
Л. В. Метлицкий, О. Л. Озерецковская
КАК РАСТЕНИЯ
ЗАЩИЩАЮТСЯ ОТ БОЛЕЗНЕЙ

*
Рецензенты:
В. Л. КРЕТОВИЧ, К. В. ПОПКОВА
Издательство «Наука», 1985 г.
ОТ АВТОРОВ
Эта книга рассказывает о защитных силах растений, их способности сопротивляться болезням, о механизмах растительного иммунитета. Признаться, слово «фитоиммунитет» звучит не слишком привычно. «Я знаю иммунитет человека, животных, наконец, дипломатический иммунитет, по растительный?» — однажды пошутил по этому поводу один из лидеров современной медицинской иммунологии академик Р. В. Петров. Говорят, что во всякой шутке есть доля правды. Действительно, фитоиммунитет пока еще не может похвастать столь блистательными успехами, как медицинская иммунология. Однако он существует. Иначе как же объяснить тот факт, что высшие растения, во много раз уступающие микроорганизмам по скорости размножения, пластичности и изменчивости, все же выживают и побеждают в борьбе с паразитами? Ведь сам факт сохранения и развития высших растений свидетельствует о том, что они не так уж и беззащитны по отношению к паразитам и что случаи их гибели от болезней являются скорее исключением, чем правилом. Иначе не могла бы существовать та форма жизни, которая сложилась на нашей планете.
Речь, естественно, идет о дикорастущих, а не о культурных растениях. Что касается последних, то сами условия их культивирования способствуют поражению болезнями. Культурные растения человеку приходится непрерывно защищать. Достаточно вспомнить, сколько используется ежегодно всевозможных пестицидов и других средств защиты растений. А как же в таком случае выживают дикие растения? Ведь их-то никто не защищает, а они гораздо слабее поражаются болезнями.
Но если дикорастущие растения не поражаются или слабо поражаются болезнями, то, значит, они обладают по отношению к ним иммунитетом, и тогда невольно возникает ряд вопросов. В чем состоит иммунитет растений и чем он отличается от иммунитета животных? Почему культивирование растений ослабляет их устойчивость, нельзя ли, познав механизмы фитоиммунитета, использовать их для защиты сельскохозяйственных растений от болезней?
Вопросов можно задавать много. Мы поняли, что нужна научно-популярная книга об иммунологии растений. Более того, нам самим захотелось ее написать. Но как и какую? Задача нелегкая, тем более что перед памп лежали блестяще написанные научно-популярные книги академика Р. В. Петрова «Беседы о новой иммунологии»
[1], английского научного обозревателя Д. Уилсона «Тело и антитело»
[2], профессора В. И. Говалло «Парадоксы иммунологии»
[3] и др.
Правда, все они посвящены медицинской иммунологии, которая благодаря своим выдающимся достижениям стала одной из центральных и наиболее увлекательных дисциплин современной биологии. Нам же предстояло написать книгу о фитоиммунитете, который до сих пор еще находится на положении Золушки в иммунологии вообще. Вероятно, одной из причин этому и является то, что фитоиммунологи пока еще мало популяризируют свою область знания. Между тем последние достижения фитоиммунологии не могут не заинтересовать иммунологов человека и животных и могли бы существенно дополнить эволюционные аспекты иммунных систем всего живого.
Нельзя пожаловаться на недостаток монографий по фитоиммунитету, однако среди них почти нет научно-популярных. Известно, что написать научно-популярную книгу гораздо сложнее, чем научную монографию. Правда, авторы научно-популярных книг могут широко пользоваться метафорами, аналогиями, ассоциациями, которые обычно отсутствуют в научных монографиях. Но использовать, их нужно с большой осторожностью, чтобы в погоне за популярностью не допустить упрощений и неточностей. Конечно, даже самая лучшая научно-популярная книга не может заменить научную монографию. И тем не менее научно-популярная литература сейчас привлекает к себе все большее внимание не только широкого круга читателей, но и узкого круга специалистов. Причин для этого немало. С одной стороны, не прекращается процесс дифференциации наук, а с другой, наоборот, стремление к их интеграции. Взять к примеру биологию. Буквально на глазах отпочковывались в самостоятельные и быстро развивающиеся области молекулярная биология, биоорганическая химия, вирусология и другие дисциплины… Вместе с тем развивается физико-химическая биология, призванная шире использовать точные подходы и методы физики и химии для познания механизмов, регулирующих самые интимные процессы, происходящие в ишвом организме.
Часто именно научно-популярные книги привлекают исследователей в смежную область знания, с тем чтобы решать ее проблемы, исходя из возможностей своей специальности. Ведь не случайно многие важные открытия последних лет совершены на стыке наук, на «ничейных землях», т. е. пограничных полосах между отдельными дисциплинами.
Поток научной информации в каждой области знания настолько стремителен, что даже специалисту невозможно за ним угнаться. Недаром говорят, что лишь тот, кто запасся путеводной идеей, может не захлебнуться в океане книг, журналов и статей. А что же тогда делать современному читателю? Научно-популярная книга должна помочь ему в этом, дать первое, самое общее, по одновременно вполне достоверное представление о состоянии знаний в той или иной области.
В написании данной книги мы руководствовались правилом, что читателю не обязательно входить во все детали науки. Цель книги — заинтересовать его нашей проблемой. В какой мере нам это удалось — судить читателю.
Пользуемся случаем выразить свою искреннюю признательность сотрудникам лаборатории иммунитета Института биохимии им. А. Н. Баха АН СССР II. И. Васюковой, Л. И. Чаловой, Г. И. Чалепко, Г. В. Леонтьевой, Т. А. Платоновой, Л. А. Юргаповой, К. А. Караваевой, с которыми нас связывают годы совместного труда всегда приносившие радость и удовлетворение.
ЖИЗНЬ РАСТЕНИЙ ПОД УГРОЗОЙ
Неужели наше счастье,
Наша жизнь от них зависит?
Г. Лонгфелло[4]
Прошло около девяти тысяч лет с того времени, как человек научился возделывать землю. В истории планеты это миг. В истории человечества за это время произошли потрясающие изменения. Нет нужды их перечислять — они хорошо известны. Они происходят на наших глазах, даже в жизни одного поколения людей. Но и сейчас, как и тысячи лет тому назад, все свои основные потребности человек удовлетворяет за счет растений. Между тем над их жизнью, и в первую очередь над жизнью культурных растений, нависла реальная угроза.
В 1970 г. в Лондоне состоялся III Международный конгресс, посвященный науке о пище и технологии ее производства. Программа конгресса была весьма обширной и привлекла ученых многих специальностей. И это неудивительно. Только полноценная пища является залогом здоровья, работоспособности, хорошего настроения, долголетия. Удивительной на первый взгляд была эмблема конгресса — земной шар, SOS-70 (рис. 1).
 Рис. 1. Эмблема III Международного конгресса, посвященного науке о пище и технологии се производства
Рис. 1. Эмблема III Международного конгресса, посвященного науке о пище и технологии се производства
Это начальные буквы трех английских слов: «Save our souls! (Спасите наши души!)». На эмблеме конгресса те же три буквы являлись начальными трех других английских слов, но весьма созвучных первым: «Science of survival (Наука о выживании)». Выжить при недостатке пищи, вследствие чего треть населения нашей планеты недоедает, а треть просто голодает.
Основой современного учения о потребностях человека в тех или иных продуктах является концепция о сбалансированном питании. Согласно этой концепции обеспечение нормальной жизнедеятельности человека возможно только при условии его снабжения определенным количеством энергии и многими самыми различными по химической природе веществами (аминокислотами, витаминами, минеральными соединениями), каждому из которых принадлежит своя специфическая роль в процессе жизнедеятельности. К тому же необходимы запасы продовольствия на случай стихийных катастроф. Человечеству предстоит решить грандиозную и многогранную проблему, включающую социальные, политические, экономические, демографические, медицинские и другие аспекты. Ее успешное решение неразрывно связано с обеспечением надежной защиты сельскохозяйственных растений от болезней, вредителей и сорняков.
Во всех странах мира немало сделано для защиты урожая от потерь. Однако потери урожая от вредителей и болезней продолжают оставаться огромными — на уровне 35–40 и даже 50 %
[5]. Данные мировой статистики интересны и поучительны. Так, средний человек планеты расходует на продукты питания примерно столько ясе, сколько он тратит на защиту растений. «Создается впечатление, что чем интенсивнее ведется борьба с вредителями и болезнями, тем быстрее повышается их вредоносность, усиливается численность вредителей, болезни завоевывают новые территории»
[6].
В чем же дело? Этот вопрос волнует не только специалистов, по и общественность всего мира. Ofi служит предметом острых дискуссий на научных конгрессах, его обсуждение не сходит со страниц специальных журналов И широкой печати.
Потребовалось время, чтобы прийти к неоспоримому выводу — огромные потери уроягая являются делом рук человеческих. Разные точки зрения высказываются по поводу стратегии и тактики защиты урожая от потерь. Но все согласны с тем, что интенсификация и концентрация сельскохозяйственного производства одновременно создают и более благоприятные условия для развития возбудителей болезней и вредителей. Тем более важно выяснить истинные причины, способствующие их развитии). Таких причин много.
Вот некоторые из них.
Первая причина состоит в том, что освоение новых территорий, продвижение земледелия в районы, где оно ранее не развивалось, приводит к разрушению исторически сложившихся в природе биологических сообществ, называемых биоценозами. В их число входит все живое на Земле, где каждый вид животных и растений в ходе эволюции настолько приспособились друг к другу, что существование одного вида зависит от другого, а всех вместе — от взаимодействия с окружающей средой. Вполне понятно, что в случае исчезновения в биоценозе одного представителя неизбежно возникают благоприятные условия для развития другого, который может оказаться весьма опасным для сообщества в целом. В определенной мере это касается и возбудителей болезней, которых известный зоолог В. Н. Беклемишев на основании анализа большого числа данных рассматривает как необходимых членов биоценозов. Их роль состоит в регулировании численности нормальных членов сложившихся биоценозов и в их защите от вторжения чуждых элементов.
Не всегда можно быстро разобраться в последствиях, связанных с нарушением мирного сосуществования биоценозов. Так, по данным известного энтомолога Г. Я. Бей-Биенко, в целинной степи обитают 312 видов насекомых, среди которых преобладают 38. На возделанном же пшеничном ноле насчитывается 135 видов, а доминантных только 19. Казалось бы, что для защиты пшеницы от вредителей это хорошо. По зато численность отдельных насекомых, личинок и куколок на одни квадратный метр поразительно разная: на целине —106, а на засеянной пашне — 332. И далее, если на целине, например, зерновая совка не причиняет вреда диким злаковым травам, то после посева пшеницы ее численность настолько сильно возрастает, что она становится одним из наиболее опасных вредителей этой ценнейшей сельскохозяйственной культуры.
Другой пример. В природных биоценозах растения мало страдают от корневых гнилей. Однако на пахотных землях корневые гнили, вызываемые чаще всего грибами, стали весьма опасными болезнями многих сельскохозяйственных растений. Фитопатологи их даже так и называют «грибами пахотных земель». Усиление их пара-янтарных свойств происходит за счет ослабления содержащихся в почве сапрофитных микроорганизмов.
Одно сорное растение может поглотить в 2 раза больше азота и фосфора, в 4 раза больше калия и воды, чем культурное. И хотя на долю сорняков приходится относительно малая часть всех растений, они могут создавать серьезную угрозу сельскому хозяйству. Сорняки не только снижают урожай, но являются очагами распространения многих вредителей и возбудителей болезней. Так, пырей и другие дикие злаки служат переносчиками возбудителей желтой и стеблевой ржавчины, мучнистой росы, корневых гнилей.
Вторая причина связана с постепенным сокращением числа растений, используемых сельским хозяйством. Из 300 видов растений, окультуренных человеком в разные времена для продовольственных целей, в настоящее время только 12 видов составляют 75 % всего мирового производства сельскохозяйственной продукции. Нужно сказать, что в древности пища людей была значительно более разнообразней. Теперешнее единообразие не оставляет возможности для эволюции. Чрезмерная стандартизация организмов со временем наказывается вымиранием. Замечено, что с каждым исчезающим видом гибнут десятки видов других организмов, что опять-таки приводит к серьезным нарушениям в биоценозах. Чем больше видов входит в состав биоценоза, тем шире их генетическое разнообразие, тем больше возможностей для приспособления к условиям внешней среды и тем выше устойчивость каждого вида.
Третьей причиной является посев семенами одного сорта на больших площадях для унификации агротехники, для получения однородного по качеству урожая. Для унификации методов его хранения и промышленной переработки это очень важно. Однако генетическое однообразие приводит к тому, что возбудители болезней довольно быстро преодолевают устойчивость растений к ним. В результате новые сорта растений, которые еще сравнительно недавно сохраняли свою устойчивость к возбудителям болезней в течение десятилетий, начинают поражаться ими уже через 5–7 лет. Например, средняя полезная жизнь сорта, устойчивого к стеблевой ржавчине, сейчас составляет не более 5 лет, после чего он начинает поражаться новой расой возбудителя этой болезни.
Четвертая причина состоит в том, что с расширением сельскохозяйственного производства происходит продвижение новых видов и сортов растении в новые районы, расположенные далеко от центров, где формировались их дикие предки. Попадая в новые районы, эти сорта чаще всего уступают по своей устойчивости своим предкам. С продвижением растений в новые районы в них проникают и новые для этих мест вредители и болезни (например, появление в СССР колорадского жука, американской белой бабочки и др.). В результате возникают опустошительные эпифитотии (эпидемии в растениеводстве).
Пожалуй, наиболее впечатляющим примером опустошительности эпифитотий был фитофтороз картофеля, заболевание которым повлияло на судьбу целой нации. Недаром тот, кто впервые описал это заболевание, — немецкий ботаник А. де Бари — назвал ее фитофторой, что означает «пожиратель растений». До 1840 г. фитофтора отсутствовала в Европе. Ее родина Мексика, откуда она в начале 1840-х годов и была завезена в Европу. Вначале это были только отдельные вспышки болезни в разных частях Франции, Бельгии и Англии, поскольку засушливое лето 1844 г. не благоприятствовало распространению болезни. Зато следующие 1845–1846 гг. стали трагическими для судеб народов Северной Европы, и особенно Ирландии, где картофель составлял основной продукт питания. а 2 млн. ирландцев питались исключительно картофелем. Размеры бедствия стали ясны к августу 1846 г., когда все растения на многих картофельных полях были уничтожены. Начался массовый голод, от которого погибли более миллиона человек, тогда как два других миллиона эмигрировали за океан.
Пятая причина связана с непрерывным повышением пищевой полноценности культурных растений, что достигается с помощью селекции, интродукции, агротехники. Но все, что полезно для человека и животных, в такой же мере полезно и для возбудителей болезней и вредителей растений. Благодаря повышению питательности растений паразитирующие организмы могут удовлетворять свои потребности в пище при меньших затратах энергии и использовать высвобождающиеся резервы для еще более быстрого размножения и повышения своей жизнедеятельности.
Показательны в этом отношении результаты селекции некоторых злаковых растений на повышение в них содержания так называемых незаменимых аминокислот, т. е. тех, которые человек и животные самостоятельно синтезировать не могут и должны получать в готовом виде вместе с пищей. А многие растения, весьма богатые по общему содержанию белков, содержат мало незаменимых аминокислот. В результате отсутствия хотя бы одной из них в организме может прекратиться синтез ряда белков, играющих первостепенную роль в нормальной жизнедеятельности. Не зря белки называют протеинами (от греческого протос — первостепенный). По этой же причине многие селекционеры сосредоточили свои усилия на повышении содержания в злаках незаменимых аминокислот, и в первую очередь наиболее дефицитной — лизина. Но стоило только добиться первых положительных результатов, как вскоре обнаружилось, что сорта, более богатые лизином, стали и более уязвимыми для паразитов. Дошло до того, что недавно был даже предложен метод определения незаменимых аминокислот в зерне по анализу личиночных шкурок мучного хрущака, питающегося этим зерном.
Селекция на качество заставляет также «убирать» из состава растений так называемые вторичные метаболиты (алкалоиды, гликозиды, фенолы), которые хотя и ухудшают питательные и вкусовые свойства урожая, но вместе с тем являются факторами его устойчивости к болезням.
Можно указать и на другие причины высоких потерь урожая, и совсем не обязательно в названной последовательности. Но даже из того, что сказано, вытекают важные практические выводы. И важнейшими из них являются два, неразрывно между собой связанные.
Во-первых, вместе с концентрацией и интенсификацией сельского хозяйства возникают одновременно и более благоприятные условия для развития болезней, вредителей и сорняков. В этих условиях многие прежние методы защиты урожая, вполне отвечавшие требованиям небольшого хозяйства, оказываются малоэффективными и даже вовсе неосуществимыми. Нужны новые подходы и методы защиты растений с максимальным, конечно, использованием многовекового опыта земледелия.
Во-вторых, применение новых подходов неизбежно связано с вмешательством в природу. Но при этом надо учитывать не только видимый эффект, но и максимально предвидеть все возможные отрицательные последствия в будущем.
Современная экология — наука о взаимоотношениях организмов и окружающей среды в одинаковой мере отвергает две крайние точки зрения, все еще встречающиеся по данному вопросу. С одной стороны, голый прагматизм, оправдывающий все средства для спасения от голода (нельзя умирать от голода сидя на мешке с золотом во имя будущих поколений). С другой стороны, экологический утопизм, отвергающий химические средства защиты растений и призывающий вернуться назад к природе.
Все очевиднее становится, что за вмешательство в природу надо платить и с ней нельзя вступать в конфликт. Еще до внедрения своего открытия в сельскохозяйственное производство ученому следует знать о том, каким образом оно может повлиять на биосферу в будущем. В этом состоит великая ответственность ученых перед природой и обществом.
ОСНОВОПОЛОЖНИК УЧЕНИЯ
О ФИТОИММУНИТЕТЕ
Если ты выстрелишь в прошлое из пистолета,
будущее выстрелит в тебя из пушки.
Р. Гамзатов[7]
Общеизвестно, что успехи науки XX в. определяются не столько открытиями отдельных талантливых ученых, сколько объединенными усилиями коллективов, способных превратить эти открытия в мощные направления науки и обеспечить их практическое применение. В связи с этим все более важную роль начинают играть ученые, сочетающие в себе талант исследователя и организатора. Таких ученых сравнительно немного, и их имена становятся быстро известными во всем мире. Но даже среди них уникальная роль принадлежит выдающемуся советскому биологу Николаю Ивановичу Вавилову. Именно поэтому ЮНЕСКО приняло решение отметить в 1987 г. во всем мире столетие со дня его рождения.
О Н. И. Вавилове, с именем которого связано становление и развитие многих направлений в биологии, в том числе и фитоиммунологии, написано очень много — писали его соратники, ученики, историки науки, писатели. И тем не менее в книге о фитоиммунологии мы не можем не рассказать о ее основоположнике.
1920 год. Гражданская война. Страна разорена. Не жалея сил, отказывая себе в самом необходимом, народ трудился над восстановлением разрушенного хозяйства. Огромным было стремление народа к науке и культуре, к строительству новой жизни. В такой обстановке собрался III Всероссийский съезд селекционеров. На трибуне 33-летний, но уже хорошо известный профессор. Зал с затаенным дыханием слушает об открытии им параллельной изменчивости у близких видов и родов растений. Свое открытие Николай Иванович назвал законом гомологических рядов в наследственной изменчивости (от греческого «гомос» — соответственный, подобный). Суть закона коротко сводится к тому, что все виды растений, отличаясь друг от друга характерными свойствами, вместе с тем имеют много сходного. Поэтому, обнаружив изменчивость признаков у одного рода или вида, можно ожидать подобную же изменчивость и у других близких нм видов и родов.
Делегаты съезда аплодируют молодому ученому. Его исследования открывают новые, ранее неизвестные пути планомерного поиска иммунных форм среди огромного числа растений, нужных человеку. Вновь открытый закон сравним с периодической системой Д. И. Менделеева, позволяющей предсказывать существование новых химических элементов с определенными свойствами.
Впервые было дано научное обоснование распределению различных форм культурных растений по земному шару. Результаты этих поистине грандиозных по замыслу и масштабам исследований легли в основу фундаментального труда «Учение о происхождении культурных растений после Дарвина». В поисках необходимых для сельского хозяйства растений Николай Иванович исколесил чуть ли не всю планету. Это не был поиск вслепую, а целенаправленные экспедиции по научно обоснованным маршрутам. Он сумел проникнуть в труднодоступные районы Азии, Африки, Южной Америки, не посетив, вероятно, Австралию потому, что она не представляла интереса для поиска нужных для сельского хозяйства растительных форм (так оно и оказалось впоследствии).
В результате Н. И. Вавилов вместе со своими сотрудниками создали самую цепную в мире коллекцию семян, она включает более 250 000 образцов растений, принадлежащих 1740 видам. Коллекция является как бы живой, поскольку непрерывно высевается и изучается более чем в 100 различных по почвенно-климатическим условиям районах Советского Союза. Опа служит неиссякаемым кладезем для создания новых сортов сельскохозяйственных растений, превосходящих старые по урожайности, скороспелости, засухоустойчивости, морозоустойчивости, устойчивости к болезням, пищевым и технологическим достоинствам.
Мы начали свой рассказ о Н. И. Вавилове с его доклада на съезде селекционеров, так как именно селекции в значительной мере были подчинены его исследования в области фитоиммунитета. Придавая большое значение всем способам защиты растений от болезней, он все же считал, что радикальное решение проблемы может быть достигнуто путем широкого использования иммунных сортов.
Теме иммунитета растений, как любимому детищу, Николай Иванович оставался верен всю свою жизнь, несмотря на широкий спектр своих интересов, многогранность деятельности и огромную занятость.
Свои исследования по фитоиммунитету П. И. Вавилов начал с изучения устойчивости хлебных злаков к паразитарным грибам в Петровской, ныне Московской сельскохозяйственной академии им. К. А. Тимирязева. Серию важных исследований он выполнил также в Англии, где ему была предоставлена обширная коллекция пшениц разных видов и сортов. После 8 лет упорного труда, в 1919 г., Н. И. Вавилов опубликовал свою работу «Иммунитет растений к инфекционным заболеваниям», которую посвятил памяти великого исследователя иммунитета Ильи Ильича Мечникова. Ничего подобного в мировой литературе еще не было.
Так были заложены научные основы учения об иммунитете растений к болезням.
Следующая монография, изданная в 1935 г., называлась «Учение об иммунитете растений к инфекционным заболеваниям (применительно к запросам практики)». Интерес к данной проблеме во всем мире непрерывно возрастал. Н. И. Вавилов писал, что огромный фактический материал и специфичность области взаимоотношений растения-хозяина и паразита, а также своеобразная экспериментальная методика в этой области дают все основания к выделению особого раздела ботанической науки — фитоиммунологии, или иммунологии растений.
В 1961 г., спустя 15 лет после смерти ученого, были впервые опубликованы рукописные материалы под названием «Законы естественного иммунитета растений к инфекционным заболеваниям (ключи к нахождению иммунных форм)».
Все работы Н. И. Вавилова по фитоиммунологии теснейшим образом связаны с генетикой, которой ученый самозабвенно служил и не изменял ни при каких обстоятельствах. Именно им были впервые разработаны в нашей стране генетические основы фитоиммунитета. В результате если изучение механизмов устойчивости растений и практического их применения все еще отстает от медицинской иммунологии, то в области генетики иммунитета медицинская иммунология давно широко пользуется достижениями фитоиммунологии.
В этих небольших заметках нет возможности даже кратко изложить учение Н. И. Вавилова об иммунитете. Да и в этом нет особой необходимости, поскольку к нему еще не раз придется вернуться в дальнейшем. Гораздо важнее, по-видимому, попытаться здесь отразить его влияние на развитие современной фитоиммунологии. А оно и сейчас очень велико и плодотворно. Можно привести немало примеров о высоком даре ученого научно предвидеть, о его умении по достоинству оценить и увидеть перспективность тех или иных направлений исследований.
Н. И. Вавилов установил, что первой и основной закономерностью, определяющей существование видов и сортов, иммунных к тому или иному паразиту, является специализация паразитов, приуроченность их к определенному кругу растений. В эволюции же паразитизма явление специализации является основным. Поэтому Н. И. Вавилов призывал селекционеров, прежде чем приступить к селекции на иммунитет, познавать биологию паразитов, степень их специализации. Чем уже специализация паразита по родам и видам растений, тем больше шансов на нахождение иммунных форм в пределах отдельных видов.
Четкая сбалансированность отношений между растением-хозяином и паразитом выражена далеко не всегда и не везде. Она возникает лишь в районах, являющихся совместной родиной растения и его патогена, в результате их так называемой сопряженной эволюции. Именно в таких районах и были обнаружены растительные формы, иммунные ко многим болезням, как это и предсказывал И. И. Вавилов. Иными словами, для того чтобы произошла взаимная притирка, растение и его паразит должны длительное время сосуществовать.
Когда Н. И. Вавилов проводил свои исследования о природе фитоиммунитета, высказывались разные, порой диаметрально противоположные точки зрения. Их авторы часто старались связать иммунитет
с одним каким-то свойством растения, отметая другие. Н. И. Вавилов, внимательно разобрав их, убедительно показал, что иммунитет представляет собой «сумму из множества слагаемых». Такое определение фитоиммунитета, ставшее крылатым, стимулировало проведение фитоиммунологических исследований в самых разных направлениях и привлекло к ним специалистов разного профиля — фитопатологов, биохимиков, генетиков, физиологов.
И еще пример научной проницательности II. И. Вавилова. сыгравший важную роль в развитии современной фитоиммуиологии. Известно, что приобретенный иммунитет, играющий решающую роль в защите человека и животных от болезней, очень ограниченно используется применительно к растениям и, по существу, еще не имеет практического значения для их защиты от болезней; А. в те времена даже сама возможность приобретенного иммунитета растений отрицалась.
Полвека тому назад известный американский фитопатолог Честер на основании обобщения результатов выполненных к тому времени исследований пришел к выводу, что доказательства приобретенного иммунитета у растений слабые, путаные и полны противоречий. Диаметрально противоположный вывод сделал Н. И. Вавилов, который писал, что отрицать теоретическую возможность и наличие приобретенного иммунитета у растений не приходится. Он придавал большое значение развитию исследований в этом направлении, и его авторитетное мнение несомненно служит и сейчас важным стимулом для проведения исследований по иммунизации растений, ставшим в настоящее время одним из центральных направлений в фитоиммунологии.
