandl, d. Kongr. in Helsingfors, 1902.
[184] 2 184 Диссертация, СПб., 1903.
[185] 185 Еженед. клинич. газета Боткина, 1897, [№ 41, стр. 1569--1573]. (См. эту книгу, стр. 226. - Ред. ).
[186] 186 Архив биол. наук, т. V, 1896, [стр. 417--519].
[187] 1 187 Диссертация, СПб., 1904.
[188] 2 188 L. cit.
[189] 1 189 [Schiff]. Lecons sur la digestion. 1867.
[190] 2 190 Еженед. клинич. газета Боткина, 1897, [№ 22, стр. 809-814].
[191] 3 191 [L e c o n t e], La Cellule, 17, 1900, [p. 284--318].
[192] 192 [Leconte], l. cit.
[193] Тр. 193 Общ. русск. врачей в СПб., 1904, [год 71-й, март- май. стр. 5-111.
[194] 1 194 Диссертация, СПб., 1898.
[195] 195 Диссертация, СПб., 1899.
[196] 2 196 Диссертация, СПб., 1900.
[197] 197 cit.
[198] Pfliger's 198 Archiv f. d. ges. Physiol., Bd. LXXXIV, 1901, [S. 101-114].
[199] 2 199 L. cit.
[200] 200 [Meisl], Wien. klin. Rundschau (1903 и. 1904).
[201] 2 201 Обозрение психиатрии, 1899.
[202] 202 L. cit.
[203] 203 Диссертация, СПб., 1882.
[204] 204 [Axenfeld], Atti e rendic. della Acad. med. -chirurg. di Perugia, 1890, [vol. 2, parte 1, p. 142-176].
[205] 2 205 2 [C o n t e j e a n]. These de Paris. 1892.
[206] 206 [S c h n e y e r], Deutsche med. Wochenschr., 1896, [№ 12, S. 173].
[207] 207 Архив биол. наук, 1896, [т. IV, стр. 425-448].
[208] 208 Архив биол наук, т. I, 1892, [стр. 586-709].
[209] 209 Русск. врач, 1903, [№ 32, стр. 1121-1123].
[210] 210 [Hornborg] Skand. Archiv. f. Physiol., Bd. 15, 1904, [S. 209-258].
[211] 108). 211 Тр. Общ. русск. врачей в СПб. (1903-1904), [год 71-й, стр. 95
[212] L. 212 cit.
[213] 2 213 [Arthusj, Compt. rend. de la Soc. de biol. a Paris, 1903, [t. 55, p. 475-476].
[214] 3 214 Русск. врач, 1903.
[215] 215 L. cit.
[216] 216 L. cit.
[217] L. 217 ait.
[218] 1 218 Тр. С.-Петербургск, общ. естествоиспыт., т. XI, 1880. (См. первую книгу этого тома, стр. 88. - Ред. ).
[219] 2 219 [Heidenhain, in:] Hermann. Handbuch der Physiologie. 1880.
[220] 220 [Fodera, in:] Moleschott, Untersuch. z. Naturlehre d. Mensch. u. d. Thiere, Bd. XVI, 1896, [S. 79-89].
[221] 221 [Frouin], Compt. rend. de I'Acad. d. Sc., 1902, [p. 890-893].
[222] 2 222 Диссертация, СПб., 1899.
[223] [Bernstein]. 223 Berichte d. Sachsisch. Ges. d. Wiss. zu Leipzig, 1869.
[224] 2 224 [Heidenhain, in:] Hermann. Handbuch der Physiologie. 1880.
[225] 225 3 Диссертация, СПб., 1888.
[226] 4 226 Архив биол. наук, 1899, [т. VII, стр. 1-86].
[227] 1 227 Диссертация, СПб., 1901.
[228] 2 228 Archiv f. exper. Pathol. u. Pharmacol., Bd. XX, [1886, S. 367--384].