Нельзя не сказать хотя бы несколько слов о Николае Ивановиче Вавилове как о человеке, о котором буквально ходили легенды, легенды живые, исходившие от людей, его близко знавших. Необычайно широкими были спектр его интересов, энциклопедические знания и поистине фантастическое трудолюбие. «Жизнь коротка, нужно спешить», — часто повторял он и действительно спешил, не теряя пи дня, ни часа для работы. Большую часть своей жизни он провел в пути, экспедициях. Радушие, простота, жизнерадостность создавали вокруг него обстановку, в которой каждому хорошо работалось и легко дышалось.
Николай Иванович очень любил свою мать и относился к ней с большой нежностью и теплотой. Любил и безмерно восхищался дарованием брата — выдающегося советского физика, президента Академии наук СССР Сергея Ивановича Вавилова.
Николай Иванович был наделен «честной храбростью» и до самых последних дней своей жизни не изменял пи себе самому, ни делу, которому отдал жизнь. Он был рассеян в силу занятости мыслями, очень доверчив и щедр. Так, будучи в Англии, он купил на свои деньги продававшиеся на торгах реликвии, связанные с жизнью Чарлза Дарвина, которые впоследствии составили ядро будущего Дарвиновского музея в Москве.
В 1930 г. в беседе с К. Г. Паустовским Н. И. Вавилов четко сформулировал цель своей жизни: «Мобилизовать растительный капитал всего земного шара и сосредоточить в СССР весь сортовой запас семян, созданный в течение тысячелетий природой и человеком».
Крупнейший ботаник-теоретик и растениевод, выдающийся генетик, селекционер, известный географ и путешественник, организатор науки, президент Всесоюзной академии сельскохозяйственных наук, президент Всесоюзного географического общества, директор Института генетики, бессменный директор любимого и созданного им Всесоюзного института растениеводства, которому обязана своим становлением советская и мировая фитоиммунология, Н. И. Вавилов был и остается примером беззаветного служения науке.
МЕДИЦИНСКАЯ ИММУНОЛОГИЯ
И ИММУНОЛОГИЯ РАСТЕНИИ
После некоторых размышлений я пришел к выводу, что в данный момент к числу самых волнующих дисциплин относится иммунология, точнее именно иммунология сильнее всего привлекает мое внимание тем новым светом, который она проливает на многие актуальные вопросы.
Д. Уилсон[8]
«Беседы о новой иммунологии» — под таким названием вышла в 1976 г. книга одного из лидеров современной иммунологии и ее блестящего популяризатора академика Рэма Викторовича Петрова. Вышла и вскоре стала бестселлером. И это неудивительно. Ведь для того чтобы назвать уже существующую науку новой, в ней должны быть совершены принципиально важные открытия и раскрыты новые перспективы для ее дальнейшего развития. Об этом и идет речь в этой замечательной книге, из которой становится ясным, что иммунология превратилась в одну из наиболее интересных и увлекательных дисциплин всей биологии, да и не только биологии.
Старая иммунология родилась в 1881 г., когда знаменитый французский микробиолог Луи Пастер, вернувшись из отпуска, обнаружил, что забыл вылить культуры возбудителей куриной холеры. Высохшие и перегретые культуры перестали вызывать болезнь у кур, по самым поразительным оказалось то, что птиц, которым вводили эти культуры, уже не удавалось заразить свежей культурой холерных бактерий. Так был создан принцип защиты от болезней, основанный на введении в организм ослабленной культуры ее возбудителей.
Однако Пастер был далек от понимания деталей болезни и создания теории иммунитета. Более того, он думал, что микроорганизмы, находящиеся в составе вакцины, потребляют в макроорганизме некие вещества, в результате чего последний оказывается питательно неполноценным и поэтому не пригодным для последующего заражения. Создание теории иммунитета стало уделом других исследователей.
В конце прошлого века Илья Ильич Мечников, работая в Пастеровском институте в Париже, открыл фагоциты — особые клетки, пожирающие чужеродные включения, и тем самым создал клеточную теорию иммунитета. Чтобы обнаружить фагоциты, достаточно ввести животному кусочки угля. Фагоциты, наглотавшись сажи, становятся темными и хорошо заметными под микроскопом.
Одновременно с Мечниковым Пауль Эрлих разработал гумморальную теорию иммунитета, открыв существование антител, связывающих высокомолекулярные антигены. В течение ряда лег не затихала острая дискуссия между Мечниковым и Эрлихом, в которой каждый отстаивал свою теорию иммунитета. Будущее показало, что оба они оказались правы. В итоге им обоим была присуждена Нобелевская премия за создание клеточной и гумморальной теорий, являющихся венцом старой иммунологии.
Старая иммунология — это наука о невосприимчивости организма к инфекционным болезням. Так ее определил Мечников в конце прошлого века. Такая иммунология, получившая еще название инфекционной, была ориентирована только на клинику. Она родилась из суровой необходимости борьбы, с болезнетворными микробами и просуществовала около 60 лет. Однако несколько десятков лет назад наши представления об иммунологии настолько расширились, что возникла необходимость в пересмотре ранее принятых понятий. Вот тут-то и зародилась новая иммунология.
Повое осмысление иммунологии началось с 1945 г. после публикации работ английского исследователя, лауреата Нобелевской премии Питера Медевара.
Шли тяжелые годы второй мировой войны. Фашисты бомбили Лондон. Медевар работал в это время в одном из госпиталей Лондона, пытаясь пересаживать обожженным людям здоровую донорскую кожу. Чужая кожа не хотела приживаться. Медевар доказал, что причиной этого является иммунная система, которая отторгает несвойственный ей чужеродный трансплантат. В последующие годы стало ясным, насколько тонкие оттенки способна различать иммунная система организма. Последняя срабатывает на чужеродную ткань, отличающуюся от собственной даже по одному гену, т. е. по минимальному признаку.
Таким образом, если раньше иммунную систему рассматривали как защиту организма от инфекции, то после работ Медевара в корне изменились представления об иммунологии. Оказалось, что задача иммунитета значительно шире — уничтожать клетки, генетически отличающиеся от клеток собственного тела. Поскольку микробы для организма также являются чужеродными, то действие иммунитета направлено и против них тоже. Защита от микробов — только частный случай деятельности иммунной системы.
Но зачем организму млекопитающих столь жесткий иммунологический контроль, способный уничтожить клетку, отличающуюся от клеток своего тела лишь одним геном? Для этого должны быть серьезные основания.
Считается, что именно этот вопрос, который наиболее четко сформулировал Нобелевский лауреат Френк Макферлан Бернет в 1964 г., и сделал иммунологию новой. И новая иммунология дала на пего следующий ответ.
В многоклеточном организме при сожительстве генетически идентичных клеток неизбежно должны возникать клетки-мутанты. Известно, что в организме человека, содержащем 10
13 клеток, в каждый данный момент должно быть не менее десяти миллионов клеток-мутантов. Поскольку иммунная система способна отличать чужеродность, заключающуюся только в одном гене, было предположено, что она и несет функцию надзора за генетическим постоянством организма, уничтожая клетки-мутанты. Это значит, что многоклеточные организмы могли возникнуть только под присмотром собственной иммунной системы.
«Иммунитет — это способ защиты организма от живых тел и веществ, несущих на себе признаки чужеродности. В понятие живых тел и веществ, несущих на себе признаки чужеродного генома, могут быть включены бактерии, вирусы, простейшие, черви, белки, ткани, измененные аутоантигены, в том числе и раковые»
[9]. Таким образом, задачи новой иммунологии значительно шире старой. Последняя является лишь частью новой иммунологии, иными словами, иммунология из медицинской дисциплины превратилась в общебиологическую.
Иммунитетом обладает все живое — от амебы до человека. Однако у организмов, находящихся на различных ступенях эволюционной лестницы, он выражен по-разному. Но как бы ни различались механизмы уничтожения чужеродности у организмов, находящихся на различных эволюционных уровнях, всех их объединяет общее свойство — необходимость распознать эту чужеродность перед тем, как ее ликвидировать. Недаром центральная догма иммунитета, сформулированная Бернетом, гласит, что главной функцией иммунных систем служит распознавание своего и чужого. Именно это свойство: распознать чужеродность, чтобы ее уничтожить или отвергнуть, и роднит между собой иммунные системы всего живого. И как будет видно из последующего изложения, высшие растения хорошо вписываются в эту систему, не являясь в ней исключением.
Полученные к настоящему времени результаты позволяют считать, что иммунитет растений контролирует не только устойчивость к паразитарным организмам, но и охраняет их целостность и генетическое постоянство. В его ведении находятся такие явления, как совместимость тканей при прививках; совместимость антигеноз пыльцы и пестика при опылении; реакции, возникающие в ответ на поранение; детоксикация экзо- и эндотоксинов, в частности ксенобиотиков. Однако механизмы иммунитета растений зачастую совершенно непохожи на те, с которыми приходится иметь дело иммунологам животных.
С давних времен люди пытались попять причины болезней растений и найти средства для их излечения. Тем не менее учение о болезнях растений стало оформляться как наука только в начале нашего века. Это и неудивительно, ведь люди в первую очередь пытались избавиться от своих болезней, а болезни растений беспокоили их в меньшей мере.
Еще в 1903 г. один из основоположников учения об иммунитете И. И. Мечников в монографии «Невосприимчивость к инфекционным болезням» посвятил отдельную главу иммунитету растений. Он писал, что в ботанике вопрос о невосприимчивости оставался на заднем плане, так что мы не имеем о нем никаких специальных работ.
Учение об иммунитете растений начало развиваться в тот период, когда основные принципы иммунитета человека и животных были уже установлены. Неудивительно поэтому, что растениеводы пытались идти путями, проложенными медиками, которые уже добились к тому времени выдающихся практических результатов. Тем более что наиболее выдающиеся фитопатологи были медиками по образованию. Однако попытки иммунизировать и вакцинировать растения в надежде на образование у них специфических антител оказались безуспешными. Правда, в ряде случаев удалось получить некоторый защитный эффект, который оказывался локальным и кратковременным.
Неудачи иммунизации растений по образцу животных в значительной мере утомили и разочаровали исследователей. Возможно, поэтому в последующие 50 лет фитоиммунологи преимущественно накапливали факты, главным образом касающиеся тех изменений, которые происходят в заболевшем растении, т. е. занималась фитопатологией (от греческого pathos — страдание, logos — учение) — наукой о страдании растений, а отнюдь не фитоиммунологией. Этот период характеризуется полным несоответствием между потоком частных сообщений и фактическим отсутствием попыток теоретически обобщить этот материал.
К этому времени медицина уже претерпела несколько поколений специализаций: она разделилась на автономные разделы, которые, в свою очередь, стали родоначальниками новых направлений. В составе иммунологии выделились такие самостоятельные разделы, как аллергология, иммунобиология, иммуногенетика, иммунопатология, клиническая иммунология, судебно-медицинская иммунология, иммунология эмбриогенеза, иммунология инфекций, иммунохимия, трансплантационный иммунитет, иммунологическая толерантность и др.
Ничего подобного в фитопатологии до самого последнего времени не происходило. Впрочем, развитием одного направления фитоиммунология все же может похвастаться. Речь идет о генетике фитоиммунттета, которая не только не отставала, но даже обгоняла медицинскую иммуногенетику. «Если при изучении биохимических основ устойчивости растений фитопатологи долгое время шли путями, проложенными медиками, и над ними довлел авторитет медицинской иммунологии… то в области генетики иммунитета фитопатология ушла далеко вперед, и ее достижения используются медицинской и ветеринарной иммунологией»
[10]. Причина заключается в различном отношении к индивидууму у медика и фитопатолога. Медик призван прежде
всего заботиться об индивидууме, т. е. о конкретном человеке, которого лечит. Между тем фитопатолога отдельное растение мало интересует, его объектами является, скорее, популяция особей, принадлежащих виду или сорту. Предметом его заботы является не растение как таковое, а целое поле, что дает ему огромные преимущества перед медиком. Он может скрещивать и заражать неограниченные количества растений, что позволяет ему во много раз быстрее получать результаты и делать необходимые выводы.
Отставание фитоиммунологии во многом объясняется значительными различиями природы иммунитета у растений и животных, которые стали ясными не сразу. Защита человека и животных, как известно, основана на приемах иммунизации и вакцинации, связанных с повышением в организме концентрации иммуноспецифических антител, являющихся продуктом длительной эволюции их организмов. Дифференцировка тканей животных зашла настолько далеко, что у них появились специализированные органы, ткани и клетки, ответственные за иммунную систему. Это такая же самостоятельная система, как нервная, сердечно-сосудистая и др. Иммунная система пронизывает все тело и представляет собой совокупность всех лимфоидных органов и клеток. Составляющими этой системы являются тимус, селезенка, костный мозг, групповые лимфотические фолликулы и, наконец, лимфоциты, которые являются центральной фигурой иммунной системы. Последние по образному выражению Р. В. Петрова являются «вездесущими детективами нашего тела», задача которых обнаружить чужеродное начало и обезвредить его.
В последнее десятилетие удалось обнаружить две независимые группы В- и Т-лимфоцитов, взаимодействие которых и их совместная работа с макрофагами обеспечивает всю гамму иммунологических реакций. Для функционирования лимфоцитов необходима гумморальная система, способная доставить антитела в любую точку тела, где возникает в них необходимость.
Ничего похожего у растений нет, их дифференцировка значительно примитивнее. Растение имеет сосудистую систему флоэмы и ксилемы, а протопласты отдельных клеток сообщаются между собой с помощью систем плазмодесм (ксилема — основная водопроводящая ткань сосудистых растений; флоэма — ткань, проводящая пластические вещества и пространственно связанная с ксилемой в общую проводящую систему; плазмодесмы — плазматические мостики между клетками, обеспечивающие непрерывность цитоплазмы). Специализированных органов иммунной системы у растений нет. Судя по тому, что каждая растительная клетка может при определенных условиях дать начало целому растению, можно полагать, что она несет в себе все функции, в том числе и иммунные. При этих условиях ожидать, что в растительных тканях могут образовываться и функционировать антитела в их классическом понимании, не представляется возможным.
Казалось, что иммунитет растений основывается на неких особых механизмах, не имеющих ничего общего с механизмами животных. Но так было только до тех пор, пока старые понятия иммунологии не были переосмыслены, в результате чего возникла новая иммунология, в корне изменившая наши представления об иммунитете вообще.
С этих позиций фитоиммунитет следовало рассматривать как систему, охраняющую структурную и функциональную целостность растений, основанную на свойстве распознавать и отторгать генетически чужеродную информацию. Растения не имеют иммунокомпетентных лимфоцитов и не способны образовывать антитела, по им, как, впрочем, и всему живому, свойственна способность распознавать генетически чужеродные клетки и молекулы.
КТО, ЧТО И КАК ВЫЗЫВАЕТ БОЛЕЗНИ РАСТЕНИЙ
Вредители и болезни сельскохозяйственных культур стары, как и само сельское хозяйство.
Г. Рассел [11]
ВОЗБУДИТЕЛИ БОЛЕЗНЕЙ РАСТЕНИИ
И ОРУДИЯ ИХ АГРЕССИИ
Трудно представить себе что-нибудь более бескорыстное и трудолюбивое, чем зеленое растение. Растения снабжают нас пищей, энергией, они пополняют запасы кислорода в атмосфере, которые отнюдь не беспредельны, да еще украшают нашу жизнь и излечивают от многих болезней. Выходит, что все наше существование так или иначе связано с иммунитетом растений. По эти уникальные производители, от существования которых зависит жизнь на Земле, сами по себе служат прекрасной мишенью для паразитов.
Известно, что паразитом называется организм, живущий на другом организме и зависящий от него как от источника питания.
К возбудителям инфекционных болезней растений относятся.
1. Паразитические грибы и бактерии. О них речь пойдет особо, так как именно устойчивости растений к ним и посвящена в основном эта книга.
2. Паразитические цветковые растения, живущие за счет других растений, носящих название растений-хозяев. Самым распространенным их представителем является повилика, которая обвивается вокруг стеблей хозяев и с помощью особых присосок высасывает из них питательные вещества. Сюда же относятся заразиха и омела.
3. Паразитические животные или растительные нематоды. Чаще всего — это обитающие в почве круглые черви, называемые фитогельминтами. Они прокалывают растение, проникают в него и питаются его содержимым. Фитогельминты выделяют ферменты, превращающие сложные органические вещества растений в простые, пригодные для усвоения. Некоторые нематоды выделяют биологически активные соединения, способствующие притоку к ним питательных веществ. Вокруг места проникновения таких нематод начинается усиленное деление клеток и образуются утолщения — галлы. Нематоды способны выделять токсические вещества, подавляющие устойчивость к ним со стороны растений. Многие нематоды и растения-паразиты являются переносчиками вирусов, и в этом состоит их большая опасность.
4. Вирусы — инфекционные болезнетворные агенты, находящиеся на грани между веществами и существами. Вирусы составляют вторую по вредоносности после грибов группу патогенов растений. В настоящее время известно около 600 фитопатогенных вирусов.
Кроме инфекционных болезней, у растений широко распространены также болезни неинфекционного характера, т. е. не передающиеся от больного растения к здоровому. Их причиной служат нарушения в нормальном обмене веществ, свойственном растению. Нарушения возникают из-за неблагоприятных условий существования либо старения, отчего в растительных тканях часто накапливается чрезмерно высокое содержание соединений, которые оказывают на клетки токсическое действие.
Все возбудители болезней растений в порядке убывающей вредоносности могут быть расположены в следующий ряд: грибы — вирусы — бактерии — паразитические цветковые растения. Настоящая книга в основном касается устойчивости растений к паразитарным грибам и в меньшей мере — к бактериям. Для этого есть все основания, поскольку грибы сильнее всего поражают растения. Они уносят более 20 % общего урожая мировой продукции. Причем подавляющее число иммунологических исследований выполнено именно на грибах.
По ориентировочным подсчетам, существует не менее 10 000 видов фитопатогенных грибов, причем не исключено, что их количество может быть чуть ли не вдвое больше. Среди бактерий возбудителями болезней растений являются только 150–200 видов. Иными словами, бактерии значительно менее вредоносны для растений, чем грибы. В то же время для теплокровных, в том числе и для человека, наоборот, бактерии значительно вредоноснее. Предполагается, что причиной этому могут быть несколько факторов.
1. Как правило, бактерии термофильны, поэтому организм млекопитающих является для них более подходящим, чем растительные ткани.
2. Бактерии лучше растут в щелочной среде, а грибы предпочитают более кислую. Сок растений обычно имеет слабокислую реакцию.
3. Бактерии лишены способности активно проникать в ткани, тогда как грибы образуют особые структуры, с помощью которых прободают покровы и клеточную стенку. В животный организм бактерии в основном проникают через дыхательные пути и пищеварительный тракт. Растения, защищенные различного рода покровами, малодоступны для проникновения бактерий, в то время как многие виды грибов способны такие покровы преодолевать.
4. Большинство грибов образуют два вида спор, одни из которых служат для распространения, а другие — для выживания в неблагоприятных условиях. Только часть бактерий образует споры, тогда как у остальных для распространения служат сами бактериальные клетки. Бактерии не имеют также специальных органов для перезимовки, благодаря чему их сохранение от одного вегетационного сезона до другого значительно затрудняется.
Предполагается, что все эти трудности, связанные с заражением растений, с распространением и перезимовкой, и определяют главные причины, благодаря которым бактерии как патогены растений уступают грибам по вредоносности.
Для того чтобы стать паразитом, микроорганизм должен приспособиться к своему будущему хозяину, или, как говорят, пройти процесс специализации. Специализация — это приуроченность патогена к одному или нескольким растениям-хозяевам. Различают узко- и широкоспециализированные группы возбудителей болезней. Широкоспециализированные вызывают заболевания различных родов внутри одного семейства растений или даже растений различных семейств. Некоторые возбудители болезней, например возбудитель серой гнили (Botrytis cinerea), заселяют и разрушают без разбора всевозможные ткани и органы различных растений. В силу этого их называют полифагами.
Наоборот, монофаги, или узкоспециализированные патогены, паразитируют только на растениях одного рода или даже одного вида. Внутри вида у таких паразитов часто возникают особые формы, приспособленные только к определенным сортам, которые не отличаются друг от друга во всем, кроме способности поражать одни сорта и не поражать другие. Такие формы получили название специализированных форм или физиологических рас.
Паразиты растений отличаются по характеру своих патогенных свойств. Среди них различают несколько групп.
Факультативные паразиты (необязательные) — наиболее низкооргаиизованные микроорганизмы, которые обычно живут как сапрофиты на мертвых остатках растений, но при определенных благоприятных для них условиях могут нападать на живые растительные ткани, предпочитая при этом старые или ослабленные. Все они полифаги, круг их растений-хозяев достаточно широк. Хорошо растут на питательных средах и очень к ним нетребовательны. Примером является уже упомянутый возбудитель серой гнили многих растений.
Внутри этой группы также существует специализация. Крайней степенью являются «всеядные» паразиты, которые способны поражать сотни видов растений — из самых различных таксономических групп. Часто успех заражения у них определяется не принадлежностью растений к той или иной группе, а тем, ослаблена ли в достаточной мере его ткань. В отличие ют «всеядных» в группу факультативных паразитов входят также «многоядные паразиты», которые имеют уже более ограниченный, но все же достаточно широкий круг хозяев.
Факультативные сапрофиты в отличие от факультативных паразитов, которые преимущественно ведут сапрофитный образ жизни и лишь иногда паразитируют, наоборот, преимущественно паразитируют и лишь иногда переходят к сапрофитному образу жизни. Круг их хозяев уже значительно уже, они плохо растут на питательных средах.
Облигатные паразиты (обязательные) в природе живут только за счет живых растительных тканей. Совсем недавно, говоря об облигатах, обязательно добавляли, что эти паразиты не способны расти на искусственных питательных средах. Теперь для некоторых из них удалось подобрать среды, на которых они с большим трудом, по все же развиваются. Поэтому точное было бы сказать, что облигатный паразит — это паразит, для которого питательная среда еще не подобрана. Они принадлежат к числу монофагов, т. е. их растением-хозяином является какой-нибудь один растительный род или близкие роды. Облигатами являются примерно 25 % всех паразитов растений. Они встречаются во всех главных группах растительных патогенов, за исключением бактерий. Что касается вирусов, то все они принадлежат к числу облигатов.
Многие облигатные паразиты чрезвычайно вредоносны (ржавчина и мучнистая роса). Если факультативные паразиты преимущественно поражают ослабленные растения, то облигаты, наоборот, часто выбирают своей жертвой сильные, активно функционирующие растения. Как правило, эти патогены обладают орудиями, позволяющими им проникать через неповрежденную поверхность растений. Паразиты этой группы в природе представлены многочисленными физиологическими расами.
Симбионты, сосуществование которых часто рассматривают как высшую стадию паразитизма, не страдают друг от друга, а даже извлекают пользу. Считается, что симбиоз подобен вооруженному миру, когда оба партнера мирно сосуществуют, однако только в том случае, если оба физиологически полноценны. Достаточно одному ослабнуть, как симбиотические отношения могут перейти в паразитические. Примером симбиозов является всем известная микориза на корнях многих растений.
Выделение этих групп паразитов весьма условно, и между ними существуют промежуточные формы. Тем не менее не трудно заметить, что группы паразитов в порядке, в котором они здесь описаны — от сапрофитов к облигатам, составляют ряд, в котором возрастает:
степень специализации (от полифагии к монофагии), доля паразитирования в жизненном цикле паразита, зависимость от своего растения-хозяина, требовательность к питательной среде, способность проникать в неповрежденную растительную ткань, способность поражать полноценные растения.
Итак, 6 раз «возрастают» различные способности фитопатогенов на пути к истинному паразитизму.
Вышеперечисленная систематика фитопатогенов зависит от того, какую долю их жизненного цикла занимает паразитирование на растении. Существует и другое разделение патогенов в зависимости от характера их питания. На этом основании все фитопатогены разделяют на некротрофы и биотрофы, причем граница между ними проходит где-то на уровне факультативных сапрофитов: все факультативные паразиты и часть факультативных сапрофитов имеют некротрофный способ питания, тогда как остальная часть факультативных сапрофитов и все облигаты питаются биотрофно.
Эволюция взаимоотношении системы паразит — растение

Некроз означает смерть. Эти паразиты перед тем, как проникнуть в клетку растения, ее убивают. Иногда их еще называют деструктивными паразитами, или паразитами теплого трупа. Гибель растительных клеток всегда предшествует внедрению таких паразитов. Зона отмерших клеток является как бы плацдармом для наступающих гиф патогена.
Эти паразиты имеют мощные системы токсинов и экстрацеллюлярных (внеклеточных) ферментов, которые паразит как бы высылает вперед. G помощью токсинов они убивают клетки, а с помощью систем ферментов превращают клеточное содержимое в удобную для усвоения форму. Поскольку токсины — низкомолекулярные вещества, они способны убивать клетки, находящиеся на значительном расстоянии от паразита. Например, Pliytophthora cinnamomi убивает клетки корня эвкалипта, находящиеся на расстоянии 0,5 мм от гиф гриба.
Теперь относительно биотрофов. Само название этих паразитов свидетельствует о том, что они могут получать продукты питания только из живых клеток хозяина. Биотрофы не губят клетки — во всяком случае на первых этанах паразитирования, а определенное время сосуществуют с ними. Орудия нападения у таких паразитов значительно менее агрессивны. Биотроф оказывает на клетку растения настолько мягкое воздействие, что пораженная клетка часто не ощущает его присутствия. Биотрофы дольше, чем некротрофы, используют растительную клетку как источник питания. Биотрофность требует, с одной стороны, отсутствия токсического действия на клетку, а с другой — способности избегать защитных реакций со стороны растения. Паразитарные грибы этого типа часто распространяются по межклетникам, тогда как в клетки проникают только их специальные присоски — гаустории, через которые выкачиваются из живой клетки необходимые паразиту питательные вещества. Наличие гаусториального аппарата вообще признак биотрофии. Целостность клеточной мембраны хозяина при этом не нарушается, гаустории как бы впячивают ее вовнутрь клетки. Гриб находится, как говорят, апопластно.
Разделение на биотрофы и некротрофы весьма условно. Во-первых, биотрофы лишь до определенного момента развиваются в живых клетках хозяина, не вызывая их быстрой гибели. Во-вторых, при многих болезнях происходит смена биотрофных и некротрофных процессов.
Поскольку биотрофы питаются готовыми органическими молекулами, которые синтезируются в клетке хозяина-растения, то паразит тесно приурочен к обмену питающего его растения и сильно от пего зависит. Изменение метаболизма хозяина может неблагоприятно сказываться на судьбе паразитирующего биотрофа.
Все паразиты — агрессоры, а агрессоры должны иметь орудие нападения. И такие орудия имеются у фитопатогенов, к числу которых в первую очередь принадлежат токсины и ферменты.