[229] 1 229 [Delezennel Compt. rend. de la Soc. de biol. a Paris (1902 et 1903), [t. 54 et t. 55].
[230] 230 Диссертация. СПб., 1899.
[231] 231 Архив биол. наук, 1893, [т. II, стр. 432-460].
[232] 2 232 [Gottlieb], Archiv f. exper. Pathol. u. Pharmacol... Bd. XXXIII, 1894, [S. 261--285].
[233] 3 233 Архив биол. наук, 1894, [т. III, стр. 395-421].
[234] 1 234 [Gottlieb], I. cit.
[235] 2 235 Диссертация, СПб., 1896.
[236] 236 [Wertheimer et Lepage], Journ. de physiol. et de pathol. gener., 1901, [t. III, р. 821].
[237] 4 Тр. 237 Общ. русск. врачей в СПб., 1896, [год 63-й, февраль, стр. 7--14].
[238] 5 238 Тр. Конгресса в Гельсингфорсе, 1902. 02.
[239] 239 [CI. Bernard]. Memoire sur le pancreas. 1856.
[240] 2 240 [B ernstein], Berichte d. konigl. Sachsisch. Ges. d. Wiss. zu Leipzig, 1869, [Bd. 21, S. 96-131].
[241] 3 241 Pfliger's Archiv f. d. ges. Physiol., Bd. XVI, [1878, S. 173-189]. (См. первую книгу этого тома, стр. 49. - Ред. ).
[242] 4 242 [H e i d e n h a i n], Pfluger's Archiv f. d. ges. Physiol., Bd. X. [1875, S. 557-632].
[243] 5 243 Archiv f. Anat. u. Physiol., 1893, [Suppl. Bd., S. 176 -200]. (См. первую книгу этого тома, стр. 96. - Ред. ).
[244] 1 244 Диссертация, СПб., 1896.
[245] 2 245 [Wertheimeret Lepage], Compt. rend. de la Soc. de biol. a Paris, 1901, [t. 53, p. 879-880].
[246] 246 Протоколы Конгресса в Гельсингфорсе, 1902.
[247] 2 247 Archiv f. Anat. u. Physiol., 1894, [Physiol. Abt., S. 83-116].
[248] 1 248 Диссертация, СПб., 1896.
[249] 2 249 [Bernstein], l. cit.
[250] 3 250 [CI. Bernard]. Lecons sur les proprietes physiologiques des liquides de lorganisme. 1859.
[251] 251 Клинич. газета Боткина, 1900, [Больн. газета Боткина, № 28, стр. 1272-1288].
[252] 2 2 252 [Wertheimer et Lepage], Journ. de physiol. et de pathol. gener... 1901, [t. III, p. 335-348, 363-374].
[253] 3 253 [B a y l i s s and Starling Journ. of Physiol., vol. 28, 1902, Ip. 325--353].
[254] 254 [E n r i q u e s et H a l l i o n], Compt. rend. de la Soc. de biol. a Paris, 1903.
[255] 255 [Wertheimer], Journ. de physiol. et pathol. gener., 1901, [t. III, p. 689-702, 708-718].
[256] 2 256 [Fleigl, Compt. rend. de la Soc. de biol. a Paris, 1903, [t. 55, p. 462 463].
[257] 3 257 Тр. Общ. русск. врачей в СПб., 1903, [год 71-й, ноябрь-декабрь, стр. 99-103].
[258] 1 258 [Fleig], Journ. de physiol. et pathol. gener., 1904, [t. VI, р. 148].
[259] 259 [Wertheimeret Lepage], l. cit.
[260] 260 Тр. Общ. русск. врачей в СПб., т. 75, март-май, 1908, стр. 456-458.
[261] 261 [Die operative Methodik des Studiums der Verdauungsdrisen]. Tigerstedt's Hande. d. physiol. Methodik, Bd. II, I. Halfte, 2. Abt., 1911, S. 150 -188.
[262] 262 Данные, стоящие в прямых скобках, принадлежат Редакции. - Ред.