Токсины — одно из самых древних средств нападения микроорганизмов на высшие растения. Без них не могут обойтись факультативные паразиты, да и часть факультативных сапрофитов тоже. Вопрос о том, имеются ли токсины у биотрофов, до сих пор еще не решен.
Некротрофы, обладающие токсинами, также различаются по уровню своей специализации, в зависимости от того, как много растительных видов они поражают. В соответствии с этим их токсины отличаются по способности повреждать много или мало растений. По этому признаку все имеющиеся токсины можно разделить на три группы.
Фитотоксины — вещества, выделяемые микроорганизмами в среду, где они растут, убивающие ткани растений и тем самым превращающие их в пригодный для заселения питательный субстрат. Они не специфичны в действии, т. е. вызывают повреждения широкого круга растений как хозяев, так и нехозяев того фитопатогена, который их выделяет.
Вивотоксины выделяются не только в среду при сапрофитном питании паразита, но и в ткани зараженного растения на паразитических стадиях существования микроорганизмов. Токсины этого типа могут вызвать у растений некоторые симптомы, близкие к симптомам болезни. Однако с помощью вивотоксинов невозможно объяснить, почему данный паразит поражает именно эти растения, но не поражает другие.
Это удается объяснить с помощью токсинов третьей группы, называемых патотоксинами, специфичными к своему растению-хозяину (host specific toxins).
Патотоксин повреждают в пределах вида лишь те растения, которые восприимчивы к патогену, вырабатывающему эти токсины. Вероятно, патотоксины являются непосредственной причиной болезни, поскольку вызывают те же симптомы, что и само заражение. Эти токсины настолько точно отражают картину болезни, что ими можно в ряде случаев заменить самого паразита.
По природе патотоксины разнообразны — среди них встречаются олигопептиды, терпеноиды и гликозиды. Патотоксины продуцируют по крайней мере 9 видов паразитарных грибов-некротрофов, причем большая часть их выделена из грибов рода Hclminthosporium. Вот один из примеров их специфичности: фильтраты культуральной жидкости Н. victori, поражающего овес, подавляли рост его восприимчивых корней даже в концентрации 1:1200 000, тогда как корни устойчивого сорта лишь частично повреждались при концентрациях в 400 000 раз больших. Были исследованы 100 различных изолятов этого паразита. Из них 80, т. е. те, которые образовывали токсины, были вирулентными, а оставшиеся 20 токсинов не образовывали и вирулентностью не обладали. Если изолированный токсин добавляли к авирулентному изоляту, он приобретал свойство вирулентности.
Другим средством агрессии фитопатогенов является система их экзоферментов. Как правило, это деполимеразы, т. е. ферменты, расщепляющие клеточные полимеры до мономеров. Этим паразит преследует как бы две цели: разрушает покровные ткани и клеточные стенки хозяина и обеспечивает себя необходимым питанием.
Из числа ферментов, разрушающих клеточные стенки, прежде всего следует назвать пектолитические ферменты, т. е. ферменты, расщепляющие пектиновые вещества. Известно, что первичная клеточная стенка растений состоит из микрофибрилл целлюлозы, погруженных в сплошной матрикс из гемицеллюлоз и пектинов. Между клетками находится срединная пластинка, которая цементирует клетки друг с другом. Главной составной частью ее также являются пектиновые вещества.
Основными компонентами пектиновых веществ являются полимеры, состоящие из остатков галактуроновых кислот, в которые периодически вкраплены остатки рамнозы. Последние соединены с гемицеллюлозой, которая, в свою очередь, связана с фибриллами целлюлозы. Карбоксильные группы полигалактуронидов метилированы.
К пектолитическим ферментам принадлежат пектип-метилэстеразы, отрывающие метильную группу (группу — СН3) от полигалактуронида, а также ферменты, расщепляющие на отдельные фрагменты или деполимеризующие пектиновые вещества (гидролазы и лиазы). Пектолитические ферменты первыми воздействуют на клеточную стенку растений.
Строение клеточной стенки таково, что только после разложения пектиновых веществ могут освободиться и стать доступными для ферментов и другие полимеры (например, целлюлоза и гемицеллюлоза), подлежащие, в свою очередь, ферментативному расщеплению. Действительно, на примере многих фитопатогенных микроорганизмов показано, что пектолитические ферменты продуцируются ими в первую очередь, тогда как гемицеллюлазы и целлюлазы появляются позднее. Такой порядок продуцирования ферментов фитопатогенов является лишним доказательством ключевой роли пектолитических ферментов в процессе патогенеза.
Трудно назвать фитопатогена, у которого бы не удалось обнаружить пектолитические ферменты, их содержат возбудители гнилей, пятнистостей, ожогов и, наконец, инфекционного увядания. За последнее время установлена способность некоторых облигатных паразитов продуцировать пектолитические ферменты. Кроме пектолитических ферментов, патогены обладают также ферментами, расщепляющими другие полимеры клеточной стенки — целлюлазами, разрушающими целлюлозу, глюканазами, расщепляющими глюканы, гемнцеллюлазами — гемицеллюлозы. Патогены выделяют также ферменты, расщепляющие белки и полипептиды — протеазы.
СОПРЯЖЕННАЯ ЭВОЛЮЦИЯ РАСТЕНИЯ И ПАТОГЕНА,
ИЛИ КАК САПРОФИТ СТАЛ ПАРАЗИТОМ
Устойчивость в растительном мире — правило, а восприимчивость — исключение. Эти слова были произнесены видным фитопатологом К. Честером полстолетия назад. Иногда эту же мысль выражают несколько по-иному, говоря, что большинство растений устойчиво к большинству существующих фитопатогенов. К примеру, яблоня поражается примерно 50 паразитами из числа фитопатогенных грибов, а картофель — болеет 300 болезнями, возбудителями которых являются грибные, бактериальные и вирусные паразиты. Иными словами, если принять, что в природе существует около 10 000 паразитарных грибов, то из их числа яблоню поражают только 50, тогда как по отношению к остальным 9950 она является иммунной. То же самое и с картофелем: из общего числа 10 800 фитопатогенов (из них 10 000 грибных, 600 вирусных и 200 бактериальных) он является иммунным к 10500. Или только 300 патогенов приспособились паразитировать на картофеле и стали тем самым для пего специфическими, а картофель стал их растением-хозяином. Подавляющее число остальных фитопатогенов поражают другие растения, но не картофель, и поэтому не являются специфическими по отношению к нему.
Иммунитет, предохраняющий большинство растений от поражения большинством паразитов, не являющихся к этим растениям специфическими, называется неспецифическим иммунитетом (иммунный обозначает свободный — в данном случае от инфекции). Иногда такую форму иммунитета называют еще общим, конституционным, врожденным, или естественным. И. И. Вавилов называл его видовым иммунитетом, поскольку он предохраняет растения от поражения теми микроорганизмами, которые не приобрели способность поражать данный растительный вид. Естественный иммунитет защищает растения и от огромного количества окружающих их разнообразных видов сапрофитных микроорганизмов, питающихся мертвыми растительными остатками. Иногда его еще называют «нехозяйским» иммунитетом, тем самым подчеркивая, что он защищает от тех микроорганизмов, для которых данный растительный вид не является хозяином.
Как это ни парадоксально, но именно об этой основной форме иммунитета мы знаем особенно мало. Исследователей значительно в большей мере интересует не полная невосприимчивость, или иммунитет, а резистентность, или устойчивость, растений, их отдельных сортов к специфическим болезням, получившая название специфической или сортовой устойчивости. Эта форма устойчивости имеет наибольшую практическую значимость, поскольку специфические патогены обусловливают более 90 % всех потерь, которые песет растениеводство.
Для того чтобы стать для растения специфическим паразитом, последний должен претерпеть вместе с растением процесс сопряженной эволюции, иными словами, «повариться вместе с растением в едином эволюционном котле». «Эволюция паразитизма в растительном мире — любопытная, хотя и не слишком благодарная тема», — писал английский фитопатолог С. Тарр в своей книге «Основы патологии растений»
[12], поскольку в этой проблеме все основано на умозаключениях.
Большинство исследователей думают, что эволюция фитопатогенных грибов шла в направлении от сапрофитизма к облигатному паразитизму. Предположим, что почвенные сапрофиты, стремясь избежать конкуренции с себе подобными, пытались захватить иную «экологическую пишу» — растительные ткани. Рядом с такими сапрофитами могли оказаться пораненные, ослабленные корни растений, либо токсические выделения сапрофитов способны были вызвать отмирание корневой системы растений, ткани которых затем могли служить им источником питания. Переход из почвы в ткани растений вначале мог происходить не полностью: на первых порах паразитарное питание имело место только на отдельных этапах цикла развития паразита, тогда как на других микроорганизм оставался сапрофитом.
Какие свойства должен приобрести сапрофит для того, чтобы превратиться в паразита? Для этого он должен прежде всего преодолеть барьеры неспецифического иммунитета своего будущего растения-хозяина. Только после этого микроб приобретает способность превращать данное растение в источник своего питания, т. е. становится его патогеном. Таким образом, ранее независимые друг от друга сапрофит и растение превращаются в систему паразит — растение-хозяин. Барьеры видового неспецифического иммунитета на рис. 2 изображены в виде кирпичной стены. Сюда относятся покровы растений, антибиотические соединения, содержащиеся в растительных тканях, и др. Соответственно, чтобы приобрести способность поражать данный растительный вид, микроорганизм должен научиться прободать растительные покровы, суметь приспособиться к химическому составу растения, чтобы использовать его в качестве источника питания, разлагать его антибиотические вещества, а также приобрести и другие свойства, необходимые для паразитирования на этом виде растения.
 Рис. 2. Предполагаемая схема сопряженной онолюции паразита и растения-хозяина
Рис. 2. Предполагаемая схема сопряженной онолюции паразита и растения-хозяина
Итак, достигнут первый этап — специализация к виду растений. На втором этапе специализации (уже сортовой, а не видовой) среди особей, составляющих популяцию растительного вида, к которому приспособился паразит, возникают формы, обладающие генами специфической (сортовой) устойчивости. В ответ на это в популяции паразита возникают формы, несущие гены вирулентности, способные преодолевать гены устойчивости растений. И формы растений с генами устойчивости, и формы паразита с генами вирулентности возникают в ходе сопряженной эволюции паразита и хозяина. Теорию сопряженной эволюции растения и его паразита на их совместной родине выдвинул Н. И. Вавилов и впоследствии развил П. М. Жуковский.
Согласно теории сопряженной эволюции наибольшее разнообразие устойчивых форм растений, которые могут служить исходным материалом для селекции, следует искать именно на совместной родине растения и паразита там, где протекала их совместная «притирка» или сопряженная эволюция. И хозяин и паразит там проявляют исключительную изменчивость, в результате чего растение образует все новые и новые устойчивые разновидности и формы, а паразит формирует все новые и новые расы и биотипы.
Научное предвидение Н. И. Вавилова оказалось пророческим и полностью себя оправдало. Поиски доноров генов устойчивости, которые ранее велись методом проб и ошибок, приобрели целенаправленный характер. Ученые всего мира благодарны за это Н. И. Вавилову, который поставил на научную основу работу селекционеров.
Хорошим примером сопряженной эволюции служит картофель и возбудитель его наиболее опасной болезни фитофтороза. Родиной этих партнеров, где протекала их совместная эволюция, являются Мексика и Гватемала. Здесь находится древний центр формирования как диких клубненосных видов картофеля, так и основной центр формирования различных рас фитофторы. Растущие здесь растения картофеля имеют самый широкий в мире набор генов устойчивости, а паразит — наибольшее разнообразие генов вирулентности.
Наверное, давно бы следовало сказать, что любая форма иммунитета растений, как, впрочем, и других организмов, от полного иммунитета до той или иной степени устойчивости и восприимчивости, находится под строгим генетическим контролем, или, как говорят ученые, «генетически детерминирована». Попробуем в самой общей форме разобраться в том, как возникают гены специфической устойчивости у растений и гены вирулентности у паразитов в процессе их сопряженной эволюции.
Итак, барьер неспецифического иммунитета преодолен, паразит специализировался к виду растения, который страдает от новой болезни. Однако виды организмов являются популяциями, состоящими из особей, которые неоднородны, несмотря на их кажущееся морфологическое сходство. Вот эта неоднородность или гетерогенность популяций, образовавшаяся в результате их изменчивости, и лежит в основе возникновения специфического иммунитета.
Предположим, что в популяции растения, повсеместно восприимчивого к паразиту, возникает мутация, превращающая данную особь в устойчивую. Иными словами, некая особь растения или некие особи приобретают ген устойчивости к патогену. В результате присутствия такого гена данное растение перестает поражаться болезнью и в силу этого становится более жизнеспособным и более продуктивным. Естественно, что растения с таким свойством постепенно накапливаются в популяции, поскольку получают преимущество перед другими, которые продолжают болеть. Число растений с геном устойчивости (назовем его ген R
1 — от английского resistance — устойчивость) возрастает. Новый барьер устойчивости, обусловленный возникновением одного признака, управляемого одним геном, называется уже не видовым иммунитетом, а специфической (сортовой) устойчивостью.
Ген устойчивости является полезным геном, и поэтому теоретически в конечном счете многие особи, составляющие популяцию растения, могут приобрести этот ген, что, конечно, не может не создать угрозы существованию паразита. Ведь последний может потерять свои позиции. Однако ничуть не бывало. В ответ на появление устойчивых форм растений у паразита также начинают отбираться и накапливаться мутантные формы, способные поражать формы устойчивых растений. Такие новые формы (расы) паразита обладают геном вирулентности, способным преодолевать ген устойчивости R
1. Все особи, обладающие этим геном, представляют собой вновь возникшую расу (назовем ее расой 1), к которой ранее устойчивые формы с геном R
1 становятся восприимчивыми.
Тогда в растительной популяции начинают отбираться мутантные формы, обладающие геном устойчивости R
2. В ответ на это у паразита возникает и накапливается раса 2, несущая в себе ген вирулентности, способный преодолевать ген устойчивости R
2. По такому же принципу в популяции растений возникают формы с генами устойчивости R
3 и R
4 и так далее и соответственно расы паразитов 3 и 4. Инициатива в возникновении новых генов в этом эволюционном марафоне принадлежит растению, поскольку новые вирулентные формы паразита не начнут накапливаться до тех пор, пока не возникнут устойчивые формы растений, которые им предстоит осваивать. Естественный отбор будет благоприятствовать тем новым признакам растений, которые защищают их от паразита, и тем новым признакам паразитов, которые преодолевают это свойство хозяина.
В природе гены могут возникать не последовательно, а параллельно, в результате чего в больших популяциях способны одновременно накапливаться несколько генов. Чем длительнее сосуществование партнеров, тем большее количество таких форм накапливается. Естественно, что на совместной родине растения и его паразита период их сосуществования был наиболее долгим. Именно поэтому там в наибольшем количестве и накапливаются устойчивые формы растений и физиологические расы паразита.
Таким образом, в результате сопряженной эволюции у хозяина и его паразита возникают комплементарные (друг другу соответствующие) пары генов — ген устойчивости у растения и преодолевающий его ген вирулентности у паразита.
Вот это и есть одна из центральных догм фитоиммунологии — теория ген — на — ген. Она была создана американским фитопатологом Т. Флором в начале 50-х годов. Основанием для ее создания была длительная работа Флора с ржавчиной льна. В его распоряжении было около 200 рас этого паразита, а также большое число сортов льна с различным количеством генов устойчивости. Оказалось, что сорта льна, имеющие один ген устойчивости, поражаются расой, имеющей один ген вирулентности. Для того чтобы поразить сорт с двумя генами устойчивости, раса должна как минимум обладать двумя генами вирулентности и т. д. Иными словами, сорт растения, обладающий определенным геном (или генами) устойчивости, будет поражаться расой только в том случае, если у нее есть соответствующий геи (или гены) вирулентности, способные преодолевать гены устойчивости растения. Если у расы такого гена нет — сорт окажется устойчивым. Возникновение комплементарных генетических систем является результатом сопряженной эволюции партнеров. Таким образом, теория Флора ген — на — ген является логическим завершением теории сопряженной эволюции.
Поскольку возникновение системы ген — на — геи возможно только в процессе длительной коэволюции хозяина и паразита, в результате чего и достигается их тесная эволюционная «притирка» или специализация, то, следовательно, такие системы могут существовать в основном у высокоспециализированных паразитов из групп облигатов или факультативных сапрофитов. У малоспециализированных паразитов такие отношения менее вероятны. В настоящее время существование системы ген — на — ген подтверждено для фитофтороза картофеля, ржавчины пшеницы и кофе, парши яблок, головни овса и других болезней, возбудителями которых являются специализированные патогены.
До сих пор еще никто не знает, что представляет собой ген устойчивости растения, равно как и геи вирулентности паразита. Ни один продукт этих генов еще не охарактеризован, не установлен путь от гена устойчивости или вирулентности к признаку устойчивости или вирулентности. Тем не менее за последнее время получен ряд фактов, которые приближают нас к пониманию природы этого явления. В главе «Загадка специфичности» мы попытаемся проиллюстрировать предложенную здесь схему сопряженной эволюции паразита и хозяина на молекулярном уровне в свете последних полученных данных.
Из того, что мы сказали, вытекает важный практический вывод: сорт растения, обладающий геном устойчивости, как правило, может быть в конечном счете побежден новой расой паразита, которую этот же сорт и вызвал к жизни. Возникновение новых вирулентных рас паразита — бич современного растениеводства. Именно их возникновение и распространение является одной из главных причин того, что жизнь вновь выведенных устойчивых сортов становится все короче и короче. Недаром J Г. М. Жуковский, говоря о быстром приспособлении паразита к новым сортам растения, пишет, что там, где подозревается адаптация паразита к хозяину, следует искать возникновение новой расы.
Не нужно думать, что новые расы паразита один раз где-то возникают, а затем распространяются по всей Земле. Расы возникают и исчезают повсеместно. Могут возникнуть там, где появляется сорт с новым геном, вызывающим их к жизни. Могут исчезнуть после того, как этот сорт исчезает из посевов. Расообразовательные процессы у патогенов протекают в природе с поразительной быстротой, поскольку скорость размножения паразита во много раз превышает скорость размножения растения. Так, на одном га посевов ежегодно формируется от 10
11 до 10
13 спор возбудителей фитофтороза, ржавчины и некоторых других болезней. А если принять во внимание, что мутации по каждому гену возникают в среднем одна на 10
7—10
9, то ежегодно на одном га посевов будут от 10
2 до 10
6 мутирующих спор патогенов, среди которых могут быть формы с тем или иным геном вирулентности, дающие начало новым расам.
Однако причинами возникновения рас, обладающих новыми генами устойчивости, могут быть не только мутации, но и гетерокариоз (разнокачественность ядер в грибной клетке), гибридизация и другие процессы.
Раса может обладать не одним, а двумя или более генами вирулентности. Такие расы называют сложными в отличие от простых, обладающих одним геном. Например, в случае возбудителя фитофтороза картофеля раса с геном вирулентности 1 будет простой расой 1, а раса с двумя генами вирулентности 1 и 2 называется сложной расой 1, 2. Раса с генами 1, 2, 3 и 4 — сложной расой 1, 2, 3, 4 и т. д.
Наличие в сорте картофеля определенного гена устойчивости, например гена R
1, делает этот сорт устойчивым ко всем расам, в названии которых цифра 1 отсутствует, и, восприимчивым к тем расам, в наименовании которых эта цифра имеется. И наоборот, раса фитофторы будет поражать сорта картофеля только в том случае, если у нее есть гены вирулентности, преодолевающие гены устойчивости хозяина.
По логике вещей, казалось бы, чем большим количеством генов вирулентности обладает раса, тем выше ее жизнеспособность. На самом деле это совсем необязательно. Если высевать на питательную среду простые и сложные расы вместе, то через некоторое время окажется, что сложные расы, не выдержав конкуренции с простыми, постепенно исчезают. То же самое будет, если смесью из простых и сложных рас заражать сорт картофеля, который не имеет генов устойчивости и потому поражается всеми расами возбудителя фитофтороза. Спустя некоторое время простые расы вытесняют сложные. Описанное явление есть результат действия стабилизирующего отбора, к которому нам еще придется вернуться в этой книге. Иными словами, в популяции паразита накапливаются лишь те гены вирулентности, которые необходимы для преодоления генов устойчивости растения-хозяина. Лишние, не использующиеся гены не только не полезны, по и вредны, и ими обладающие расы исчезают из популяции. Когда в популяции растений появляются новые сложные гены устойчивости, тогда и возникают сложные, их преодолевающие расы, которые существуют до той поры, пока существуют сорта, которые их вызывали к жизни. Простые расы такие сорта поразить не могут, и поэтому сложные расы на этих сортах не имеют конкурентов. Но зато последние по выдерживают конкуренции с простыми расами на сортах с одним геном устойчивости. Вот уже поистине за все в этом мире приходится платить, и за излишнюю вирулентность тоже.
Раса, которая поражает сорт растения, называется совместимой к нему; раса, неспособная поражать, т. е. та, к которой сорт устойчив, несовместимой. Таким образом, хорошее человеческое понятие — совместимость (которое обычно связано с положительными моментами — совместимостью людей, характеров) в фитопатологии понимается наоборот: совместимая к сорту раса вызывает его болезнь, тогда как по отношению к несовместимой он остается здоровым.
Наличие генов вирулентности у рас определяют по их способности заражать набор сортов растения-хозяина, обладающих определенными генами устойчивости. Набор таких сортов носит название сортов-дифференциаторов, поскольку по ним удается различить расы патогенов.
Число генов вирулентности в популяции паразита, как уже говорилось выше, будет равно числу генов устойчивости в популяции растения-хозяина. Если число генов вирулентности обозначить буквой
п, то количество теоретически существующих рас паразита будет составлять 2
n. Например, у возбудителя фитофтороза картофеля сейчас известно не менее 11 генов вирулентности (соответственно у картофеля имеется не менее 11 генов устойчивости). Это означает, что число рас паразита, которое теоретически может существовать в природе, будет составлять 2
11, или 2048 рас. У возбудителя стеблевой ржавчины пшеницы свыше 20 генов вирулентности, а значит, число рас будет баснословно велико. Это не означает, что все эти расы разом существуют в природе, но теоретически они могут возникать, если появятся посевы с соответствующими генами устойчивости, которые их и вызывают к жизни.
Известный фитопатолог Я. Ван-дер-Планк считает, что в природе могут существовать два типа рас. Расы, о которых только что шла речь, поражают одни сорта растений, но зато совсем не поражают другие. Иными словами, один и тот же сорт одними расами поражается, а другими — не поражается.
Фитопатологи в таком случае говорят, что расы дифференцированно (различно) реагируют с сортами растений. В этом случае взаимодействие сортов и рас идет по принципу да — нет. Есть поражение — да, нет поражения — нет. Мы уже знаем, что поражение есть тогда, когда ген вирулентности расы может преодолевать ген устойчивости растения. Такие расы различаются по вирулентности. Взаимоотношения именно таких рас и сортов описываются системой ген — на — ген.
Другие встречающиеся в природе расы не дают дифференцированной реакции с сортами растений. Предположим, выделен набор рас от А до Е, которые по их способности заражать определенный сорт растения можно выстроить в порядке А>Б>В>Г>Д>Е, где раса А поражает данный сорт в самой большой степени, а раса Е — в наименьшей. Такой порядок поражения сохранится на любом сорте данного растения: раса А будет самой вредоносной, а раса Е — наименее вредоносной. Разница между ними состоит в степени поражения, т. е. она не качественная (да — нет), а количественная (меньше-больше). Такие расы различаются не по вирулентности, как ранее описанные, а по степени агрессивности. Вирулентность и агрессивность — две стороны патогенности паразита. Вирулентность определяется отдельными генами, агрессивность контролируется многими генами или является полигонной системой. Если вирулентность контролирует качественное свойство (да или нет, поражение или непоражение), то агрессивность количественное — степень поражения, что определяется скоростью роста паразита, интенсивностью его спороношения и другими факторами. Вирулентность мало зависит от условий окружающей среды, она может изменяться только в результате изменения генома. На агрессивность условия среды оказывают большее влияние, и поэтому она может изменяться в довольно широких пределах, Иными словами, вирулентность — качественная сторона патогенности, тогда как агрессивность — количественная.
Раса паразита с геном вирулентности может обладать различной степенью агрессивности. К примеру, раса паразита обладает геном вирулентности, комплементарным к соответствующему гену устойчивости сорта растения, и поэтому способна его поражать. Но она может его поражать в большей или меньшей степени — вот это и есть проявление агрессивности. Если патоген с геном вирулентности долгое время растет на питательной среде, то его способность проникать в ткани растения-хозяина уменьшается, спороношение сокращается, агрессивность падает. Но достаточно сделать несколько пассажей этой расы через растение, т. е. культивировать ее не на питательной среде, а на растительных тканях, раса вновь приобретает агрессивность, при условии, конечно, что за время ее роста в среде не утерялся ген вирулентности.
Как правило, расы, различающиеся по генам вирулентности и дающие дифференцированную реакцию с сортами, свойственны высокоспециализированным паразитам: облигатам или факультативным сапрофитам, которые претерпели со своим растением длительный период совместной эволюции, в результате чего и выработались эти гены вирулентности. Расы, не имеющие генов вирулентности, различающиеся количественно по степени агрессивности, обычно присущи менее специализированным паразитам — полифагам, стоящим на низкой эволюционной ступени паразитизма. Вирулентность — это как бы дополнительная надстройка на агрессивность паразита, выработавшаяся в процессе его специализации к отдельным сортам или формам внутри вида хозяина.
Свойству вирулентности и агрессивности у паразитов соответствуют вертикальная и горизонтальная устойчивость у растений. Именно она и определяет устойчивость отдельных сортов к несовместимым для них расам патогенов, в том случае, если сорт обладает геном, некомплементарным гену вирулентности паразита. Такой вид устойчивости определяется наличием у сорта растения одного или нескольких генов устойчивости. На рис. 3 приведено графическое изображение такой фитофтороустойчивости для сорта картофеля, обладающего генами Р
1. Сорт с таким генотипом будет поражаться совместимыми к нему расами паразита 1; 1,2 и 1,3, но не будет поражаться несовместимыми расами
2 и 3. Устойчивость на этом графике выглядит в виде вертикальных столбцов, почему и получила название вертикальной.
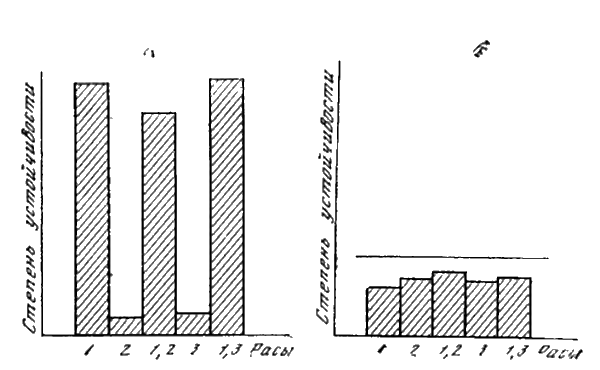 Рис. 3. Вертикальная (а) и горизонтальная (б) устойчивость
Рис. 3. Вертикальная (а) и горизонтальная (б) устойчивость
Второй тип устойчивости в отличие от вертикальной получил название горизонтальной. Иногда ее еще называют полевой или относительной. Механизм горизонтальной устойчивости еще недостаточно изучен. Опа контролируется многими генами и поэтому называется полиген-ной. Если вертикальная устойчивость обеспечивает полную устойчивость, но лишь к отдельным расам паразита, то горизонтальная обусловливает хотя и относительно низкий уровень устойчивости, но зато по отношению ко всем без исключения расам. Постоянство и эффективность против всех рас паразита — ее наиболее ценное свойство. Соответственно на правой части рисунка представлено поражение тем же набором рас фитофторы, но уже другого сорта картофеля, не имеющего R-генов, но обладающего горизонтальной устойчивостью. Из рисунка видно, что полевая устойчивость примерно в равной, хотя и в небольшой мере предохраняет сорт от поражения всеми расами и поэтому изображена на графике в виде горизонтальной полосы, отчего и получила название горизонтальной.
Итак, если сорт к одним расам патогена проявляет устойчивость, а к другим нет, то его устойчивость называется вертикальной. Если же небольшой уровень устойчивости расы равномерно распределяется по отношению ко всем расам патогена — она является горизонтальной. Некоторые сорта растений могут сочетать в себе и горизонтальную и вертикальную устойчивость. Первая — количественный признак. Сорту может быть свойствен низкий, средний или высокий уровень горизонтальной устойчивости. Вторая — признак качественный и работает по принципу «да или нет».
В большинстве случаев от полифагов растение защищает горизонтальная устойчивость. От высокоспециализированных монофагов растения бывают защищены вертикальной, а иногда и сочетанием вертикальной и горизонтальной устойчивости. Вертикальная устойчивость подразумевает существование дифференцированной реакции между сортами хозяина и расами паразита, а при горизонтальной такая реакция отсутствует.
Нам еще не раз придется возвращаться к этим вопросам в следующих главах этой книги.
СТРАТЕГИЯ САМООБОРОНЫ
Иммунитет приходится рассматривать как результат взаимодействия многих слагаемых.
Н. И. Вавилов
[13]
СУММА СЛАГАЕМЫХ
Н. И. Вавилову принадлежит высказывание о том, что фитоиммунитет представляет собой сумму из множества слагаемых. «Новейшие данные показали еще более ясно сложность природы иммунитета растений и невозможность свести ее к единой физиологической и биологической причине»
[14]. Слагаемыми являются отдельные защитные реакции, которые в сумме и определяют устойчивость растения. Речь, конечно, идет не о механической сумме отдельных реакций, а об их взаимодействии друг с другом. Одни защитные реакции дополняют друг друга, в результате чего вместе взятые они и оказывают больший эффект, чем каждая в отдельности. В ряде случаев они наносят паразиту последовательную цепь разных по природе ударов, под воздействием которых последний в конечном счете погибает.
В этом отношении растения мало чем отличаются от животных организмов, устойчивость к болезням у которых также основывается на множество разнообразных защитных механизмов, как внутренних, так и внешних, многие из которых обладают неспецифическим действием.
Многочисленные защитные механизмы растений против возбудителей инфекционных болезней обычно разделяют на конституционные, т. е. присутствующие в интактных тканях хозяина, и индуцированные, т. е. возникающие в них в ответ на контакт с паразитом или его экстрацеллюлярными соединениями. Вроде бы нетрудно доказать, что в большинстве случаев первостепенная роль принадлежит индуцированным защитным механизмам. Ведь один и тот же сорт растения с одними и теми же конституционными механизмами является устойчивым к одной расе патогена и одновременно восприимчивым к другой. Следовательно, устойчивость, равно как и восприимчивость, развивается при взаимодействии неких комплементарных механизмов обоих партнеров. Но ведь сама комплементарность теснейшим образом связана с какими-то конституционными элементами хозяина и паразита, которые присутствуют в них еще до контакта друг с другом и которым, по-видимому, принадлежит первостепенная роль во взаимодействии партнеров.
К числу защитных реакций растений принадлежат различного рода механические барьеры, как предсуществующие в растении, так и образующиеся в нем в ответ на инфицирование, состоящие из клеток, стенки которых пропитаны стойким веществом типа суберина, кутина и лигнина. Сюда же относятся механизмы, с помощью которых паразит обездвиживается. Важнейшим средством защиты является наличие в растительных тканях антибиотических соединений, способных подавить рост паразита и инактивировать его токсины и экзоферменты. К числу защитных реакций растений относится и их способность создавать в своих тканях недостаток веществ, необходимых паразиту для роста и развития, и другие способы уничтожить паразита или ограничить его рост.
Предметом фитоиммунологии является множество болезней, вызываемых множеством патогенов у множества растений. Если в центре внимания медицины стоит один-единственный вид — Homo sapiens — человек разумный, то фитоиммунология занимается сотнями видов растений, из которых каждый подвержен многим болезням. Естественно, что у разных партнеров могут быть разные защитные механизмы, по крайней мере одним и тем же защитным реакциям может принадлежать разный удельный вес в сумме защитных механизмов разных болезней. Последнее только усиливает необходимость поиска среди них тех общих механизмов устойчивости, которые характерны для растений в целом.
Одной из таких реакций, вероятно, свойственной всему живому, является необходимость обеспечить энергией средства защиты. Ведь для того чтобы бороться, нужны сырье, машины, энергия. Поэтому универсальной ответной реакцией устойчивых растений на большинство неблагоприятных воздействий, в том числе и на инфицирование фитопатогенными микроорганизмами, служит усиление процессов дыхания и энергетического обмена. Большой вклад в исследование роли окисления в больном растении внес советский исследователь профессор Б. А. Рубин, который является основоположником школы о защитных функциях окислительных и энергетических процессов в явлении фитоиммунитета.
Процессы окисления и связанные, или, как говорят, сопряженные, с ними процессы фосфорилирования снабжают клетку энергией. Последняя запасается в виде богатых энергией (макроэргических) фосфорных связей в универсальном биологическом горючем — аденозинтрифосфорной кислоте (АТФ). Молекулы АТФ направляются в те части клеток, где в них возникает необходимость. В устойчивом растении, находящемся в условиях стресса, производство таких молекул возрастает, поскольку построение механических заграждений, обездвиживание паразита и особенно синтез антибиотических веществ требуют затрат энергии.
Дыхание служит не только источником энергии, но и поставщиком исходного пластического материала для различного рода биосинтетических процессов. Поэтому усиление активности дыхания одновременно с энергией поставляет и сырье для синтеза соединений, необходимых для обороны.
Каким образом удается клеткам усиливать процессы окисления и фосфорилирования? Главным образом за счет возрастания числа митохондрий. Митохондрии, как известно, являются энергетическими центрами клетки, в которых и вырабатывается АТФ. Естественно, что две митохондрии вместо одной образуют и вдвое большее количество энергии. Для добывания необходимой энергии и сырья защищающееся растение пускает в ход все свои запасы и резервы. Запасные полимеры клетки в виде различного рода полисахаридов распадаются на составные части, которые сжигаются в акте дыхания. Только такой ценой достигается противостояние паразиту.
Естественно, что генерировать энергию и сырье могут только живые клетки растений. Клетки убитые либо поврежденные токсином, так же как колонизированные паразитом, уже выведены последним из строя и ни на что не пригодны. Поэтому устойчивость прежде всего и состоит в способности растительных тканей не допустить воздействия паразита на свои клетки, с тем чтобы дать возможность соседним здоровым клеткам мобилизовать свои ресурсы на борьбу с инфекцией.
И. И. Мечников писал, что у организмов существуют два типа реакций, защищающих от инфекций. Один из них заключается в обезвреживании токсических выделений паразита, тогда как другой (основной) состоит в подавлении самого паразита. Такое разделение поразительно соответствует различной стратегии защитных реакций растений, направленных против некротрофов и биотрофов.
Выше уже говорилось о том, что некротрофные паразиты еще до проникновения в растительную клетку убивают ее токсинами, а затем питаются содержимым убитых клеток, переваривая их с помощью своих ферментов. Поэтому если стратегия некротрофов состоит в совершенствовании своих орудий убийства, то стратегия защиты от них заключается в обезвреживании этих орудий.
Иное дело биотрофы. Если условием паразитирования некротрофов является убийство растительных клеток, то биотроф, напротив, не может погубить питающую его клетку, поскольку ее гибель ставит под угрозу существование паразита. Растение безошибочно находит уязвимое место биотрофа. Если для его существования необходима живая клетка, то следует самому погибнуть, чтобы не допустить развитие паразита. А уже потом за паразита, находящегося в погибшей клетке, принимаются антибиотики, которые и доводят его до гибели. Естественно, что так гибнут не все растительные клетки, а только те из них, в которые паразит проник. Гибель части во имя спасения целого и лежит в основе стратегии защиты растительной клетки от биотрофно питающихся паразитов. Она получила название сверхчувствительности и является своеобразной защитной реакцией, характерной для растений. Реакцию сверхчувствительности мы будем еще подробно рассматривать в дальнейшем. Ниже коротко суммированы общие принципы агрессии некротрофов и биотрофов, так же как стратегия защиты от них.

РАНЕВОЙ СТРЕСС
Трудно, пожалуй, найти другой научный термин, который так прочно вошел бы в жизнь современного общества, как стресс. Впервые этот термин ввел известный канадский физиолог Ганс Селье в 1936 г. для объяснения реакции организма на самые различные невзгоды, для преодоления которых требуется напряжение всех его сил.
Согласно определению самого Селье, стресс представляет собой совокупность всех неспецифических изменений, возникающих под влиянием любых сильных воздействий, и сопровождается перестройкой защитных сил организма. Уже из определения следует, что неспецифические реакции имеют прямое отношение к иммунитету против инфекционных болезней, хотя в целом иммунитет рассматривается как проблема специфичности. Ведь что может быть более специфичным, чем реакция антиген-антитело. И все же очень большую роль, особенно в иммунитете растений, играют неспецифические реакции. Более того, они в ряде случаев могут определить исход взаимоотношений между хозяином и паразитом. К тому же изучение неспецифических реакций может приблизить нас к пониманию специфических и, следовательно, к проблеме иммунитета в целом.
К числу самых распространенных неспецифических защитных реакций, по крайней мере в растительном мире, относится реакция на механические поранения, которые неизбежны при ветре, граде и других неблагоприятных условиях. Такого рода реакции, несомненно, возникли еще до появления паразитов, и можно думать, что они в какой-то мере явились основой для развития у растений защитных реакций против паразитов. Ведь многие паразиты способны проникать в растительную ткань только лишь после того, как механически ее повредят.
Реакцию растений на механическое повреждение часто называют еще процессами репарации, залечивания, заживления, образования раневой перидермы. Особенно хорошо исследована раневая реакция клубней картофеля, которые являются излюбленным объектом для проведения такого рода работ. Неповрежденный клубень покрыт так называемой естественной перидермой, точной копией которой является раневая перидерма, которая образуется на пораненном участке. Естественная перидерма защищает клубни от неблагоприятных воздействий и излишнего испарения.
Под микроскопом видно (рис. 4 см. вклейку), что естественная перидерма представляет собой многослойные ряды уплощенных клеток, расположенных одна под другой. Оболочки таких клеток содержат особое вещество типа пробки, носящее название суберина. Суберин адкрустирует клеточную оболочку, накладываясь на нее изнутри. Под электронным микроскопом суберинизация клеточных стенок выглядит слоистой из-за чередования слоев суберина и воска.
Суберин представляет собой полимер оксимонооксидин дикарбоновых кислот. В составе его находятся также некоторые полициклические вещества терпеновой природы. Вещества, входящие в состав пробки, делают ее высокоустойчивой к влиянию множества химических воздействий, таких, как органические кислоты, разбавленная соляная кислота, слабые щелочи. Наличие суберина в составе покровных тканей усиливает их механические свойства, поскольку большинство микроорганизмов не содержит ферментов, расщепляющих эти полимеры.
В процессе суберинизации происходит закупоривание пор в клеточных оболочках, в результате чего прерываются межклеточные контакты. Содержимое клеток, оболочки которых пропитываются суберином, постепенно отмирает, их полости наполняются воздухом, в результате чего клетки становятся термо- и влагонепроницаемыми.
В ответ на механическое поранение растения отвечают защитной раневой реакцией, суть которой сводится к образованию на пораненном участке точно такой же перидермы, какой является естественная. В результате поранения нарушается структурная и функциональная целостность растения или отделенного от него органа. В ответ на нарушение включаются системы иммунологического контроля, имеющие целью ликвидировать повреждение и восстановить нарушенную целостность. Образующаяся на месте поранения раневая перидерма по своим структурным и функциональным характеристикам ничем не отличается от естественной. Ее образование имеет целью как можно скорее закрыть ворота в кладовые растительных тканей, которые оказались открытыми в результате поранения. Неспособность растительных тканей быстро реагировать на повреждение может оказаться гибельной для организма в целом.
Как же происходит образование раневой перидермы? Взрослые клетки, давно прекратившие клеточные деления и находящиеся на поверхности поранения или залегающие непосредственно под ним, вновь приобретают меристематическую активность (способность к делению). Клетки обычно делятся параллельно пораненной поверхности, в них закладывается феллоген или пробковый камбий (см. рис. 4). Феллоген начинает делиться, отделяя наружу себе подобные по форме клетки, которые называются феллемой. В большинстве случаев это мертвые клетки, оболочки которых пропитаны суберином. Внутрь клубня феллоген также отпочковывает часть клеток такой же формы, которые остаются живыми и носят название феллодермы. Весь перидермальный комплекс состоит из приблизительно одинаковых расположенных друг под другом клеток феллемы, феллогена и феллодермы.
Для того чтобы взрослые клетки на поверхности поранения приобрели меристематическую активность, они должны получить стимул к делению или раневому возбуждению. Вокруг природы этого стимула развернулась длительная дискуссия. Существование раневого гормона предсказал еще 60 лет тому назад Г. Габерлапд на основании своих опытов. На разрезанной поверхности кольраби, как и на клубне картофеля, образовывалась раневая перидерма. В том случае, если поверхность ломтика промывалась водой, образование перидермы задерживалось. Если же на предварительно промытую поверхность наносили кашицу из растертых тканей, перидерма вновь начинала образовываться. Это позволило Габерланду прийти к заключению, что из продуктов отмерших и разрушенных клеток образуются вещества, вызывающие раневое возбуждение. Он назвал их раневыми гормонами или пекрогормонамп.
Вещество, получившее название раневого гормона, было выделено в конце 30-х годов Д. Инглишем и Д. Боннером. Им оказалась Д-децеи-1,10-дикарбоновая кислота, получившая название травматиновой кислоты, или травматина. Это вещество было обнаружено в картофеле, брюссельской капусте, люцерне, рисе, томатах, апельсинах и других растениях, но отсутствовало в тканях животных. В качестве раневого гормона испытывались также иные кислоты жирного ряда. Выяснилось, что активность коррелировала с присутствием двойной связи в молекуле и возрастала при увеличении числа углеродных атомов.
В самое последнее время другое вещество, напоминающее по своим свойствам раневой гормон, было обнаружено у ряда растений. Под влиянием различных стрессовых воздействий, в том числе и поранения, это вещество освобождалось из клеточных стенок растений, быстро передавалось по проводящей системе и индуцировало в здоровых тканях, удаленных от места повреждения, образование ингибиторов протеиназ, т. е. веществ, подавляющих ферментные системы, расщепляющие белки. Растения, таким образом, как бы заранее готовятся отражать атаку ферментов паразитарного микроорганизма, который может проникнуть через места механических поранений. Поскольку образующийся сигнал принадлежит самому растению, а не вводится паразитом, он подходит под определение раневой гормон. По предварительным данным, вещество представляет собой рамногалактуронан.
Вещество широко распространено в растительном мире. Если экстракты из 37 видов растений наносили на молодые листья томатов, то они индуцировали в них накопление ингибиторов протеиназ. Только два вида экстрактов из сельдерея и кабачка не вызвали их накопления.
Еще 40 лет назад А. Л. Курсанов установил, что процесс раневого залечивания сопровождается возрастанием дыхания. Субстратом возросшего дыхания в залечивающихся клубнях картофеля являлся крахмал, количество которого в тканях, прилегающих к месту поранения, постепенно уменьшалось, взамен чего возрастали моносахара, сгорающие в акте дыхания.
Процессы репарации нуждаются в источниках энергии. Показано, что в зоне поранения возрастало число митохондрий. Дополнительно образованные митохондрии генерировали энергию с такой же скоростью, как и пред-существующие в свеженарезанных клубнях. Очевидно, энергия, необходимая растениям для залечивания поранений, реализуется за счет дополнительно образовавшихся митохондрий.
В ответ на механическое поранение в клубнях картофеля происходило также образование нуклеиновых кислот. Показано, например, что через 10 минут после разрезания в бобе гороха в 10 раз возрастало количество рибонуклеиновой кислоты. В зоне поранения увеличивалось также и содержание аскорбиновой кислоты. К примеру, в раневой зоне клубня картофеля содержание аскорбиновой кислоты в 4–5 раз выше, чем ее количество в интактных клубнях.
Неоднократно предпринимались попытки установить корреляцию между устойчивостью клубней картофеля и толщиной их покровов. В конце концов исследователи вынуждены были признать, что такой зависимости не существует. Не обнаружено связи и между способностью клубней картофеля образовывать раневую перидерму и ее устойчивостью к ряду паразитарных микроорганизмов. По-видимому, образование перидермы на поверхности поранения клубней картофеля является второстепенным фактором в устойчивости клубней к ряду фнтопатогенов, поскольку для ее образования требуется время, в течение которого паразит может проникнуть в клубень. Однако уже сформированная раневая перидерма вполне способна надежно защитить клубни от проникновения инфекции.
Еще сравнительно недавно считалось, что защитная роль перидермы по отношению к фитопатогенным микроорганизмам определяется только ее механическими свойствами. Позднее стало ясным, что перидерма является не только механическим, по и токсикологическим барьером на пути проникновения инфекции. Дело в том, что в зоне поранения накапливаются антибиотические вещества, которые подавляют проникновение инфекции. Эти вещества различной химической природы и, следовательно, обладают различным механизмом действия. Такие антибиотические вещества присутствуют как в составе раневой, так и естественной перидермы. Они как бы защитным чехлом покрывают растение и его органы снаружи. Вот, например, как располагаются ингибиторы в составе перидермы клубня картофеля: в наружных отмерших клетках феллемы находятся некоторые фенольные соединения, в том числе кофейная кислота. Под ними в меристематической ткани залегают высокотоксические гликоалкалоиды, а еще ниже располагается хлорогеновая кислота. Таким образом, проникающему паразиту, помимо механических покровов, приходится преодолевать несколько последовательно расположенных токсических барьеров из ингибиторов различной химической природы с различными спектрами антибиотического действия. К тому же расположение ингибитора в отмерших тканях наружных растительных покровов позволяет создать их чрезвычайно высокие локальные концентрации
РАЗОРУЖЕНИЕ ПАРАЗИТА
Разоружать можно только вооруженного врага. А таким до зубов вооруженным паразитом является некротроф, обладающий мощными орудиями агрессии в лице токсинов и экзоферментов. Однако и биотрофы не лишены оружия нападения, хотя оно у них утонченно и усовершенствовано. Ведь этим паразитам все время приходится лавировать между опасностью погубить питающую их клетку хозяина и необходимостью прокормить самого себя. Итак, токсины…
Еще в начале нашего века предполагалось, что оксидазы растений, в частности пероксидаза, могут участвовать в разрушении токсинов микроорганизмов. Однако экспериментальные данные о способности растительных тканей детоксицировать токсины патогенов пока еще очень ограниченны. Известно, например, что устойчивые растения овса, райграса и сорго способны поглощать и детоксицировать виктории — токсин возбудителя гельминтоспориоза овса. Из тканей устойчивых растений токсин быстро исчезал, тогда как восприимчивые растения теряли тургор и выделяли викторин в окружающую среду. Механизм обезвреживания токсина пока не ясен.
Известен и другой способ растений избежать воздействия токсинов паразита — потерять к ним чувствительность, но это возможно только лишь по отношению к специализированным патотоксинам, имеющим в клетке определенную мишень действия. Что касается фито- и вивотоксинов, не обладающих избирательностью действия и одновременно нарушающих многие звенья обмена, то к ним вряд ли могут возникнуть нечувствительные формы. Вообще к ингибиторам, не имеющим определенной мишени, на которую они действуют, а неспецифически повреждающим многие «сайты», адаптация маловероятна. Возможно, поэтому к неспециализированным некротрофным паразитам, обладающим неспецифическими токсинами с широким спектром антибиотического действия, нет и не может быть устойчивых форм растений, а существуют только более или менее восприимчивые.
Примером устойчивости растений, основанной на его нечувствительности к токсину паразита, являются взаимоотношения сахарного тростника и возбудителя глазчатой пятнистости Helminthosporium sacchari. Гриб вызывает на листьях тростника своеобразные пятна в виде глаз, от которых распространяются красно-коричневые полосы. Поскольку возбудитель удается выделить только из пятен, считается, что сами гифы гриба находятся в местах пятен, а полосы — это результат воздействия его токсина. Токсин получил название гельминтоспорозида и представляет собой 2-оксициклопропил-
D-галактопиранозид.
Было испытано 182 клона сахарного тростника и установлено, что устойчивые к паразиту клоны устойчивы и к его токсину. В чем дело? Может быть, в устойчивых растениях токсин разлагается, поскольку в них есть ферменты, способные его детоксицировать? Определения показали, что это не так. Меченый токсин вводили в растения как устойчивых, так и восприимчивых клопов тростника, а затем спустя некоторое время определяли его количество, оставшееся в растениях. Разницы не нашли.
Картина несколько прояснилась, когда удалось обнаружить, что токсин связывается
с цитоплазматической мембраной хозяина, однако не у всех клонов растений. Только у восприимчивых. Значит, у восприимчивых растений в отличие от устойчивых на мембране есть некое вещество — рецептор, который связывает токсин. И такой рецептор удалось выделить. Для этого мембрану восприимчивого клона обработали детергентом, а потом расфракционировали выделенные белки. Рецептор оказался белковым тетрамером, т. е. состоял из 4 белковых субъединиц, с молекулярной массой 48 000. Токсин связывается с рецептором на восприимчивых сортах, в результате чего их клетки погибали.
Далее оказалось, что мембрана устойчивого клона тоже содержала такой же (правда, не точно такой же!) белок. Во всяком случае, белок из устойчивого клона выделялся тем же способом и имел ту же молекулярную массу.
Более того, белок-рецептор устойчивого и восприимчивого сортов не смогли различить даже антитела кролика, поскольку они связывались как с одним, так и с другим белком.
Оказалось, что различие между белками, которое не заметили даже антитела, заключается в 4 аминокислотах (лизине, серине, глицине, глютаминовой кислоте). Эти аминокислоты маскируют у устойчивого клопа центры связывания белка с токсином, что и делает растение нечувствительным к нему. Кстати, если белок из устойчивого клона тростника обработать детергентом, в результате чего его маскировка нарушается, ои приобретает свойство связываться с токсином паразита, и растение из устойчивого превращается в восприимчивое. Наоборот, если ткани восприимчивого клона тростника обработать антисывороткой, полученной на их белок-рецептор, то восприимчивый клон тростника превратится в устойчивый. Дело в том, что имеющиеся в антисыворотке антитела свяжутся
с белком-рецептором и защитят его от действия токсина.
Таким образом, из проведенных опытов можно сделать вывод, что восприимчивость растения зависит от наличия специфического рецептора, тогда как устойчивость определяется его отсутствием. Устойчивость в этом случае можно рассматривать не как активную защитную реакцию, а как пассивный признак.
Теперь относительно защиты от системы экзоферментов паразита, в первую очередь пектолитических, которым, как уже говорилось, принадлежит ведущая роль в патогенезе растений.
Нужно сказать, что стратегия обороны растения от экзоферментов у некротрофов и биотрофов не может не быть различной. Для защиты от некротрофов, для которых быстрая гибель растительных клеток является залогом их успешного паразитирования, экзоферменты паразита должны быть как можно скорее инактивированы. Защита от биотрофов предполагает не столько инактивирование фермента, сколько его регулирование. Некоторая доля активности ферментов, в частности пектолитических, может быть даже полезна для растения. Так, К. В. Васильевой было показано, что пектолитические ферменты некоторых ш разитов могут служить не только патогенными факторами, но и индукторами защитных реакций растения.
Регулирование действия пектолитических ферментов, как, впрочем, и некоторых других экзоферментов фитопатогенов, достигается с помощью нескольких механизмов. Это прежде всего регулирование на уровне образования молекул фермента либо за счет его подавления, либо, наоборот, индуцирования. Дело в том, что пектолитические ферменты у паразитов вырабатываются только в том случае, если в среде, где они растут, присутствует пектин, который и вызывает их образование. Это так называемое явление субстратной индукции. Таким образом, пектолитические ферменты индуцируются только веществами, которые им предстоит разлагать. Поэтому наличие либо отсутствие субстрата так же, как его состав, является средством регулирования синтеза этих ферментов.
Одним из способов регулирования ферментов является их катаболитная репрессия. Катаболитная репрессия — механизм экономного и целесообразного образования лишь тех ферментов, которые позволяют организму существовать с наименьшими энергетическими затратами. Предположим, у микроорганизма есть на выбор несколько источников питания. В таком случае он будет в первую очередь потреблять те вещества, использование которых ему наиболее выгодно. Например, в среду с пектином, где растет некий фитопатоген, добавляется глюкоза. Естественно, что паразит предпочтет глюкозу пектину, поскольку для использования пектина его нужно сначала разложить, а для этого образовывать ферменты, расщепляющие его пектиновые вещества, тогда как можно начать прямо с глюкозы, для превращения которой ферменты всегда имеются в наличии. Патоген начинает потреблять глюкозу вместо пектина, а пектолитические ферменты за ненадобностью перестает образовывать. Вот и получается, что добавление глюкозы (катаболита) к пектину подавляет (репрессирует) выделение паразитом пектолитических ферментов. Это и есть катаболитная репрессия.
Явление катаболитной репрессии удалось использовать для защиты от болезни платана. На места поражения наливали раствор сахара, в результате чего образование грибом экзоферментов, являющихся фактором его патогенности, прекращалось. Так сахаром удавалось вылечить болезнь.
Предохранить клеточные стенки растений от действия разрушающих ферментов возможно и с помощью их суберинизации и лигнификации. Суберин и лигнин, откладываясь в составе стенок, стабилизируют их и делают содержащиеся в них вещества недоступными для ферментов фитопатогенов.
И все-таки главным фактором защиты от экзоферментов фитопатогенов является их инактивирование с помощью различного рода ингибиторов. Важная роль в этом принадлежит фенольным соединениям, особенно продуктам их окисления, которые являются мощными факторами песпецифического ингибирования ферментов паразита.
Дело в том, что окисленные производные фенолов легко образуют комплексы с SH-, NH
2- и NH-группами белков-ферментов, в результате чего последние теряют сдою активность.
Подобный механизм ингибирования может иметь значение и в защите растений от некротрофиых патогенов, и в явлении сверхчувствительности, возникающей в ответ на проникновение несовместимых биотрофов. Например, катехины, присутствующие в клеточных стенках плодов винограда, подавляют активность пектолитических ферментов возбудителя серой гнили, причем чем больше их содержание, тем выше степень устойчивости. А продукты окисления хлорогеновой и кофейной кислот, содержащиеся в некротизирующихся в ответ на проникновение возбудителя фитофтороза тканях картофеля, подавляют пектиназы этого паразита.
Но если паразиты имеют ферменты, разрушающие клеточные стенки растения, то и растения в свою очередь тоже образуют ряд ферментов, повреждающих клеточные стенки паразитов: если стенки патогена состоят из хитина — хитиназы, если из глюкана — различного рода глюканазы и т. д. В ответ на это паразит образует ингибиторы, подавляющие действие этих ферментов. Получается, что взаимоотношения паразита и хозяина заключаются в непрерывном процессе взаимной индукции: субстрат (клеточная стейка паразита) индуцирует ее разрушающий фермент, а присутствие фермента индуцирует у паразита ингибитор, его подавляющий.
К. В. Васильевой было показано, что, для того чтобы подавить активность ферментов патогена, нужна значительно меньшая концентрация ингибиторов, чем для подавления роста самих патогенов. Например, в ксилемных сосудах вилтоустойчивых сортов хлопчатника присутствуют живые споры возбудителя болезни, однако сами симптомы болезни отсутствуют. По-видимому, присутствующие в сосудах ингибиторы подавляют экстрацеллюлярные (внеклеточные) ферменты паразита, не нарушая или почти не нарушая его жизнеспособности. Не подсказывает ли этим сама природа новый подход к использованию химических средств защиты растений, основанный на ингибировании не самих патогенов, а их патогенных факторов. Иными словами, не столько уничтожать паразита, сколько его разоружать, что вполне соответствует духу времени, в котором мы живем.
К. Т. Сухоруковым с сотрудниками была проведена серия опытов, свидетельствующих о способности растений образовывать белки-аитиферменты. Так, в ответ на введение гидролитических ферментов паразитов в растениях образовывались антиферментные ингибиторы. Причем образовывались не просто ингибиторы, а ингибиторы специфические, т. е. подавляющие только тот фермент, который вводился в растение. Ведь до сих пор речь шла только о неспецифических низкомолекулярных ингибиторах ферментов, к которым относятся окисленные производные фенолов. Эти вещества инактивируют любой фермент, лишь бы в составе его белка (а все ферменты — белки) содержались сульфгидрильные и другие реакционноспособные группы.
Вопрос о наличии специфических ингибиторов фер-меитов в растениях до сих пор оставался открытым, и лишь в последние годы удалось получить некоторые доказательства их существования. Оказалось, например, что в клеточных стенках двудольных, но не однодольных растений присутствуют высокомолекулярные белки, ингибирующие эндонолигалактуроназы паразитарных грибов. Более того, количество таких ингибиторов возрастало в ответ на инфицирование.
В последнее время выяснилось, что у растений имеются высокоспецифическиё ингибиторы белковой природы, обладающие способностью подавлять протеолитические ферменты, т. е. ферменты, расщепляющие белки (протеиназы). Такие ингибиторы обнаружены в семенах растений семейства бобовых, злаковых, сложноцветных, а также в клубнях картофеля, топинамбура, корнях турнепса. Большинство ингибиторов представляют собой белки со сравнительно невысокой молекулярной массой от 8000 до 30 000 Дальтон.
Особенностью белковых ингибиторов является их устойчивость к денатурации и протеолитическому расщеплению. Ингибиторы образуют стойкие комплексы с протеиназами, в результате чего последние теряют активность. Наличие в клетках протеиназ позволяет разрушать, а следовательно, выводить из строя нужный фермент в нужный момент, что является одним из факторов их регулировки. Но и сами протеиназы также нуждаются в регуляции, которую и берут на себя их ингибиторы.
Однако большинство ингибиторов протеиназ, обнаруженных в растениях, проявляют специфичность к протеиназам животных и микроорганизмам, и только несколько ингибиторов растений могут подавлять собственные растительные протеиназы, да и то эти ингибиторы присутствуют лишь в небольших количествах. Отчасти из-за этого, а отчасти из-за того, что большинство ингибиторов протеиназ были обнаружены в семенах и хранящихся органах растений, считается, что эти ингибиторы выполняют защитные функции, препятствуя деятельности протеиназ патогенов.
Защитная роль ингибиторов протеиназ еще более подтвердилась, когда стало известно, что после нападения колорадского жука в тканях картофеля, удаленных на некоторое расстояние от места внедрения, быстро накапливались. ингибиторы протеиназ. А затем стало очевидным, что ингибиторы протеиназ образуются не только в ответ на насекомых, а на любой тип нарушений, включая механические поранения. Эти нарушения высвобождают вещества, названные фактором, индуцирующим накопление протеиназ.
Если лист томатов подвергнуть сильному механическому поранению, то в других листьях образуются два ингибитора протеиназ, которые составляют свыше 10 % от всех растворимых белков. Накопление этих ингибиторов требует определенных условий освещенности и температуры 37 °C. Ингибитор I с молекулярной массой 39 000 состоит из 4 идентичных субъединиц, ингибитор II с молекулярной массой 25 000 состоит из 2 идентичных субъединиц. Оба сильно ингибируют протеиназы — хемотрппсип и субтилизин и слабо — трипсин. Фактор, индуцирующий накопление ингибиторов протеиназ, освобождался в воду, если листья молодых томатов срезали бритвой, а срезанный конец помещали в сосуд с водой. Этот фактор, будучи введен в листья, вызывал накопление ингибитора протеиназ с линейной скоростью.
В качестве индуктора ингибитора испытывали пидолилуксусную кислоту, кинетин, гиббереллин, абсцизовую кислоту и этилен — пи одно из них не оказывало такого действия, как фактор, индуцирующий ингибиторы протеиназ. Образовавшись в клетках, ингибитор откладывается в центральной вакуоли в виде белковых кристаллов. Там они могут сохраняться длительное время, не разрушаясь.
Большим числом исследователей доказано участие ингибиторов протеиназ в регулировании свертывания крови, процесса оплодотворения, секреции ферментов и гормонов. Их недостаток в организме может приводить к тяжелым заболеваниям. В частности, нарушение равновесия между протеолитическими ферментами и их ингибиторами является одной из причин злокачественных новообразований. Препараты ингибиторов протеиназ широко используются в качестве терапевтических средств при лечении эмфиземы легких, панкреатита, шока и других болезней.
Таким образом, наличие специфических высокомолекулярных ингибиторов ферментов, защищающих растения от фитопатогенов, сомнений не оставляет. Сомнительным остается другое, а именно способность растений в
ответ на введение в них определенного фермента образовывать строго специфические антиферменты, подавляющие только этот и никакой другой фермент. Признать это все равно, что признать за растениями способность образовывать антитела в ответ на введение антигена.
МЕХАНИЧЕСКИЕ ПРЕГРАДЫ
Для того чтобы превратить растение в источник своего питания, паразит прежде всего должен в пего проникнуть. А это не так уже и просто, поскольку каждое растение, его листья, корни, стебли, плоды, клубни и корнеплоды, одеты покровными тканями. Покровные ткани растений включают эпидермис и перидерму. Эпидермис — первичная покровная ткань растений, образующаяся из клеток, возникающих в результате первичного роста. У многих растений стебли и корпи еще дополнительно нарастают в толщину. На таких вторичных утолщениях эпидермис заменяется перидермой.
Эпидермис, образующий непрерывный слой на поверхности органа, как правило, состоит из одного ряда клеток таблитчатой формы. Наружная стенка клеток эпидермиса пропитывается липофильным веществом — кутином, представляющим собой полимер оксимонокарбоновых кислот, образующих пространственную сетку. Кутин входит также в состав кутикулы — бесклеточной пленки, покрывающей поверхность эпидермиса. На внешней поверхности кутикулы находится слой воска, благодаря чему инфекционные капли, содержащие споры патогена, не могут удерживаться на их поверхности и скатываются, что предохраняет растения от инфицирования.
Вторичная покровная ткань называется перидермой. О перидерме и цементирующем ее липофильном соединении суберине мы уже говорили в разделе «Раневой стресс».
На эпидермисе надземной части растений встречаются особые отверстия — устьица, через которые осуществляется транспирация и воздухообмен растений. На поверхности перидермы находятся чечевички — рыхло расположенные клетки, также обеспечивающие обмен воздуха и воды.
Бактерии преимущественно проникают в растительные ткани сквозь естественные отверстия на поверхности растений — чечевички, устьица, либо через раны. Вирусы могут проникать через раны, поэтому механические поранения растений, возникающие при уходе за ними, способствуют проникновению инфекции. Патогенные грибы обладают способностью к активному самостоятельному проникновению в неповрежденные растительные ткани. Правда, отдельные грибы различаются по своей проникающей способности. Группа эволюционно более древних грибов получила название «раневых», поскольку они проникают в растения только через раны. Другие грибы хотя и способны проникать через неповрежденные покровы, по все же предпочитают естественные отверстия или раны. Некоторые из них развивают на копчике гифы давление, не уступающее давлению на поршень в цилиндре мотора самолета.
Покровные ткани представляют собой не только механическую преграду на пути инфекции, но и токсический барьер, поскольку в их составе содержатся различные по природе антибиотические вещества. Об антибиотических веществах, содержащихся в составе перидермы, мы уже рассказывали.
Токсические соединения присутствуют также и в составе кутикулы и волосков, представляющих собой выросты эпидермиса. Подобные выросты обламываются при соприкасании с ними, выделяя ядовитые вещества на поверхность растительных тканей (вспомните ваши ощущения после того, как вы дотронулись рукой до крапивы).
Все, о чем мы рассказывали до сих пор, представляет собой предсуществующие механические барьеры, т. е. барьеры, имеющиеся в растении еще до того, как оно подвергается паразитарной атаке. Это так называемые конституционные барьеры. Ранее их относили к пассивным факторам иммунитета. Однако теперь стало очевидным, что такие барьеры могут активно образовываться или индуцироваться в ответ на повреждение, в том числе и на инфицирование.
Например, перидерма отграничивает область некротизированных клеток, которые погибли в результате реакций сверхчувствительности, возникшей в ответ на проникновение несовместимой расы возбудителя фитофторы в клетки картофеля.
Очаг инфекции тем самым как бы отделяется от здоровой ткани.
Иногда удается наблюдать выпадение участков некротизированной ткани в результате их окружения зоной перидермы. Так, листья миндаля, пораженные Clasterosporium carpophilum (Lev.) Aderh., на просвет кажутся как бы простреленными дробью, поскольку единичные образовавшиеся некрозы, окруженные перидермой, выпадают из ткани листа. Ограничивание очагов поражения на ветвях выглядит как изъязвление и шелушение участков. Некротизированная ткань как бы заключается в капсулу и активно вытесняется здоровыми тканями.
Выпадение пораженных очагов наблюдается тогда, когда перидерма, локализующая инфекцию, закладывается в направлении, перпендикулярном поверхности листа. В том случае, если деление происходит в периклинальном направлении, очаг инфекции как бы вышелушивается.
Очаг инфекции в растении чаще всего не удаляется, а только локализуется. Последний хотя и отграничивается перидермой, по остается соединенным со здоровой тканью. Перидерма, отграничивающая зараженный участок, выполняет три важные функции:
локализует паразита и тем самым препятствует его распространению;
прекращает приток питательных веществ из здоровых тканей растений в очаг инфекции;
защищает организм хозяина от токсических продуктов взаимодействия партнеров.
Очень интересны наблюдения внедрения возбудителя мучнистой росы в клетки ячменя и вигны. Оказалось, что еще до момента проникновения паразита в клетку, как раз под тем местом, где находится аппрессорий гриба (орган, с помощью которого паразит прободает клеточную стенку), последняя начинает утолщаться. Материал для утолщения стенки доставляется специальными цитоплазматическими пузырьками (пузырьки аппарата Гольджи), которые скапливаются под местом предполагаемого проникновения паразита. Возникает своеобразная «папилла» (бугорок), в составе которой обнаружен лигнин и кремний. Тем самым укрепляются механические свойства клеточной стенки и ее устойчивость к воздействию разрушающих ферментов гриба. Если папилла образуется своевременно, то паразит проникнуть не может, если образование папиллы запаздывает — проникновение патогена все же происходит.
Своеобразной механической преградой на пути распространения паразита служат тиллы, образующиеся в сосудах растений, пораженных возбудителями инфекционного увядания. Вилт (увядание) — одна из наиболее вредоносных болезней растений, которые поражают многие травы, деревья и кустарники. Но особенно страдает от вилта хлопчатник, который поражают грибы из родов Verticiilium и Fusarium. У заболевших растений нарушается водопроводящая система, они теряют упругость, желтеют, темнеют и, наконец, засыхают. Паразит через корпи пробирается в сосуды ксилемы, а затем с током воды распространяется по всему растению. Так происходит в восприимчивых сортах. А в устойчивых — распространению паразита по сосудам препятствуют пробки, которые образуются в сосудах выше местонахождения гриба. При этом в просвет сосуда выпячивается содержимое расположенных рядом с ннм паренхимных клеток — образуются тиллы, которые заливаются затем пектиновыми гелями. Гриб как бы замуровывается в просвете сосуда, а затем подвергается воздействию антибиотических веществ. Если такие закупорки многочисленны, то растение начинает страдать от недостатка воды. Для того чтобы этого избежать, растению приходится образовывать новые дополнительные сосуды ксилемы, которые компенсируют закупоренные.
13 последнее время все большее внимание фитоимму-пологов привлекает процесс лигнификации (одревеснения) клеточных оболочек как защитная реакция против фитопатогенов. Лигнин представляет собой гетерополимер трех фенилпропановых производных — кумарилового, копиферилового и синапового спиртов. Предполагается, что в образовании лигнина принимает участие фермент — пероксидаза и перекись водорода.
Известно, что оболочки растительных клеток в основном состоят из целлюлозы, которая образует каркас, погруженный в матрикс из веществ неуглеводной природы. Лигнин, откладывающийся в матриксе, связывается с полисахаридами клеточной стенки с помощью ковалентных связей, придавая оболочке жесткость. Важным свойством лигнина является его устойчивость к микроорганизмам: даже из числа паразитарных лишь немногие (например, дереворазрушающие грибы) могут расщеплять лигнин, да и то достаточно медленно. Поэтому одревесневшие оболочки клеток служат преградой на пути распространения инфекции. Накапливается все больше данных, свидетельствующих о том, что устойчивые растения в ответ на заражение способны лигнифицировать оболочки тех клеток, в которых ранее лигнин отсутствовал. В результате этого вокруг внедряющегося паразита образуется кольцо лигнифицированных клеток, что создает ему враждебное окружение в силу следующих причин:
лигнин повышает механическую прочность оболочек растительных клеток, в результате чего они становятся устойчивыми к проникновению паразита;
лигнин химически связывается с основными компонентами клеточных стенок и поэтому экранирует их от атаки ферментами паразита;
присутствие лигнина в клеточных стенках ограничивает не только распространение токсинов паразита по клеткам растений, но и приток питательных веществ из клеток в структуры паразита;
фенольные предшественники лигнина, так же как свободные радикалы, возникающие при его образовании, являются высоко токсичными для паразита.
И, наконец, последнее — сенсационное сообщение. Оказывается, лигнин, образуемый растением-хозяином, может откладываться в клеточной стенке гиф паразитарных грибов, ограничивая их рост, сковывая и обездвиживая. Более того, хитин — основной компонент клеточных стенок многих паразитарных грибов, с одной стороны, является как бы индуктором процессов лигнификации, т. е. сигналом, оповещающим растение, что лигнификацию следует начинать, а с другой — матриксом, где лигнин и откладывается. На какие только уловки не пускается растение, чтобы выиграть бой.
СВЕРХЧУВСТВИТЕЛЬНОСТЬ,
ИЛИ ГИБЕЛЬ ЧАСТИ ВО ИМЯ
СПАСЕНИЯ ЦЕЛОГО
Термин сверхчувствительность (СВЧ), или гиперчувствительность, т. е. повышенная чувствительность, заимствован из медицины. Это воспалительная реакция повышенной интенсивности, возникающая у предварительно сенсибилизированного организма в ответ на повторное введение того же антигена. Различают гиперчувствительность быстрого и замедленного типа. Гиперчувствительность быстрого типа возникает у животного, в крови которого уже имеются антитела на определенный антиген, поэтому его повторное введение вызывает воспалительную реакцию повышенной интенсивности. При гиперчувствительности замедленного типа готовых антител в крови еще нет, но зато есть предварительно сенсибилизированные антигеном лимфоциты, и поэтому повторное введение антигена вызывает воспалительную реакцию, однако не столь быстро возникающую.
СВЧ в фитоиммунологии имеет мало общего с медициной. Эта местная защитная реакция растений, основанная на быстрой локальной гибели части инфицированных, клеток вместе с проникшим в них патогеном. Таким образом, в медицине и фитоиммунологии под одним и тем же термином подразумевают различные явления — СВЧ у растений возникает при первичном контакте с инфекцией, тогда как у человека только в ответ на повторное введение антигена. Это своеобразная жертва частью клеток во имя сохранения целого растения, в результате чего паразит локализуется и погибает. Растение может позволить себе потерять часть своих тканей, группу клеток, лист, часть корней или даже ветвь, но остаться при этом живым, тогда как животное, как правило, не может лишиться ни одного из своих специализированных органов. Тем не менее защитная реакция, основанная на гибели части своих тканей, вряд ли может считаться эволюционно совершенной.
Впервые реакцию СВЧ обнаружил и описал ботаник из Кембриджа Г. Вард еще в 1902 г. Оп заметил, что клетки устойчивых сортов злаков настолько чувствительны к ржавчине, что погибали сразу же после установления физиологического контакта с паразитом. У восприимчивых растений инфицированные клетки оставались живыми, во всяком случае в первый период после заражения, их обмен даже активизировался, а паразит беспрепятственно распространялся по тканям. Хотя явление СВЧ впервые описал Вард, однако сам термин ввел в литературу не он, а американский фитопатолог Э. Стокман спустя 14 лет.
Кажется парадоксальным, что устойчивые растения не поражаются инфекцией именно потому, что они к ней слишком чувствительны. Казалось бы, наоборот, чувствительными должны быть восприимчивые, а не устойчивые растения. Однако именно в силу повышенной чувствительности клетка погибает, а ее гибель влечет за собой гибель проникающего патогена.
Именно благодаря сверхчувствительной реакции растения паразит не может проникнуть дальше, чем в первую или первые несколько клеток хозяина. При совместимом взаимодействии клетки хозяина и находящийся внутри них паразит длительное время сосуществуют, оставаясь живыми.
Реакция СВЧ известна также под названием гиперэргической, некрогенной или абортивной. Абортивной она называется еще и потому, что отмершая часть клеток как бы отторгается от живых механическим барьером из перидермы. При этом некротизированная ткань заключается в капсулу
и активно вытесняется здоровыми тканями (рис. 5, см. вклейку).
Чем интенсивнее выражена реакция СВЧ, тем быстрее отмирают инфицированные клетки и, наоборот, чем больше степень совместимости паразита и хозяина, тем дольше они сосуществуют. Так, клетка листа картофеля отмирает уже спустя 30 минут после того, как гифа несовместимой расы возбудителя фитофтороза соприкасается с цитоплазматическим содержимым хозяина. Чем выше степень устойчивости, тем меньшее число клеток отмирает, т. е. тем меньше та жертва, которую приходится платить инфицированному растению за выживание и освобождение от паразита, а самое главное — тем быстрее локализуется инфекция. Иногда это всего лишь одна-две потемневшие клетки, которые удается увидеть только под микроскопом.
Поскольку отмершие клетки спустя некоторое время темнеют или «некротизируются», то наличие потемневших клеток на месте инфицирования долгое время было единственным критерием для того, чтобы судить о том, произошла ли реакция СВЧ или нет. В результате любую гибель клеток, сопровождающуюся их покоричневением, стали объяснять реакцией СВЧ, тогда как причины гибели растительных клеток могут быть самыми различными. Например, гибель клеток под влиянием токсинов некротрофов также в ряде случаев сопровождается покоричневением, однако к СВЧ она никакого отношения не имеет. Мало того, чувствительность клеток к токсину свидетельствует об их восприимчивости, а совсем не об устойчивости. Создается положение, когда слишком широкое использование термина СВЧ отнюдь не способствует прогрессу фитоиммунологии, но, наоборот, даже замедляет его. Более того, мы знаем сейчас о природе СВЧ слишком мало, поскольку ученые до сих пор еще ограничиваются только использованием термина СВЧ, не предпринимая попыток понять ее природу.
Правда, фитоиммунологов давно интересовало, отчего паразит в отмершей клетке прекращает свой рост. Этот вопрос задавал себе еще Вард, глядя в микроскоп на гифы ржавчины, погибающие в некротизированных клетках. Однако, по его собственному признанию, это оказалось выше его понимания. Поскольку реакция СВЧ главным образом защищает растения от биотрофных паразитов, которые могут питаться содержимым живых клеток, то их гибель должна приводить к голодной смерти паразита. Однако вскоре стало ясным, что голодание не является основной причиной гибели паразита. Если стать на точку зрения, что паразит внутри отмершей клетки погибает голодной смертью, то гибель растительной клетки должна вызывать последующую гибель паразита. Однако это происходит далеко не всегда, т. е. гибели паразита, как правило, предшествует гибель клетки, но совсем не всякая гибель клетки приводит к гибели в ней паразита. Это заставляло задуматься, не определяется ли гибель паразита накоплением в некротизированных клетках токсических соединений. Ведь не всякая гибель клетки сопровождается образованием таких ингибиторов. Иными словами, бывает токсический некроз, приводящий паразита к гибели, а бывает нетоксический, при котором паразит продолжает жить.
Собственно, вопрос о том, как быстро погибает паразит в некротизированной клетке и всегда ли он погибает, до сих пор еще не слишком ясен. Важно то, что развитие паразита локализуется, и в силу этого он не способен к дальнейшему размножению. Поэтому главный смысл защитной реакции — СВЧ состоит не в уничтожении паразита, а в лишении его возможности спороносить.
В этой главе мы намеревались познакомить читателя только лишь с внешним проявлением столь своеобразной защитной реакции растений, какой является СВЧ, не касаясь механизмов ее сущности. Фактически при этом мы не исказили положения вещей, поскольку явление СВЧ до сих пор еще является загадкой для фитоиммунологов. Неужели растения, поражающие нас своим эволюционным совершенством, научившиеся усваивать углерод атмосферы с невиданно высоким КПД, не сумели выработать ничего лучшего, чем жертвовать частью своих клеток для того, чтобы избежать инфекций? Но так ли это на самом деле? Мы еще не раз вернемся к СВЧ в свете новых данных и попробуем показать, что подобная жертва не является столь уж бессмысленной и эволюционно несовершенной.
ИММОБИЛИЗАЦИЯ ПАТОГЕНА
В латинском словаре слово «immobilis» означает «неподвижный». Устойчивое растение обездвиживает паразита, который пытается в пего проникнуть. Прорастающие споры паразита склеиваются между собой, кончик гиф грибов перестает расти, а бактериальные клетки приковываются к клеточным оболочкам растения и постепенно теряют жизнеспособность. Как это происходит? Исследования последних лет свидетельствуют о том, что свойством обездвиживать патогенов обладают особые вещества, получившие название лектинов.
Лектины — белковые соединения, которые избирательно связывают углеводы, не вызывая при этом их химического превращения. Избирательность их взаимодействия с углеводами подчеркивается самим термином «лектин» (от латинского legere — выбирать).
Все разнообразие функциональных свойств лектинов связано с их способностью связывать различные углеводные группировки, которыми могут быть как свободные моно- и олигосахара, так и углеводные остатки в составе высокомолекулярных гликопротеидов, гликолипидов и полисахаридов. Поскольку в молекуле лектина находится не менее двух участков связывания углеводов, то лектины способны агглютинировать (агрегировать, группировать, склеивать) между собой живые клетки, в том числе и эритроциты. При этом один участок связывания в молекуле лектина прикрепляется к одной клетке, а второй — к другой.
Примерно 100 лет назад в составе бобов клещевины было найдено вещество (лектин), вызывающее агглютинацию эритроцитов и их гемолиз. С тех пор и до 1965 г. было получено только 3 изолированных лектина: рицин из семян клещевины, конканавалин А из семян конкава-валии мечевидной и лектин из фасоли. Поскольку на первых этапах все лектины были выделены только из тканей растений, они получили название фитоагглютининов. Впоследствии, когда стало ясным, что лектины присутствуют почти у всех организмов — от бактерий до млекопитающих, название лектины постепенно вытеснило фитоагглютинины.
К настоящему времени известно не менее 100 лектинов, полученных в чистом виде. Исследование лектинов стало модным направлением в науке. Ежегодно им посвящается большое число экспериментальных сообщений и обзоров. Появилась даже новая наука — лектинология.
Из числа всех обследованных растений, а их более 3000 видов, лектины найдены примерно у 1000. Практически в каждом семействе растений обнаружены представители, содержащие лектины.
Каких-либо структурных особенностей, характерных для лектинов, не существует. Большинство из них представляют собой гликопротеины с молекулярной массой от 25 000 до 400 000, но есть и исключения. Таким образом, лектины — достаточно гетерогенная группа веществ, объединенная лишь общим функциональным свойством.
Различные группы лектинов связывают различные углеводные группировки. На этом основании лектины разделяют на 6 классов. Одни из них связывают D-маннозу или D-глюкозу, другие N-ацетил-D-глюкозамин, третьи — N-ацетил-D-галактозамин, четвертые — D-галактозу и т. д. Избирательность связывания углеводов лектинами в какой-то мере напоминает реакцию антиген-антитело. Однако антитела обладают значительно более высокой специфичностью, при взаимодействии их с антигенами образуются уникальные структуры. Что касается лектинов, то один и тот же лектин может связываться со многими веществами, лишь бы они содержали одну и ту же углеводную группировку. К тому же сродство антител к антигену на несколько порядков выше, чем у лектинов.
Основным тестом для определения лектинов является агглютинация эритроцитов, поскольку на поверхности последних находится широкий набор углеводов, принадлежащий гликопротеидам и гликолипидам. Последние ориентированы таким образом, что направляют свои углеводные группы к внешней поверхности эритроцита, образуя своеобразный углеводный чехол.
Поскольку каждый лектин связывается с определенным углеводом, то добавление извне такого же углевода препятствует связыванию и соответственно предотвращает агглютинацию эритроцитов. Такие моносахара (или олигосахара), блокирующие активные центры лектинов, получили название гаптенов.
В настоящее время лектины успешно используются как инструменты для исследования поверхности клеток и биологических мембран. Особенно перспективным является исследование физиологической функции лектинов. Предполагается, что лектины участвуют в транспорте и накоплении углеводов в растениях, запасании веществ в семенах, процессах клеточного деления, распознавании пестиком типа пыльцы и других процессах. Однако особо важная роль принадлежит лектинам в процессах узнавания и взаимодействия клеток. Именно на этом свойстве и основана защитная роль лектинов в растениях, способных иммобилизовывать фитопатогены.
Иммобилизация может быть нескольких типов, например склеивание между собой проросших и непроросших спор фитопатогенных грибов или клеток бактерий. Так, лектины, выделенные из батата, вызывают слипание спор возбудителя черной гнили; лектин картофеля агглютинирует споры и цисты возбудителя фитофтороза, бактериальные клетки Pseudomonas solanacearum. Естественно, что склеенные клетки и споры не могут прорастать и распространяться и, следовательно, теряют свою вредоносность.
Предположим, споры паразита попадают на пораненный участок растительной ткани. Лектины, присутствующие в растительном соке, выступившем на месте поранения, агглютинируют их и тем самым спасают растение от заражения.
Однако тут есть одна существенная деталь. Дело в том, что лектины растения, как правило, способны склеивать клетки и споры только тех форм патогенов, к которым данное растение устойчиво. Так, лектины батата агглютинируют споры штаммов черной гнили, выделенных из растений кофе, сливы, какао, дуба, лимона, но не из батата. Лектин картофеля агглютинирует 34 авирулентных штамма Р. solanacearum, но не склеивает ни одного из 55 вирулентных. Лектин кукурузы агглютинирует 16 штаммов Erwinia sp., которые авирулентны или слабо вирулентны к кукурузе, и почти не склеивает клетки вирулентных штаммов.
Оказалось, что у вирулентных штаммов бактерий имеются вещества, препятствующие их агглютинации лектинами своего растения-хозяина. Это особого рода экстрацеллюлярная слизь, которая выделяется патогенными штаммами бактерий, окружает их чехлом и препятствует доступу лектинов к поверхности.
Собственно, логично, что вирулентные штаммы патогенов не попадают в лектиновую ловушку своего растения-хозяина. Ведь если бы они в нее попадали, то восприимчивых растений бы не существовало.
Вторым способом обездвиживания патогена является способность лектинов связывать кончик растущей гифы паразитарных грибов. Оказалось, что лектин прорастающих семян пшеницы связывается с гифами паразитарных грибов, оболочки которых состоят из полимера N-ацетил-D-глюкозамина — хитина. Лектин прикрепляется к N-ацетил-В-глюкозамину, образуя поперечные связи между цепями хитина в оболочке грибов. Интересно, что лектин связывает хитин на копчике гиф гриба, поскольку это единственное место, где хитин находится на их поверхности, так как по всей длине гифы, кроме кончика, хитин сверху покрыт слоем глюкана и, следовательно, не доступен для лектина. Но именно кончик гифы — ее наиболее ответственный участок. Здесь формируется новая клеточная стенка, синтезируются новые молекулы хитина, т. е. гифа растет на конце. Растение безошибочно находит наиболее уязвимое место у фитопатогена. Связывая цепи хитина на кончике гифы, лектин растения тем самым прекращает рост паразита.
Весьма возможно, что пшеница таким способом предохраняет свои прорастающие семена от поражения теми паразитарными микроорганизмами, на поверхности которых находится хитин (а таких патогенов чрезвычайно много). Точно так же можно предположить, что лектины семян, обладающих сродством к иным углеводам (например, глюкозе), могут стоять на страже процесса прорастания, охраняя семена от паразитов, оболочки которых состоят из глюканов. Недаром содержание лектинов в семенах настолько высоко, что вызывает удивление» исследователей.
Третьим способом иммобилизации патогенов с помощью лектинов является возможность связывания ими клеток авирулентных штаммов бактерий в сосудах флоэмы, по которым некоторые бактерии (Erwinia amylovorа) распространяются. Возможно, что лектины участвуют в иммобилизации паразита и при других сосудистых заболеваниях.
И, наконец, наиболее впечатляющими работами по иммобилизации бактерий являются исследования американского бактериолога Л. Секвейра. Он последовательно, этап за этапом на протяжении многих лет изучал поведение бактерий Р. solanacearum в тканях табака и картофеля, постепенно раскрывая тайны их взаимоотношений. Большинство фитопатогенных бактерий развивается в межклеточном пространстве растений за счет веществ, выделяемых клетками через плазмалемму и клеточную стенку.
Все началось с наблюдения, что сапрофитные бактерии, либо бактерии, авирулентные к табаку, либо, наконец, вирулентные бактерии, но предварительно убитые нагреванием, после введения в ткани табака как бы окутывались каким-то веществом и прикреплялись снаружи к клеточным стенкам. Клетки, к стенкам которых бактерии прикреплялись, погибали в процессе реакции СВЧ. Если же в ткани вводились живые вирулентные бактерии, ничего подобного не происходило — бактерии быстро развивались в межклеточном пространстве, не вызывая СВЧ-гибели растительных клеток.
Под электронным микроскопом удалось увидеть, что стенка клетки в том месте, где к ней прикрепляется бактерия, сильно изменена, а пленка, окружающая бактерию, как бы выходит из места повреждения клеточной оболочки.
События, предшествующие прикреплению бактерии, развивались следующим образом. На внутренней стороне клеточной стенки, как раз там, где находится бактерия, начинали скапливаться пузырьки (везикулы), которые, по всей вероятности, и транспортируют обволакивающий бактерию материал. Бактериальные клетки как бы попадают в ловушку, которая захлопывается. Разве это не напоминает лист росянки, покрытой капельками слизи, к которым прилипают невезучая муха или комар, которые потом с помощью ресничек притягиваются к поверхности листа?
Удалось показать, что прикрепление бактерии к клеточной стенке происходит с помощью лектина растения. Лектин табака и картофеля связывается с N-ацетил-D-глюкозамином, который присутствует в большинство оболочек грибов и бактерий. В случае Р. solanacearum лектин прикрепляется к липополисахариду, который находится на наружной мембране бактерий.
По-видимому, на иммобилизованную бактерию начинают затем воздействовать ингибиторы, поступающие из погибшей клетки. Ведь легче попасть в неподвижную мишень, чем в движущуюся. К тому же в месте прикрепления бактерии проще создать локальную токсичную концентрацию ингибиторов.
Иммобилизация бактерий на клеточной стенке растения, помимо ее обездвиживания, преследует и другую не менее важную цель — распознавание клеткой растения чужеродного начала в лице проникающего патогена. Обволакивание и прикрепление бактерий служит обязательным этапом в распознавании. Однако эта тема специальной главы, которую вам еще предстоит прочесть.
Что же происходит с вирулентной бактерией? Вот в том-то и дело, что с ней ничего не происходит. Она преспокойно питается содержимым клеток, размножается и распространяется по тканям растений. И причиной всему та самая экстрацеллюлярная слизь, выделяющаяся бактерией наружу и не позволяющая растению ни прикрепить бактерию, ни распознать ее.
До сих пор речь шла об участии лектинов в защите растения от фитопатогенов при несовместимой комбинации паразита и хозяина. Однако в целом ряде случаев связывание поверхностей взаимодействия растения и паразита посредством лектина является условием, необходимым для их совместимости. Особенно ярким примером этому служат симбионтные взаимоотношения корней растений семейства бобовых с клубеньковыми бактериями из рода Rhizobium.
Аэробные бактерии из этого рода, обитая сапрофитно в почве, образуют слизистые нити, которые проникают в корни бобовых растений через корневые волоски. Клетки корня в ответ на проникновение увеличиваются в размерах и делятся, в результате чего возникают корневые клубеньки. Бактерии проникают в клетки растения, где окружаются мембраной и образуют бактероиды. На этой стадии защитные реакции затихают и отношения партнеров становятся симбиотическими. Клубеньковые бактерии, неспособные усваивать азот воздуха пока они ведут сапрофитный образ жизни, попав в корневые клубеньки, начинают интенсивно фиксировать азот. Образующиеся при этом аминокислоты частично достаются растению.
Каждому виду бобовых соответствует своя бактерия. Так, один вид Rhizobium специфичен к сое, другой — к клеверу и т. д. Растение как бы «узнает» свой вид бактерий. Вернее, узнает не растение, а его лектины, которые связывают углеводные рецепторы на поверхности клеток только подходящего вида бактерий. Иными словами, образование или необразованно клубеньков у растения определяется тем, узнает или не узнает его лектин соответствующие углеводные группировки на поверхности бактерий. Если узнает, клубеньки образуются и азот фиксируется, если не узнает, ничего не получится.
На конечной стадии бактерии размножаются и покидают растение, а оставшиеся бактероиды перевариваются растительной клеткой и служат, таким образом, для нее ценным источником азота.
Существует много видов Rhizobium, специализированных к различным видам бобовых. Наиболее хорошо исследованы две системы симбионтов: это соя и ее специфический симбионт Rhizobium japanicum, а также клевер и R. trifoli. Даже в этих двух системах лектины локализованы по-разному, хотя и выполняют одну и ту же роль притягивания поверхностей растения и бактерии. У сои лектин находится на поверхности корневых волосков. А вот у клевера лектин выделяется из корней наружу и служит как бы мостиком между сходными углеводными группировками на поверхности клеток бактерий и соответствующего ей растения. Однако в том и другом случае лектин растения узнает свою бактерию и притягивает ее к себе.
Итак, две, казалось бы, противоположные функции лектинов во взаимодействии растения
с фитопатогенами и симбионтами основаны на процессе притягивания и распознавания микроорганизмов. Только в первом случае (фитопатогены) распознается несовместимость, в результате чего происходит СВЧ-реакция и гибель паразита, а во втором — возникает, наоборот, совместимое взаимодействие. Однако и в том и в другом случаях функция лектинов приносит очевидную пользу растению: избавляя его от поражения патогеном в первом и обеспечивая снабжение азотом во втором случае.
Чтобы покончить с ролью лектинов в процессе защиты растения, следует еще рассказать о небольшой группе токсических лектинов, хотя об этом, скорее, следовало бы говорить в разделе антибиотических веществ. Речь идет о двух токсических лектинах семян — абрине и рицине, которые рассматриваются как своеобразная защита от поедания этих семян животными и насекомыми. Устойчивые к действию гидролитических ферментов, эти лектины вызывают нарушение функции всасывания в кишечнике, повышая его проницаемость для бактериальных токсинов и продуктов гниения.
ФИТОНЦИДЫ
Кто из нас не испытывал на себе благотворное влияние леса, парка, сада, поля — наших давних и верных зеленых друзей, по праву называемых царством жизни и красоты. Растения не только снабжают нас пищей и энергией, не только пополняют запасы кислорода в атмосфере, которые отнюдь не беспредельны, они исцеляют нас от самых разнообразных болезней. О целебном действии многих растений знал еще первобытный человек. На использовании лекарственных растений основана народная медицина. Известна старая русская поговорка «лук от семи недуг». Во многих странах простудные заболевания и сейчас часто лечат домашним способом путем вдыхания распаренных растений, например лаванды, картофельной кожуры, шелухи овса и т. д. Многим эмпирическим наблюдениям народных лекарей сейчас дано научное обоснование. Современная медицина широко использует растения для получения разнообразных лекарств. И тем не менее далеко не все лечебные свойства растений еще раскрыты, многие из них продолжают оставаться тайнами живой природы.
В конце 20-х годов нынешнего столетия важное открытие сделал профессор Борис Петрович Токин. Его исследования показали, что растения выделяют вещества, способные задержать развитие и даже убивать микроорганизмы, вызывающие болезни животных и самих растений. Он назвал эти вещества фитонцидами (от греческого фитон — растение и латинского цедере — убивать) и охарактеризовал как «продуцируемые растениями бактерицидные, фунгицидные, протистацидные вещества, являющиеся одним из факторов их иммунитета и играющие роль во взаимоотношениях организма в биоценозах»
[15].
Сам автор открытия часто говорил, что фитонциды — его «незаконнорожденное дитя в науке». Дело в том, что он открыл их, уже будучи видным эмбриологом. Все эти годы, ведя фундаментальные исследования, ученый при активном участии многих исследователей и любителей природы успешно развивал и свое учение о фитонцидах, которые получили всеобщее признание в науке.
Подсчитано, например, что на 1 га лиственного леса выделяется летом 2 кг фитонцидов, хвойного — 5 кг, а можжевелового — даже 30 кг. Разумеется, дело не только в количестве, так как биологическая активность разных веществ далеко не одинакова. Например, бегония и герань снижали содержание микроорганизмов в окружающем воздухе на 43 %, циперус — на 51 %, мелкоцветная хризантема — на 66 %. Выделение фитонцидов зависит от физиологического состояния растений. Так, многие из них больше всего выделяются во время цветения. Их общее количество, содержащееся в растениях на гектаре соснового бора, достаточно для того, чтобы оздоровить воздух от патогенных микробов среднего по величине города. В этом одна из основных причин благотворного действия воздуха вблизи растений.
Уже в первых опытах было установлено, что грибы, бактерии и даже лягушки и мыши погибали в атмосфере разрезанного лука. Но гибель каждого из них наступала в разное время и зависела от продолжительности воздействия выделений, их концентрации и многих других факторов. Летучие фитонциды ряда растений, вызывающие смерть лягушки в течение нескольких минут, не могли убить некоторые инфузории даже в течение нескольких часов. Подобные различия в действии зависели от того, в какой мере подавлялись важнейшие процессы жизнедеятельности того или иного организма.
Видный советский фитопатолог Д. Д. Вердеревский. много занимавшийся изучением роли фитонцидов, пришел к выводу, что в иммунитете растений они имеют столь же важное значение, как и фагоцитоз у животных. В самом фанте более сильного выделения этих веществ пораненными растениями заложен глубокий биологический смысл. Мы уже говорили, что поранения как бы открывают ворота для проникновения микроорганизмов в растительную ткань, а интенсивным выделением фитонцидов растение перехватывает патогены еще на лету, создавая против них первую линию обороны. Ведь в природных условиях растения в той или иной степени непрерывно повреждаются от ветра, дождя, града, насекомых, птиц и т. д.
Однако далеко не все фитонциды являются летучими, имеются и нелетучие. Сосредоточены они в основном в покровных тканях растений и создают как бы вторую линию обороны против проникновения чужеродных пришельцев.
Ядовитость растений по праву рассматривается как такое же важное приспособление к их выживанию, как и другие защитные реакции. Но фитонциды не только яды, по и лекарства. Ведь недаром книга о них названа «Целебные яды растений…». Все зависит от дозы. Известно, что лекарственные вещества при повышенных концентрациях тоже могут стать ядами. Еще во времена средневековой медицины писалось: «Все есть яд, и ничто не лишено ядовитости, одна лишь доза делает яд незаметным». Так, выделения лука в малых концентрациях не только не вызывают гибель микроорганизмов, но даже стимулируют их рост. Поскольку различные организмы сильно различаются между собой по чувствительности к фитонцидам, то легко понять, что этим веществам принадлежит важная роль в биоценозах, т. е. в сообществе живых организмов, населяющих однородный по условиям жизни участок природы и связанных друг с другом определенными взаимоотношениями.
Итак, все антибиотические вещества растений являются фитонцидами. Однако они настолько разнообразны по составу, свойствам и местонахождению, что в целях упорядоченности должны быть систематизированы. Многие исследователи неоднократно предпринимали попытки систематизировать антибиотические вещества высших растений, и к настоящему времени существует несколько классификаций, разделяющих их по химическому составу, активности, механизму действия и другим признакам. Для целей настоящей книги, наверное, наиболее интересна классификация, основанная на способности этих веществ оказывать защитное действие против фитопатогенов. Именно на этих свойствах антибиотиков и построил свою систему Ингхем, который разделил все антибиотические вещества высших растений на 4 группы:
1) конституционные ингибиторы, содержащиеся в интактных (неповрежденных) растительных тканях в количествах, достаточных для подавления роста патогена;
2) полуконституционные ингибиторы, которые содержатся в интактных тканях в количествах, недостаточных для ингибирования патогенов, по накапливаются в них в ответ на повреждение до токсических концентраций;
3) полуиндуцированные антибиотики, которые отсутствуют в неповрежденных растительных тканях, но возникают в них в ответ на повреждение в результате распада более сложных нетоксических или малотоксических соединений;
4) индуцированные антибиотики ничем, кроме происхождения, не отличающиеся от веществ третьей группы: они так же, как и предыдущая группа, отсутствуют в интактных тканях растений, возникают в ответ на инфицирование, однако своим образованием обязаны не гидролизу сложных веществ, а, наоборот, синтезу антибиотических веществ из более простых (к ним относятся фитоалексины, которым будет посвящена особая глава в этой книге).
Однако одно дело содержаться в ткани в концентрациях, токсических для паразита, и совсем другое — защищать от него растения. Для того чтобы токсическое вещество защищало растительную ткань от определенного фитопатогена, оно должно оказаться в нужном месте, в нужное время, в нужной (токсической) концентрации в клетке хозяина-растения. Доказать, что в каждом конкретном случае это так, очень нелегко. Особенно сложен вопрос о пространственной локализации антибиотических веществ в клетке или межклетнике растения, так как методы определения местонахождения антибиотиков в клетке еще мало разработаны. Поскольку, как правило, они не обладают избирательным действием, то будут оказывать токсическое влияние не только на проникающего патогена, по и на растительные клетки, в которых содержатся. В силу этого содержащиеся в клетках антибиотики обычно изолированы от клеточных органоидов (митохондрий, рибосом и др.) и сосредоточены в определенных структурах клетки, чаще всего в ее вакуолях, где отделены мембраной (тонопластом) от цитоплазмы клетки и не могут поэтому вредить клеточному метаболизму. Для того чтобы вещества оказали токсическое действие на проникшего в клетку паразита, ее структура должна быть нарушена, в результате чего только и может, произойти соприкосновение антибиотических веществ с проникшим патогеном. Именно так это и происходит при поражении растительных тканей некротрофными паразитами, которые прежде, чем проникнуть в клетку, убивают ее с помощью токсических выделений и, следовательно, нарушают ее структуру.
Иное дело биотрофный паразит, который никогда не убивает (во всяком случае, на первых этапах паразитирования) клетки растений, а следовательно, не нарушает их целостности. Так содержащиеся в клетках ингибиторы продолжают оставаться должным образом пространственно отделенными от него и не могут поэтому вредить патогену. Разумеется, имеется в виду взаимодействие биотрофа с восприимчивым к нему растением-хозяином или совместимое взаимодействие. А вот при несовместимости все происходит как раз наоборот. Реакция СВЧ, которая, как вы, наверное, помните, лежит в основе
устойчивости растений к биотрофно питающимся паразитам, возможно, и рассчитана на нарушение целостности тех вместилищ, в которых хранятся яды. Таким образом, если стратегия патогенности биотрофа заключается в сохранении целостности клетки растения, то стратегия защиты от него, наоборот, состоит в ее нарушении.
Так что же представляют собой антибиотические вещества высших растений? Они представлены широким набором низкомолекулярных соединений, принадлежащих к различным химическим группам. Как правило, это вещества вторичного происхождения, природа которых определяется систематической принадлежностью растений, их продуцирующих.
Известно, что к первичным веществам относятся соединения, встречающиеся в каждой живой клетке, интенсивно превращающиеся в основном обмене веществ. Это прежде всего углеводы, белки, липиды и нуклеиновые кислоты'. Но наряду с ними встречаются и более редкие, не повсеместно распространенные вторичные вещества, которые образуются в процессах вторичного обмена и не имеют значения пи как источники энергии, пи как запасные вещества. У растений существуют тысячи таких вторичных веществ, у животных они встречаются реже. Как правило, у растений они образуются из первичных продуктов, возникающих в процессе фотосинтеза.
К антибиотикам растений принадлежат алифатические и ароматические соединения, хиноны, гетероциклические вещества, гликозиды. Сюда относятся терпеноидные соединения. многие из которых летучи, и именно они создают фитонцидное облако вокруг хвойных пород, предохраняя их древесину от древоразрушающпх грибов. Некоторые терпеноидные соединения входят также в состав воска кутикулы на поверхности листьев и плодов. Большую группу антибиотиков растений представляют различные гликозиды, молекулы которых состоят из сахарного остатка, соединенного с веществом пеуглеводной природы, называемой агликоном. В качестве агликона могут выступать фенолы, спирты, гетероциклические соединения и другие вещества.
Аглпкрны гликозидов часто бывают высокотоксичными не только для патогена, по и для живой клетки, в которой они находятся. Поэтому гликозиды и ферменты, их расщепляющие (гликозидазы), находятся в различных частях клетки: гликозиды — в вакуоли, а гликозидазы — в цитоплазме. При повреждении целостности клетки ферменты и их субстраты приходят в соприкосновение, в результате чего высвобождаются чрезвычайно токсичные агликоны.
Терпеновые гликозиды содержат в качестве агликонов тритерпены и стероидные соединения. К их числу относятся многие сапонины и гликоалкалоиды (последние встречаются у растений семейств пасленовых и лилейных). Эти соединения, особенно их агликоны, нарушают свойства клеточных мембран.
Цианогенные гликозиды, обпаружейпые не менее чем у 200 видов растений, содержат циан в качестве агликона, который и накапливается в клетках после разрыва гликозидной связи и освобождения агликона. Поскольку циан является дыхательным ядом, то патогены, устойчивые к этим ядам, обладают способностью переключать свое дыхание на обходной альтернативный путь, нечувствительный к циану.
Наибольшую группу составляют фенольные гликозиды, агликонами которых служат фенольные соединения. Последние вообще играют исключительно важную роль в устойчивости растений к фитопатогенам, особенно основанной на СВЧ-ответе. Фенолы были первыми антибиотиками, наличием которых исследователи пытались объяснить устойчивость растений к болезням. Им посвящено бесчисленное число работ. Была предложена даже фенольная гипотеза устойчивости (1929 г.), которая сейчас представляет скорее исторический интерес.
Фенольные соединения всегда присутствуют в тканях здоровых растений. Их количество обычно сильно возрастает в поврежденных тканях (инфицированных, механически пораненных, облученных УФ-лучами или подвергнутых воздействию какого-либо химического агента). Многие фенольные соединения, ранее отсутствующие в растениях, возникают в них заново, либо за счет разложения гликозидов, либо в результате образования из простых предшественников. Таким образом, фенольные соединения имеются во всех 4 группах антибиотиков по классификации Ингхема.
Отличительным свойством фенольных соединений является их способность к окислению с помощью ферментов, носящих название полифенолоксидаз, активность которых также резко возрастает в ответ на повреждение растительной ткани. Первым продуктом, возникающим при окислении полифенолов, являются хиноны — высокотоксические, крайне реакционноспособные вещества, имеющие в силу этого короткий период жизни, которые затем быстро полимеризуются.
Фенольные соединения в здоровой растительной клетке находятся в вакуоли, тогда как полифенолоксидазы — в цитоплазме.
Иными словами, субстраты и превращающие их ферменты в клетке пространственно разобщены, и поэтому их окисление, если и происходит, то в ограниченных количествах. Последнее контролируется проницаемостью тонопласта — мембраны, окружающей вакуоль. К тому же процессы окисления в клетках компенсируются процессами восстановления, и поэтому продукты окисления фенолов не накапливаются.
В погибшей или погибающей в результате реакции СВЧ клетке проницаемость мембран нарушается, а затем они разрушаются совсем. В результате фенолы бесконтрольно и необратимо окисляются полифенолоксидазами, в конечном счете образуя меланины, присутствием которых в основном и объясняется темный цвет некротизированных клеток.
Защитные свойства фенольных соединений многообразны. Фенолы, содержащиеся в интактных растительных тканях, обычно недостаточно токсичны для того, чтобы подавлять развитие паразитов. Другое дело продукты, образующиеся при их окислении, особенно хиноны. Известно, что чем большей способностью к окислению обладает фенольное соединение, тем выше его токсичность по отношению к патогенам. Однако время существования хинонов настолько мало, что делает маловероятным их непосредственное воздействие на паразита. По мере дальнейшего окисления фенольных соединений и полимеризации их токсичность обычно падает.
Даже не действуя непосредственно пд сам патоген, фенольные соединения, особенно их окисленные формы, оказывают значительное влияние на процессы патогенеза. Прежде всего это инактивация экзоферментов паразита, в первую очередь пектолитических, с помощью которых паразиты прободают клеточную стенку растений. Мы уже упоминали, что окисленные производные фенолов реагируют с сульфгидрильными, а также амино- и аминогруппами белков, чем и объясняется в значительной мере их инактивирующее воздействие на ферменты. Эти же свойства фенольных соединений лежат в основе их — способности дезорганизовывать мембранные структуры, а также нарушать ростовые процессы растений. Выше уже говорилось, что фенольные соединения, конденсируясь, образуют лигнин, предохраняющий оболочки клеток от разрушения.
Паразитарные микроорганизмы, обладающие в силу высокого коэффициента размножения широкими возможностями адаптации, обычно легко приспосабливаются к антибиотикам своего растения-хозяина. Как правило, антибиотики I, II и частично III группы мало защищают растения от поражения специфическими патогенами. В течение коэволюции со своим хозяином эти паразиты приобрели способность либо превращать и детоксицировать такие ингибиторы, либо вырабатывать толерантность к ним. Однако антибиотики этих групп выполняют весьма важную роль в механизме видового песпецифического фитоиммунитета.
Значительно сложнее осуществляется адаптация паразитов к антибиотикам IV группы — фитоалексинам, поскольку они отсутствуют в здоровой растительной ткани и возникают только в ответ на инфицирование. Именно этим антибиотикам и принадлежит ведущая роль в защите растений от фитопатогенов.
ФИТОАЛЕКСИНЫ
В 1940 г. немецкий профессор К. Мюллер со своим ассистентом Г. Бергером опубликовал статью, которой суждено было открыть новую страницу в учении об иммунитете растений к инфекционным болезням. В статье были представлены доказательства (правда, косвенные), что устойчивость растений, основанная на уже известной реакции СВЧ, связана
с накоплением в некротизированных клетках антибиотиков. Эти вещества были названы фитоалексинами (от греческого фитон — растение, алексо — отражение атаки). Одновременно в статье были сформулированы основные положения будущей фитоалексинной теории.
Мюллер с сотрудниками уже в течение ряда лет работал над выведением фитофтороустойчивых сортов картофеля. Он знал, что у фитофторы существуют различные по вирулентности расы и что один и тот же сорт картофеля, высокоустойчивый к одной расе возбудителя фитофтороза, т. е. несовместимый к ней, легко поражается другой, совместимой расой. Совместимая раса проникала в клетки картофеля, долго и успешно в них развивалась, прежде чем клетки погибали. Но те же клетки погибали сразу (некротизировались), как только в них проникала несовместимая раса, вслед за тем погибали и гифы гриба. Разыгрывалась типичная реакция СВЧ, о которой в те годы уже хорошо знали.
Мюллер задумал выяснить, а нет ли в погибших некротических клетках картофеля некоего токсического начала, которое и губит паразита. Взяли и заразили ломтики клубней картофеля вначале несовместимой расой, которая вызывала некроз клеток, а затем день спустя ту же поверхность снова заразили, но уже совместимой расой, которая в обычных условиях должна расти на этом сорте картофеля. И хотя она должна была расти, однако почему-то не росла. Тогда попробовали заразить несовместимой расой только часть ломтика, нанося взвесь спор паразита на поверхность среза в виде креста. А затем через день, как и раньше, всю поверхность ломтика заражали спорами совместимой расы. Спустя несколько дней увидели, что получилось. Весь ломтик был покрыт белым пушком споропошения гриба, кроме темных мест, на которые предварительно наносили несовместимую расу. На белом фоне заражения чернел незаряженный крест (рис. 6 см. вклейку). Совместимая раса не сумела поразить некротизированную ткань.
Итак, предварительное инфицирование несовместимой расой паразита предохранило картофель от последующего заражения. Более того, оно защищало ломтики и от второго паразита картофеля — возбудителя сухой гнили Fusarium coeruleuro. Защищенными оказались только поверхностные клетки и несколько нижних, прилегающих к ним рядов. Остальные клетки остались восприимчивыми. Вот, собственно, и весь опыт, который позволил его авторам не столько доказать, сколько предсказать существование в погибших клетках токсических веществ. Таг кпм образом, на основании очень простых по технике исполнения опытов были сделаны далеко идущие широкие обобщения.
А теперь предоставим слово самим автором и послушаем, какие выводы они сделали из своего эксперимента. Воспроизведем их почти дословно, но в более популярной форме.
1. Причиной гибели паразита в клетках устойчивого сорта картофеля не может быть присутствие антибиотического вещества, которое уже имелось в клубне до его инфицирования. Ею не может быть и отсутствие в клетке веществ, необходимых для развития паразита. Причина гибели заключается в изменении состояния клеток хозяина, которые контактировали с паразитом. В результате их изменения образуется новое антибиотическое соединение, которое мы называем «защитной реакцией».
2. Защитная реакция связана с живыми клетками хозяина.
3. Антибиотическое вещество возникает либо активируется в клетках хозяина и может быть конечным продуктом «некробиоза», вызванного паразитом.
4. Это еще не выделенное и поэтому пока гипотетическое защитное вещество, названное фитоалексином, не специфично по своему действию. Оно подавляет по только фитофтору, по и другие грибы, паразитирующие на картофеле, а также сапрофитные грибы. Различные виды паразитов различаются по чувствительности к фитоалексину.
5. Судьбу паразита, так же как и хозяина, решает чувствительность его клеток к некоему соединению, образованному фитофторой, причем чем больше чувствительность клеток растения, тем выше его устойчивость. Способность клеток хозяина реагировать определена гепотинически и проявляется в скорости, с которой образуется защитное вещество.
6. Защитная реакция проявляется только в тканях, оккупированных грибом, а также непосредственно прилегающих к ним, а не распространяется на весь индивидуум.
7. Если в устойчивых сортах происходит иммунизация части ткани, пораженной патогеном, то в восприимчивых наоборот: клетки хозяина, пораженные вирулентной расой, через некоторое время становятся чувствительны даже к грибам, неспособным поражать интактные клубни картофеля. При этом различные грибы способны поражать зараженные фитофторой ткани в различной степени.
8. Состояние устойчивости не наследуется; наследуется только способность к его приобретению, что происходит после контакта с паразитом. В результате этого срабатывает механизм, превращающий часть атакуемых паразитом клеток из состояния индифферентности в состояние устойчивости.
Прошло 45 лет. Многое прояснилось, некоторые выводы пришлось пересмотреть и уточнить. Но в целом они выдержали испытания временем.
Сейчас, оглядываясь назад, становится ясным, что в 1940 г. было открыто новое биологическое явление — способность растительных тканей образовывать в ответ на инфекцию антибиотические вещества, подавляющие развитие патогена.
А что же было дальше? А собственно, дальше ничего не было, по крайней мере в течение последующих 18 лет до того момента, когда тот же Мюллер выступил с новой статьей, в которой изложил метод быстрого и простого обнаружения фитоалексинов. Метод получил название капельных диффузатов.
Он очень прост. Из зеленых бобов фасоли осторожно вынимают семена, а на их место в семянные впадины помещают капли воды со спорами патогена. Споры прорастают, выделяя вещества, проникающие в ткани стручка. В ответ на это в растительной ткани образуются фитоалексины, которые диффундируют в инфекционные капли с тем, чтобы подавить в них рост паразита. Если через 2 суток такие капли собрать, проросшие споры удалить, то это и есть диффузат. Чем хороши стручки? Тем, что на их внутренней поверхности нет кутикулы, и фитоалексины поэтому легко проникают в инфекционные капли из тканей, в которых образуются (рис. 7).
 Рис. 7.
Метод капельных диффузатов (пояснения в тексте)
Рис. 7.
Метод капельных диффузатов (пояснения в тексте)
Последствия нового метода вскоре дали себя знать. Уже через год таким способом из бобов гороха был выделен первый фитоалексин, который был назван пизатин (от латинского Pisuni — горох). Честь открытия принадлежит австралийскому фитоиммунологу Круикчанку и его сотруднице Перрин. Еще через год эти же авторы получили другое вещество, на этот раз из фасоли, и назвали его фазеоллином (от латинского Pliaseolis — фасоль). Оба вещества оказались хроманокумаранами.
И тогда стало ясным, что два фитоалексина уже давно известны, только никому не приходило в голову их так называть. Это фуранотерпеноид — иномеамарон, который еще в 1943 г. обнаружил в батате Хиура, и фенантрен — орхинол, выделенный Гойманом и Керном из клубней орхидей в 1945 г. Пятым и шестым были сесквитерпеноиды картофеля — риширии и любимин, выделенные в Японии в лаборатории Томияма и в СССР, в нашей лаборатории.
Ежегодно появлялись работы, которые захватывали все новые и новые ряды исследователей. В их числе оказались и авторы этих строк. Затем поток сообщений принял прямо-таки лавинообразный характер. В среднем 25–30 % всех сообщений в мировой литературе по фитоиммунологии посвящалось и посвящается фитоалексинам. Последние стали самой горячей точкой фитоиммунологии.
Вот что мы имеем сегодня. Сейчас уже открыты около 200 фитоалексинов. Они обнаружены у голосемянных и покрытосемянных, однодольных и двудольных, в число которых входят растения, принадлежащие более чем 20 семействам. Среди них бобовые, пасленовые, мальвовые, розоцветные, вьюнковые, зонтичные, сложноцветные и др. Долгое время никак не удавалось обнаружить фитоалексины у злаковых и тыквенных. Во всяком случае, их не удавалось обнаружить теми методами, с помощью которых у представителей других семейств это сделать удалось. Однако в самое последнее время появились сообщения, что и у растений этих двух важнейших семейств такие вещества имеются.
До настоящего времени нет общепринятой номенклатуры. А каждый исследователь, открывающий новый фитоалексин, естественно, хочет как-то его назвать: кто по названию вида растений, из которого он выделен, кто по наименованию сорта, а кто — в соответствии с химической природой. Например, ришитии и любимиц получили свои названия от сортов картофеля (Ришири и Любимец), из которых были впервые выделены, фитуберип — от наименования патогена — фитофтора и видового названия культурного картофеля — Solannm tuberosum, солаветивон — от родового наименования Solannm.
До сих пор много споров вызывает само определение фитоалексинов. Некоторые ученые предлагают их называть стрессовыми метаболитами, т. е. веществами, возникающими в растениях в ответ на неблагоприятные воздействия. Ведь стрессы есть и у растений тоже. Но, во-первых, эти вещества возникают совсем не на всякие стрессы, а во-вторых, в растительных тканях, попавших в условия стресса, возникает много разных веществ, по совсем не все они являются фитоалексинами.
В определении фитоалексинов обязательно должна быть отражена их главная биологическая функция — защита растений. Поэтому нам представляется, что фитоалексины — низкомолекулярные антибиотики высших растений, которые практически отсутствуют в их здоровых тканях, возникают в ответ на контакт с фитопатогенами и при быстром достижении антимикробных концентраций могут выполнять защитную роль в фитоиммунитете. Поскольку слишком много соединений-самозванцев претендуют быть названными фитоалексинами, канадский исследователь Стоессл установил, что эти вещества должны иметь 4 обязательных признака:
иметь умеренно низкую молекулярную массу;
обладать фунгитоксичностью;
отсутствовать в здоровой ткани растений, но индуцироваться в ней патогенами;
синтезироваться заново из мелких универсальных блоков (о последнем мы уже говорили, когда обсуждали классификацию антибиотических веществ).
Но вот насчет фунгитоксичности. Мюллер хотя и обнаружил фитоалексины в зараженных грибом тканях картофеля, однако назвал их не антигрибными, а антибиотическими веществами, подавляющими рост патогенных микроорганизмов, а не только грибов. До самого последнего времени выделение фитоалексинов рассматривали только как средство защиты растений от паразитарных грибов, однако сейчас установлено, что многие из них обладают антибактериальным и даже антинематодным действием.
Сюда относятся вещества, различные по химическому строению. Они являются конечными продуктами того измененного обмена веществ, который возникает в растительных тканях в ответ на заражение. А у растений разных таксономических групп происходят разные изменения обмена, в результате чего у разных растений образуются его разные конечные продукты. Например, у пасленовых под влиянием заражения начинают образовываться сесквитерпеноидные вещества, которых раньше в здоровых тканях не было, у бобовых — изофлавоноиды, у сложноцветных — полиацетилены.

Получается, что структура фитоалексина является как бы признаком определенного таксона: вида, рода, а иногда и семейства растений.
Однако у всякого правила есть свое исключение. Например, только что обнаружили, что у томатов, наряду с сесквитерпеноидными, существуют и фитоалексины ацетиленовой природы.
У каждого растения образуется не один фитоалексин, а несколько. Однако все они представляют собой группу химически родственных соединений. То, что их много — очень важно, поскольку микроорганизмы, как известно, легко приобретают нечувствительность к антибиотикам. Но если привыкнуть к одному антибиотику относительно легко, то к двум уже труднее, а к нескольким — совсем трудно. К тому же образующиеся вещества хотя и близкие родственники, но часто обладают различными способами воздействия на патоген, т. е. действуют вкупе, дополняя друг друга. Так, у картофеля, зараженного фитофторой, обнаружено около десятка стрессовых метаболитов, примерно столько же их в инфицированных тканях фасоли. Правда, не все из них еще завоевали право называться фитоалексинами, поскольку пока недостаточно исследованы.
Еще Мюллер заметил, что фитоалексины являются «продуктом некробиоза» клеток хозяина. Он и здесь не ошибся, поскольку сейчас установлена связь между некрозом клеток растений, погибающих в ходе реакции СВЧ, и образованием этих веществ. Правда, продуктом некроза фитоалексины быть не могут, поскольку их синтез требует энергии и поэтому не может протекать в погибших или погибающих клетках. Скорее погибающие клетки подают своеобразный сигнал для синтеза.
Синтез фитоалексинов берут на себя здоровые клетки, окружающие некроз, так называемые «бордюрные» клетки. Именно в них эти вещества и образуются, а затем направляются навстречу опасности — в некротизированные клетки, в которых находится паразит. Нечто вроде армии, которая мобилизуется в тылу и посылается на фронт. Кстати, для того чтобы защищаться, тыл определенного размера необходим. Например, ломтик фитофтороустойчивого картофеля теряет свою устойчивость, если он слишком топкий. Подсчитано, что для успешного сопротивления нужен слой не менее чем из 15 рядов клеток, иначе при заражении не из чего будет синтезировать фитоалексины. Их перемещения из здоровых клеток в некротизированные показаны для устойчивых сортов батата, картофеля, бобов и сои. Что руководит этим направленным током в некротизированные клетки — никто не знает. Может быть, у некротизированных клеток нарушается проницаемость клеточных мембран, в результате чего через них легче проникают фитоалексины? Ведь большинство из них липидорастворимые вещества, а чем лучше вещество растворяется в липидах, тем больше у пего шансов пробраться через мембрану.
Фитоалексины не только подавляют прорастание спор и рост фитопатогенов, но некоторые из них, кроме того, ингибируют активность их экзоферментов, иными словами, и убивают, и разоружают агрессора.
Считается, что фитоалексины являются антибиотиками «широкого спектра действия», поскольку они токсичны не только для грибов и бактерий, но и для некоторых нематод и насекомых, а также для клеток растений и животных. Известно, например, что ипомеамарон батата повреждает печень, является нервно-паралитическим ядом. Госсинол хлопчатника вызывает застой и отек органов, падение гемоглобина, разрушение эритроцитов, блокирует спермообразоваиие. Некоторые фитоалексины бобовых вызывают гемолиз эритроцитов.
Растительные клетки не способны выдерживать высокие концентрации фитоалексинов, может быть поэтому они и накапливаются в токсических дозах в погибших клетках, которым уже нечего терять. Вероятно, поэтому их передвижение из здоровых клеток в некротизированные может происходить, минуя содержимое клеток, за пределами их цитоплазматической мембраны, т. е. апопластно по взаимосвязанной системе клеточных стенок и межклетников. Такой способ передвижения удалось установить у клубня картофеля, инфицированного возбудителем фитофтороза.
Поскольку фитоалексины принадлежат к различным химическим классам, трудно ожидать, что они будут иметь единый механизм токсического действия. Считается, что фитоалексины действуют не на специфическую мишень в клетке (в отличие от микробиальных антибиотиков и системных фунгицидов), а подавляют сразу многие звенья обмена, т. е. являются мультисайтовыми ингибиторами. Однако у многих из них есть общее свойство — они повреждают мембраны, прежде всего окружающую клетку цитоплазматическую мембрану.
Большинство устойчивых растений в ответ на инфицирование образует большее количество фитоалексинов, чем восприимчивые.
Однако при оценке защитной роли фитоалексинов нужно учитывать не только их суммарное количество, по также скорость образования и местонахождение (локализацию фитоалексинов в растении). Правда, сделать это очень трудно.
Представьте себе сосуд ксилемы (трахею) хлопчатника, по которой снизу вверх от корней к листьям вместе с током воды распространяются споры возбудителя инфекционного увядания (вилта). Что происходит в устойчивом растении? Просвет сосуда быстро перекрывается за счет образования в нем тилл и пробок, о которых уже шла речь. В закупоренном сосуде прекращается ток воды и споры попадают в ловушку, накапливаясь в пространстве сосуда ниже места закупорки. Вот тут-то за них и принимаются фитоалексины. Локальное местонахождение спор создает возможность создания вокруг них токсических концентраций антибиотиков, которые быстро с ними расправляются.
Вот так в хлопчатнике реализуется принцип «в нужное время, в нужном месте и в нужном количестве». Кстати, обратите внимание, что реализовать этот принцип возможно только, если в дело включаются два типа защитных реакций: формирование механических преград в виде пробок и образование фитоалексинов. Каждая реакция в отдельности не смогла бы сработать. Это и есть синергизм защитных реакций растений, при котором одна из них дополняет другую.
А в восприимчивом сорте хлопчатника споры паразита, не встречая препятствий со стороны растений, спокойно распространяются по его сосудам. Образование фитоалексинов происходит даже в большем объеме, чем в устойчивом сорте (во многих местах, где рассеяны споры паразита), да только это мало помогает. Восприимчивое растение тратит много сил на образование фитоалексинов, но не в состоянии вовремя остановить распространение спор и создать вокруг них токсическую зону, поэтому паразиту удается захватить все растение.
Известны данные, когда удаление фитоалексинов из тканей устойчивого растения превращает его в восприимчивое и, наоборот, добавление этих веществ к восприимчивому растению делает его устойчивым.
Если растение выдержать в плохих для него условиях (при повышенной температуре, недостатке кислорода), то его болезнеустойчивость падает и параллельно уменьшается способность образовывать фитоалексины. Б хороших условиях, наоборот, возрастает как устойчивость, так и образование антибиотиков. Интересно, что если после временного воздействия повышенной температуры растение вновь перенести в условия нормальной температуры, где оно через некоторый срок приходит в себя, то вместе с возвращением болезнеустойчивости восстанавливается и способность образовывать фитоалексины.
Если устойчивые растения обработать определенными ингибиторами обмена, которые, не затрагивая паразита, подавят в растительных тканях образование ферментов, ответственных за синтез фитоалексинов, то такие растения из устойчивых превращаются в восприимчивые. Более того, описаны случаи, когда устойчивое растение, в котором тем или иным способом подавлена способность к образованию фитоалексинов, становится восприимчивым не только к своему специализированному патогену, но и к тем микроорганизмам, которые никогда ранее это растение не поражали. Иными словами, фитоалексины защищают растение не только от его специализированных паразитов (сортовая устойчивость), но и от неспециализированных к нему патогенов (видовой иммунитет).
Можно привести еще много примеров того, как коррелирует устойчивость растений с их способностью образовывать фитоалексины. Однако ученые со свойственной им скрупулезностью требуют все новых и новых доказательств участия этих веществ в защите растений. Корреляция между их образованием и устойчивостью к болезням получена для десятков сортов растений. Однако когда сравнивают между собой разные сорта растений, всегда есть опасность, что их способность образовывать фитоалексины определяется какими-либо иными свойствами сорта, а не только его устойчивостью к тому или иному патогену. Генотип сортов может оказаться настолько различным, что их вообще трудно сравнивать между собой но устойчивости.
Более достоверные сведения можно получить на изогенных линиях растений. Эти линии выравнены по всем генам, кроме одного, в нашем случае по гену устойчивости к той или иной болезни. Правда, на практике получаются не изогенные, а скорее близкие к изогенным линии. Оказалось, что устойчивые линии кукурузы, льна, перца, томатов, сои образуют фитоалексины вдвое, втрое, а то и в десять раз интенсивнее, чем соответствующие восприимчивые линии.
Собственно, что более всего смущает фитоиммунологов? Да тот же самый уже знакомый нам вопрос: а всегда ли в устойчивых растениях фитоалексины образуются в нужном месте, в нужное время и в нужном количестве, чтобы покончить с паразитом? Таких данных пока еще очень мало. И не потому, что поведение этих веществ редко соответствует такому требованию, а потому, что методы определения их локального местонахождения еще не разработаны. К тому же о токсичности фитоалексинов мы пока еще судим только по тому влиянию, которое они оказывают на патогены, будучи добавленными в среду его роста. А будет ли эта величина соответствовать их токсичности в растительных тканях? Например, возбудитель корневой гнили гороха индуцирует в его тканях концентрации пизатина, в несколько десятков раз превышающие те дозы, при которых рост гриба полностью подавляется в пробирке, тем не менее это не мешает ему развиваться в содержащих пизатин тканях. Значит, токсичность веществ в растениях и в пробирке может не совпадать?
Тем не менее из числа всех биохимических показателей, известных нам, образование фитоалексинов лучше остальных коррелирует со свойством болезнеустойчивости. И это краппе важно, поскольку для решения ряда практических задач, таких, как селекция, интродукция, сортоиспытание растений, необходимы показатели, характеризующие устойчивость растительных тканей к болезням. Насколько нам известно, уже сейчас не без успеха предпринимаются попытки использовать фитоалексины как объективный критерий устойчивости растений.
Хотя роль фитоалексинов в сортовой устойчивости более очевидна, чем других групп антибиотиков, тем не менее многие специализированные патогены обладают способностью и их преодолевать. Если бы это было не так, то на свете не существовало бы пи специализированных патогенов, ни восприимчивых к ним сортов растений. Многие специализированные некротрофы обладают способностью разлагать образовавшиеся фитоалексины своего растения-хозяина — пизатин, фазеоллин, помеамарон, вайероновую кислоту, капсидиол и др. В основе процесса лежат самые разнообразные химические превращения — окисления, гидроксилирования и др. Часто детосикация осуществляется фитопатогеном в несколько этапов, причем каждый последующим продукт ферментативного превращения обладает меньшей фунгитоксичностыо, чем предыдущий. Например, пектрия галлообразующая детоксицирует фитоалексин яблони — бензойную кислоту в 2 этапа. Вначале бензойная кислота превращается в геоксибензойную, при этом токсичность по сравнению с исходной падала в 20 раз. Затем геоксибензойная кислота деградировала в протекатеховую, токсичность которой в 40 раз ниже исходной.
Интересно, что различные грибы детоксицируют один и тот же фитоалексин различными путями, а один и тот же патоген превращает родственные по строению фитоалексины одним и тем же путем.
Однако детоксикация фитоалексинов своего растения-хозяина не является единственным путем их преодоления. Другой возможностью является способность специфических патогенов препятствовать образованию фитоалексинов в растении. Это может достигаться за счет их способности проникать в растение незамеченным, о чем' пойдет речь при дальнейшем изложении. Такой механизм преодоления защитных реакций растения патогеном наиболее вероятен среди партнеров, взаимоотношения которых укладываются в систему геи — на — ген, т. е. для высокоспециализированиых биотрофов.
ЛИШЕНИЕ ПАТОГЕНА САМОГО НЕОБХОДИМОГО
Если целью паразита является превращение хозяина в источник своего питания, то растение, наоборот, стремится этого не допустить. Отсюда следует, что если растение сумеет превратить свои ткани в неудобоваримые, а то и просто в ядовитые для паразита, то оно перестанет им поражаться.
Малоспециализированные некротрофы слабо зависят от состава питательных веществ поражаемого ими растения. Прежде всего их биосинтетические возможности значительно шире, они больше привыкли рассчитывать на себя, чем на питающее их растение. Наоборот, высокоспециализированпые биотрофы потому и не могут существовать вне живой клетки, что используют готовые органические вещества, которые в них синтезируются. Иными словами, такие паразиты заставляют растения биосинтезировать определенные соединения вместо себя, а сами часто даже утрачивают эту способность. И одни из механизмов уже описанной реакции СВЧ как раз и состоит в лишении паразита необходимого ему питания и в навязывании голодовки.
Чем теснее приурочен биотроф к обмену своего растения-хозяина, чем более он от него зависим, тем более вероятно, что такая зависимость может быть нарушена при том или ином изменении обмена растения. Известно, что биотрофы потенциально более подвержены вымиранию, чем некротрофы, поскольку целиком зависят от судьбы своего хозяина (гибель растения приводит и к гибели его облигатного паразита). Может быть, именно поэтому облигаты так дорожат жизнью своего хозяина, не оказывая на пего летального воздействия. Приспособленность паразита к составу клеток своего растения-хозяина, как к единственному источнику питания, носит название пищевой специализации.
Конечно, паразиты в меньшей мере испытывают дефицит от наличия в составе растений первичных соединений — углеводов, липидов, белков, которые встречаются в каждой живой клетке и превращаются в основном обмене веществ. Более вероятно, что пищевая специализация определяется наличием либо концентраций относительно редко встречающихся соединений, так называемых веществ вторичного обмена.
В середине 50-х годов Э. Гарбер предложил гипотезу устойчивости растений, основанную на зависимости паразита от состава питательных веществ хозяина. Работая с 7 мутантами бактерий, зависимыми по ряду аминокислот, он пришел к выводу, что растение окажется устойчивым, если не обеспечивает патогена необходимым питанием. В том случае, если оно способно прокормить паразита, имеет место восприимчивость.
Согласно гипотезе Гарбера, устойчивость растений определяется отсутствием в их тканях некоторых физиологически активных веществ, которые паразит не в состоянии самостоятельно синтезировать. Это могут быть некоторые витамины, незаменимые аминокислоты, ростовые вещества и ряд других соединений. Подобная гипотеза фитоиммунитета получила название питательно-тормозящей.
Еще до Гарбера близкая концепция устойчивости была сформулирована советским исследователем, основоположником патофизиологии растений К. Т. Сухоруковым, который провел серию интересных исследований, касающихся роли питания паразита в устойчивости хлопчатника к возбудителю инфекционного увядания и картофеля к возбудителю фитофтороза. Вот что Стало ясным после его работ. Для роста фитофторы необходимы растворимые сахара, однако в картофеле они находятся в неудобоваримой для фитофторы форме — в виде крахмала. Казалось бы, чего проще: фитофтора должна выделять фермент — амилазу, расщепляющую крахмал до сахаров, и проблема решена. Но в том-то и дело, что амилазы у фитофторы пет, зато она есть у картофеля, а следовательно, ею можно воспользоваться. Однако опять неудача: амилаза у картофеля хотя и есть, по она заблокирована особым ингибитором — систоамплазой. Паразит преодолевает и эту трудность: освобождает амилазу из блока, с ее помощью расщепляет крахмал и потребляет образующиеся при этом моносахара. Видите, как сложно и с каким дальним прицелом управляет фитофтора в своих интересах обменом пораженного картофеля. Правда, в конце паразиту все же приходится расплачиваться. На заготовленных им для себя сахарах поселяются сапрофиты, которые раньше не могли поражать картофель, и постепенно вытесняют неспособную с ними конкурировать фитофтору.
Обмен биотрофа настолько тесно приспособлен к обмену его растения-хозяина, что всегда существуют некие узкие места, в которых паразит особенно уязвим. Вопрос только в том, чтобы эти места обнаружить. Чаще всего узкие места находит само растение-хозяин, изменяя свой обмен таким образом, что паразит лишается некоего фактора, необходимого ему для существования. Тем самым растение как бы само указывает на узкий участок паразитирования патогена, а человеку остается только попять, как оно это делает. Читатель простит нас за излишний антропоморфизм. Конечно, на самом деле все происходит по-другому: некая мутация превращает растение в устойчивое и тем самым позволяет избежать заражения. Отбор подхватывает эту полезную мутацию, благодаря чему мутанты и получают преимущество в потомстве.
В качестве примера можно привести наши работы по изучению того же возбудителя фитофтороза, который в ходе сопряженной эволюции со своим хозяином — картофелем утратил способность самостоятельно синтезировать стерины. И не потому, что они ему не нужны. Стерины фитофторе просто необходимы. Во-первых, они являются обязательным компонентом цитоплазматических мембран эукариотов, создавая их прочный каркас. Во-вторых, выполняют функции гормонов либо их предшественников. Оказалось, например, что без стеринов фитофтора не может образовывать споры. Удалось даже установить ту мишень, которая нарушается в отсутствие стеринов: без них не происходит формирования цитоплазматической мембраны зооспор паразита, т. е. нарушается его главный процесс — воспроизведение.
Паразит утрачивает способность самостоятельно синтезировать стерины не потому, что последние ему не нужны, а потому что он может воспользоваться готовыми стеринами, имеющимися в составе хозяина-картофеля. Зачем утруждать себя синтезом, если можно без лишних хлопот получить сразу готовый продукт? Более того, фитофтора сумела приспособиться и к качественному составу стеринов своего хозяина. Главным стершим картофеля является β-ситостерин, который составляет более половины от всех имеющихся стеринов. Именно этот стерин стал самым «любимым» для фитофторы, поскольку он наиболее соответствует ее питательным потребностям.
Вскоре последовал ответный удар со стороны растения. В инфицированном картофеле обмен изменился так, что стерины синтезироваться перестали. Конечно, не во всем растении (прожить без стеринов картофелю невозможно), а только в зоне инфицирования. Особенно резко сокращалось количество любимого фитофторой β-ситостерина. Фитофтора оказалась как бы в кольце бесстериновых клеток, а следовательно, была лишена возможности размножаться.
Но и этого оказалось мало. Те вещества, которые ранее использовались в биосинтезе в качестве предшественников для стеринов, теперь стали предшественниками для фитоалексинов, ингибирующих развитие паразита. Дело в том, что сесквитерпеноидные фитоалексины картофеля и стерины образуются из общих предшественников единым биогенетическим путем (рис. 8). На определенном этапе биогенеза находится поворотный пункт: если метаболизм пойдет в одном направлении, образуются стерины, если в другом — сескитерпеноиды. Поскольку первых в зоне инфицирования фитофтороустойчивого картофеля нет, а вторые присутствуют в избытке, можно полагать, что вместо стеринов образуются токсические фитоалексины. Так что же получается? Паразит не только голодает, но и отравляется? Два удара по паразиту и оба достигаются одним и тем же изменением процесса метаболизма.
 Рис. 8. Биогенез терпеновых соединений в картофеле, инфицированном совместимой и несовместимой расами возбудителя фитофтороза
Рис. 8. Биогенез терпеновых соединений в картофеле, инфицированном совместимой и несовместимой расами возбудителя фитофтороза
Но и это еще не все. Казалось бы, несмотря на общее происхождение обеих защитных реакций, каждая из них воздействует на паразита самостоятельно: отсутствие стеринов делает невозможным спороношепие паразита, а фитоалексины оказывают на него токсическое воздействие. Но это только на первый взгляд. В отсутствие стеринов ослабляется ригидность цитоплазматических мембран паразита, и он становится значительно чувствительнее к воздействию фитоалексинов. Так, если для полного подавления роста паразита, содержащего стерины, необходимо 200 мкг/мл ришитина, то для гибели того же паразита, лишенного стеринов, их нужно только 6–7 мкг/мл, т. е. в 30 раз меньше. Для бесстеринового паразита фитоалексины значительно более опасны, и, для того чтобы его погубить, растению нужно синтезировать их значительно в меньшем количестве. Недаром считается, что стерины защищают организм от неблагоприятных факторов среды — температурного стресса, ионизирующего излучения, воздействия токсических веществ. Усиленный синтез стеринов представляет собой приспособительную реакцию организма на выживание.
Таким образом, изменение терпеноидного биогенеза, происходящее в фитофтороустойчивом картофеле, наносит по паразиту не двойной, а тройной удар:
1) гриб лишается стеринов, необходимых ему для размножения и патогенных свойств;
2) взамен стеринов образуются токсические фитоалексины;
3) лишенный стеринов гриб становится чувствительнее к фитоалексинам.
Итак, отдельные защитные реакции растения действуют сообща, взаимодополняя и усиливая друг друга. Результат их воздействия представляет собой пример синергизма, поскольку, взятые вместе, они оказывают больший эффект, чем сумма эффектов каждой реакции в отдельности. Да так, собственно, и должно быть, поскольку каждая из защитных реакций ослабляет паразита. А ослабленный предыдущим
воздействием паразит, естественно, становится более уязвим для последующего удара.
Как вы, наверное, помните, Гарбер считал, что устойчивость растения объясняется отсутствием в его тканях того или иного необходимого паразиту вещества еще до контакта с патогеном. Растение с дефицитом такого вещества уже должно было быть устойчиво к данному паразиту. Однако из сказанного выше следует, что растение до встречи с паразитом может содержать все продукты, необходимые для того, чтобы его прокормить.
Но как только паразит, «соблазнившись» богатыми растительными запасами, пробует проникнуть в растение, последнее изменяет свой обмен так, что в его тканях перестают образовываться нужные паразиту вещества, взамен чего начинают синтезироваться яды. Теоретическая возможность подобного механизма устойчивости была предсказана еще В. Н. Эфроимсоном, который предполагал, что устойчивость растений может проявляться в изменении структуры того гена, который ответствен за выработку продукта, необходимого для существования паразита, и превращения его в соединение, тормозящее деятельность патогена или активность его ферментов.
Приспособленность паразита к стероидному обмену хозяина не может не удивлять. Мало того, что паразит утратил способность самостоятельно синтезировать стерины с тем, чтобы пользоваться стеринами картофеля, а фитофтора приспособилась к качественному составу стеринов хозяина (более всего «полюбив» главный стерин-β-ситостерин), оказалось еще, что развитие паразита точно повторяет содержание стеринов в вегетирующих растениях картофеля.
Возбудитель фитофтороза картофеля, согласно теории иммуногенеза М. С. Дунина, относится к болезням нисходящей фазы онтогенеза. Фитофтороз впервые появляется к моменту цветения картофеля и достигает своего максимального развития в конце лета. Недаром эта болезнь называется еще поздней гнилью. Динамика появления и накопления стеринов в листьях точно отражает появление и развитие болезни: содержание стеринов во всходах очень низкое, оно резко возрастает к началу цветения и достигает своего максимума к началу естественного отмирания ботвы (рис. 9). Получается, что слабо функционирующий стериновый биогенез ограничивает развитие гриба на первых этапах вегетации, а возрастание содержания стеринов позволяет ему распространяться и спороносить на более поздних фазах развития растений.
 Рис. 9. Соответствие накопления стеринов развитию фитофтороза
Рис. 9. Соответствие накопления стеринов развитию фитофтороза
ЧТО ТАКОЕ КСЕНОБИОТИКИ
И КАК ОНИ УНИЧТОЖАЮТСЯ
Лекарственные вещества и промышленные загрязнения, пестициды и продукты бытовой химии, пищевые добавки и консерванты — вот тот поток чужеродных соединений, который со всевозрастающей силой обрушивается на нашу планету и живущие на ней организмы. Эти синтетические компоненты добавляются к огромному разнообразию чужеродных веществ природного происхождения, которые образуются растениями, грибами, бактериями и другими организмами. Недаром эти соединения получили название «ксенобиотики», т. е. «чуждые жизни».
В столь острой ситуации все живое давно бы оказалось под угрозой гибели, если бы не обладало механизмами, неустанно поддерживающими свою «химическую чистоту». Организмы высших животных и человека в ответ на введение антигенов образуют антитела и тем самым нейтрализуют их воздействие на организм. Однако антигенными свойствами, т. е. способностью вызывать образование антител, обладают только высокомолекулярные ксенобиотики — белки, гликопротеиды, некоторые полисахариды и нуклеиновые кислоты. А как же обезвреживаются ксенобиотики низкомолекулярные? Исследования показали, что такую функцию берет на себя цитохром Р-450-оксигеназпая система, присутствующая в печени млекопитающих.
Недаром говорят о «барьерной» роли печени, которая является своеобразным фильтром, очищающим организм от вредных веществ. G помощью этой ферментной системы превращаются и тем самым обезвреживаются многие ядовитые для организма неполярные, а значит, нерастворимые в воде соединения — лекарственные вещества, наркотики и др. Задача этой системы — превращение нерастворимых соединений в растворимые в воде, с тем чтобы можно было вывести их из организма.
Цитохром Р-450 обнаружен у многих животных, растений и бактерий. Его нет у бактерий-анаэробов, живущих в бескислородных условиях.
А. И. Арчаков называет цитохром Р-450 «мембранным иммуноглобулином». Последний находится в мембранах эндоплазматического ретикулума. К 1980 г. было известно не менее 20 форм цитохрома Р-450. Множественность форм характерна именно для высших организмов, тогда как бактерии содержат лишь один тип цитохрома Р-450.
Существование множественных форм, вероятно, объясняет широкую субстратную специфичность оксигеназной системы, которая может окислять самые различные молекулы. Предполагается, что в ответ на введение в организм определенного класса ксенобиотиков синтезируется и определенная группа цитохрома Р-450, подобно тому как в ответ на введение макромолекулярного антигена возникают строго комплементарные к нему антитела.
Таким образом, в организме млекопитающих существуют две системы иммунологического надзора. Первая из них — лимфоидная система, уничтожающая клетки и высокомолекулярные соединения, вторая — монооксигеназная система, детоксицирующая ксенобиотики. Если первая иммунная система защищает организм от чужеродных макромолекул, то вторая — от чужеродных низкомолекулярных веществ. Предполагается, что иногда обе иммунологические системы действуют в совокупности. После окисления оксигеназной системой ксенобиотика его окисленная форма связывается с определенным белком. Образовавшийся коньюгат приобретает антигенные свойства и начинает вызывать образование антител. Роль коньюгазы опять-таки выполняет цитохром Р-450. Получается, что ксенобиотик, попадая в организм животного, индуцирует не только свое окисление, по и биосинтез соответствующих антител.
С помощью оксигеназной системы окисляются не только экзогенные ксенобиотики, по и ряд эндогенных (внутренних), образующихся в организме: стероидные гормоны, жирные кислоты, простагландины и др.
В печени млекопитающих существует еще одна система, помогающая им убирать из организма ксенобиотики. Это присоединение, или конъюгация, к различного рода лекарствам, ядам, наркотикам и другим соединениям глютатиона, в результате чего ксенобиотики обезвреживаются, а затем и выводятся из организма.
Однако в действии обезвреживающих систем бывают и осечки. Известны случаи, когда эти системы, стремясь обезвредить какое-нибудь токсическое вещество, превращают его в канцероген, т. е. в соединение, способное вызывать злокачественную опухоль.
Все, что сказано, относится к системам обезвреживания ксенобиотиков в организмах млекопитающих, где эти процессы усиленно исследовались и продолжают исследоваться, А как обстоит дело у растений? Вопрос далеко не праздный, поскольку именно растениям приходится в основном принимать на себя тот бесконечный поток чужеродных веществ, который сам человек и созданная им промышленность обрушивают на их поверхность. К сожалению, такие исследования если и проводились, то в крайне ограниченном количестве. А те сведения, которыми мы располагаем, в основном относятся к способности растительных тканей превращать гербициды (главным образом 2,4-дихлорфенолуксусную кислоту), а также некоторые инсектициды. Даже знаменитый ДДТ в этом отношении до сих пор остается почти неисследованным, более того, существует мнение, что растения не в состоянии ею метаболизировать.
Однако те ограниченные сведения, которые все же имеются в литературе, позволяют заключить, что и у растений имеются системы детоксикации ксенобиотиков, напоминающие по своим свойствам оксигеназную систему микросом печени млекопитающих. В составе растений, принадлежащих к 20 видам, обнаружен цитохром Р-450, спектральные характеристики которого удивительно похожи на спектры соответствующих цитохромов из печени млекопитающих. В микросомах более чем 20 видов растений обнаружено наличие оксигеназной активности, способной превращать ряд ксенобиотиков. Эта ферментная система зависит от наличия липидного кофактора и подавляется теми же ингибиторами, что и оксигеназы из микросом печени.
У растешгы присутствует также ряд ферментов, ответственных за присоединение к гербицидам глютатиона. Предполагают, что такой механизм обезвреживания может объяснить нечувствительность некоторых растений к гербицидам.
К числу эндогенных ксенобиотиков высших растений принадлежат и фитоалексины. Тот факт, что они являются чужеродными соединениями для растительных тканей, в сочетании со свойством фитотоксичиости диктует настоятельную необходимость их удаления из растений. И действительно, эти вещества, накопившись в тканях устойчивых растений в токсических для паразита концептрациях и тем самым уничтожив находящихся в них паразитов, затем исчезают из растительной ткани. И хотя роль оксигеназных цитохромсодержащих систем растений до сих пор никем не исследовалась, однако имеющаяся к настоящему времени, пусть даже и ограниченная, информация позволяет предполагать их участие в детоксикации фитоалексинов.
Получение прямых доказательств участия монооксигеназной системы в способности растений детоксицировать экзогенные и эндогенные ксенобиотики и тем самым поддерживать свой химический гомеостаз, нуждается в более пристальном внимании фитоиммунологов, чем то, которое до сих пор ему уделялось. Не исключено, что результаты этих исследований покажут, что растения на нашей планете функционируют не только как «зеленые легкие», образуя кислород в процессе фотосинтеза, но и как «зеленая печень», осуществляющая метаболизм ксенобиотиков и защищающая биосферу от загрязнения.
РАСПОЗНАТЬ,
ЧТОБЫ ПРИНЯТЬ ИЛИ УНИЧТОЖИТЬ
Распознавание «своего» и «чужого», иммунологический надзор, охрана биологической индивидуальности, иммунологическая цензура, полицейская функция иммунитета — все это синонимы, используемые различными учеными с одной целью: подчеркнуть главную задачу иммунитета — охрану генетического постоянства внутренней среды организма.
Р. В. Петров[16]
КТО ЕСТЬ КТО
Мы рассмотрели основные защитные реакции растений, от согласованного протекания которых зависит существование как проникшего паразита, так и растения-хозяина. Включатся вовремя защитные реакции — растение выживает, а паразит погибает, не включатся — все наоборот. Но от чего же зависит их включение? Ведь гибель паразита на устойчивом растении или его развитие на восприимчивом является уже конечным этапом, исходом болезни. А что же происходит на начальных стадиях, на которых определяется, будут или нет защитные реакции вызваны, или индуцированы? Недаром же иммунитет растений так же, как и животных, является не столько конституционным, сколько индуцированным свойством. Значит, должен существовать некий механизм, который срабатывает на первых этапах соприкосновения паразита и хозяина или взаимодействия их метаболитов. Этот механизм должен вызывать защитные реакции у устойчивого растения, но не вызывать их у восприимчивого.
Недаром Р. Толбойс еще в начале 50-х годов сумел увидеть во взаимодействии паразита и хозяина две последовательные стадии, которые назвал детерминантной и экспрессивной. На детерминантной или определяющей стадии (от английского to determinate — определять) и решается, будет или не будет защищаться растение. На второй экспрессивной (от английского to express — выражать) фазе происходит выражение того, что уже было ранее решено на детерминантной фазе.
До сих пор мы говорили только об экспрессивной фазе, хотя не она является первой и определяющей. Эта фаза болезни недаром названа экспрессивной: ее проявление легко заметить. Она выражается в пятнистости, загнивании, некрозе, хлорозе, увядании и других симптомах, свидетельствующих либо о течении болезни, либо о ее прекращении в результате подавления паразита механизмами защиты хозяина. Детерминантная фаза внешних симптомов не имеет. Эта последняя фаза до сих пор оставалась не только неизученной, но се существование подвергали сомнению.
Перемещение внимания фитоиммунологов с экспрессивной на детерминантную фазу, т. е. от самих защитных реакций к тем причинам, которые их вызывают, стало возможным лишь в последние 10–15 лет. Именно это новое направление исследований и обеспечило тот очевидный прогресс, который сейчас имеет место в фитоиммунологии. Причиной его возникновения явилось переосмысливание ранее принятых в иммунологии понятий, которое происходило в последние годы и суть которого может быть сформулирована следующим образом: главная функция иммунитета состоит в распознавании чужеродной генетической информации.
Д. Уилсон в своей книге «Тело и антитело» определил иммунологию как науку о «самости». По его образному выражению, главная миссия иммунитета состоит в способности организма отличать свое «Я» от не своего «я». Любая клетка, любая ткань, любой белок должны быть уничтожены, если они не являются своими. Иммунная система признает только свое.
Иммунная система, роль которой в самом общем виде можно определить как распознавание и элиминацию чужеродных веществ, клеток и тканей, возникла на самых ранних этапах эволюции. История эволюции иммунного ответа насчитывает 400 млн. лет и восходит к палеозою. По образному выражению американского эволюциониста Э. Купера, «иммунная система так же стара, как самое старое беспозвоночное, и столь же юна, как человек»
[17].
Все организмы, начиная с амебы, обладают способностью к фагоцитозу — внутриклеточному перевариванию. У одноклеточных таким путем одновременно осуществляются процессы как питания, так и защиты, тогда как у многоклеточных, начиная с губок, функцию защиты взяли на себя особые клетки иммунной системы, называемые фагоцитами.
На более высокой ступени эволюции к клеточному иммунитету добавляется гумморальный. Клеточным и гумморальным иммунитетами обладают только позвоночные — рыбы, амфибии, рептилии, птицы и млекопитающие. Способность к образованию антител впервые появляется у наиболее примитивных позвоночных — круглоротых рыб (миног и миксин). Однако у них присутствует лишь единственный вид антител — иммуноглобулины М, тогда как у самых совершенных позвоночных, в том числе и у человека, имеется пять классов иммуноглобулинов: М, G, Е, A, D.
Хотя иммунные системы организмов, находящихся на различных ступенях эволюции, различаются, всех их роднит общее свойство — способность распознавать чужеродность. Таким свойством обладает все живое независимо от степени его эволюционного совершенства. Наличие антител при этом совсем необязательно. Купер утверждает, что все компоненты, иммуногенные для позвоночных, являются иммуногенными и для беспозвоночных. И те и другие умеют распознавать чужеродность и обладают способностью ее устранять. Вернее так: все они перед тем, как ликвидировать чужое, должны его распознать.
С этих позиций иммунитет растений следует рассматривать как систему, охраняющую их структурную и функциональную целостность, основанную на способности распознавать и отторгать генетически чужеродную информацию. Несмотря на то что растения в силу присущих им особенностей неспособны образовывать иммунокомпетентные лимфоциты и антитела, им, как и всему живому, присуща способность распознавать чужеродные ткани, клетки и вещества.
Распознавание — это функция клеточной поверхности, в обязанности которой входит первой встретить внешний агент, правильно его расцепить и адекватно на него отреагировать. Клеточная поверхность прежде всего должна выяснить, кто есть кто. Кто приближается к ней: друг или враг, свой или чужой. Если чужой — его нужно задержать и уничтожить, если свой — пропустить и принять. Любой непрошенный чужеродный гость, будь это ткань, клетка или вещество, должен быть немедленно выдворен.
Но для того чтобы уничтожить, нужно предварительно распознать. Это делают специальные поверхностные рецепторы — своеобразные органы чувств клетки. Поверхность каждой клетки представляет собой мозаику из таких рецепторов. Рецепторы, в большинстве своем имеющие углеводную природу (но встречаются и белки), представляют собой специализированные участки, которые могут избирательно связываться с определенными молекулами, находящимися либо в свободном состоянии, либо на поверхности других клеток. С одними молекулами рецепторы реагируют, другие — игнорируют. Таким способом поверхность клеток отделяет чужое от своего. И обо всех событиях, происходящих на границе, поверхность клетки быстро передает в центр, где и принимается соответствующее решение — принять либо отторгнуть.
Самым удивительным примером способности клеток к распознаванию «своего» является оплодотворение у морских животных, у которых большое число яйцеклеток и миллионы сперматозоидов выделяются прямо в воду. В этом случае вероятность встречи яйцеклетки, к примеру, морского ежа с его же сперматозоидом, а не сперматозоидом устрицы, омара и других представителей морской фауны очень невелика. Это оказывается возможным только благодаря способности яйцеклетки «выбирать» сперматозоид своего вида из множества присутствующих во внешней среде. И это, в свою очередь, определяется наличием на поверхности яйцеклетки специфических рецепторов, которые узнают соответствующие вещества на поверхности сперматозоидов. В результате последние прикрепляются (иммобилизируются) к поверхности яйцеклетки, активизируются и проникают внутрь ее. Сперматозоиды других видов яйцеклеткой не распознаются и поэтому не оплодотворяют ее.
Хорошо известно, что бактериофаги (вирусы, поражающие бактерии) определенного штамма прикрепляются только к определенным видам бактериальных клеток, которые они поражают.
Процесс распознавания лежит и в основе питания некоторых простейших, к поверхности которых прикрепляются только те бактерии, которые могут служить им источником пищи.
Очень интересна сортировка клеток в процессе дифференцировки и органогенеза, которая также происходит на основании способности узнавать. Оказалось, что те или иные клетки эмбриона мигрируют в определенные области тела, где распознают друг друга, слипаются и таким образом формируют нужный орган. Если суспензии клеток различных органов равномерно распределить в объеме, то через некоторое время они начнут мигрировать, а затем агрегируют, образуя зачатки того органа, которому они и принадлежат. Таким образом, клетки способны узнавать либо отвергать друг друга.
Известно, что иммунокомпетентные клетки млекопитающих распознают антигены с помощью рецепторов на клеточной поверхности, что необходимо для реализации иммунного ответа. Проблема специфических рецепторов на таких иммунокомпетентных клетках непосредственно касается проблемы узнавания генетически чужеродных субстанций.
Сущность рецепторов В-клеток достаточно изучена. Эти рецепторы или точки распознавания на лимфоцитах в действительности представляют собой антитела иммуноглобулиновой природы.
Предполагается, что часть иммуноглобулиновых рецепторов свободно выделяется наружу, а часть остается прикрепленной к мембране лимфоцита. Рецепторы синтезируюся постоянно и отделяются от клеточной поверхности лимфоцита. На месте отделенных рецепторов тут же возникают новые.
Что касается рецепторов у Т-клеток, то здесь пока еще не все ясно. Но во всяком случае процесс взаимодействия Т-клеток, или клеток киллеров (убийц), с клеткой-мишенью удалось заснять на кинопленку. При этом видно, как иммунные Т-клетки окружают клетку-мишень, прикрепляются с помощью распознающих рецепторов к ее поверхности и вытягивают по направлению к своей мишени отростки. Как только такой отросток касается поверхности клетки-мишени, последняя погибает, а вместе с ней погибают и нападающие клетки. Иммунологи называют этот феномен «поцелуем смерти».
Явление распознавания определяет также первые этапы совместимости либо несовместимости антигенов пыльцы и пестика, при опылении растений, совместимость трансплантата и растения-реципиента при прививках, симбиотические отношения у растений и грибов микоризообразователей, а также растений семейства бобовых и бактерий-азотофиксаторов. В этом случае, если антигены, расположенные на поверхности взаимодействующих тканей, окажутся совместимыми, отторжения не произойдет и результаты опыления, прививок, симбиоза окажутся положительными. Если, напротив, антигены окажутся несовместимыми, произойдет отторжение.
Распознавание пестиком пыльцы представляет собой наиболее четкий пример способности растительных тканей отличать свое от чужого. Показано, что определяющая роль в этом процессе принадлежит поверхностным белкам гликопротеидной природы, так называемым S-белкам, которые являются продуктами S-генов.
На рыльце того или иного растения обычно попадает пыльца многих растительных видов, однако прорастать через столбик может только пыльца своего вида. Рыльце как бы узнает свою пыльцу. При этом из особых полостей, имеющихся в стенках пыльцевых зерен, выделяются на поверхность рыльца определенные вещества, которые связываются с поверхностью рыльца, покрытого липкими выделениями. Такое связывание специфических факторов узнавания и определяет успех опыления.
Теперь что касается прививок. Нужно сказать, что трудности при трансплантации у животных много серьезнее, чем при прививках у растений. Если у млекопитающих и человека иммунная система распознает и отторгает ткани, принадлежащие другому индивидууму того же вида (пересадка органов возможна только между однояйцевыми близнецами), то у растений внутривидовые прививки являются обычным делом, тогда как известны случаи межродовых и межсемейственных прививок. Таким образом, хотя «самость» у растений и существует, но она выражена значительно менее категорично, чем у млекопитающих.
Однако в этой, казалось бы, четкой закономерности при прививках растений существует много исключений. Известны примеры, когда растения даже внутри вида не прививаются. Если совместимость при прививках тем успешнее, чем ближе таксономическая близость партнеров, то почему же тогда орех трудно прививается сам на себя, тогда как хурма и каштан — растения, принадлежащие к различным ботаническим семействам, — прививаются по высшему баллу? Почему вишня хорошо прививается на черешню, а черешня на вишню не прививается или прививается плохо? Почему на айве прививаются только некоторые сорта груш? Эти и многие другие вопросы пока остаются без ответа.
В случае несовместимости привоя и подвоя между ними возникает некроз и разыгрывается реакция, внешне напоминающая реакцию СВЧ. Очевидно, на поверхности соприкосновения привоя и подвоя происходит взаимное распознавание, вероятно определяемое, как и у трансплантатов животных, наличием антигенов гистосовместимости.
Особо важная роль принадлежит прививкам в виноградарстве. До появления в Европе филлоксеры прививки здесь применялись только в ограниченных размерах или, во всяком случае, не имели для виноградарства того значения, которое они имеют сейчас в районах, зараженных или находящихся под угрозой заражения филлоксерой. Дело в том, что корневая система американских или франко-американских лоз мало страдает от нападения на них филлоксеры, поэтому единственным средством борьбы с филлоксерой в Европе является прививка европейских лоз на эти подвои.
Для массового размножения виноградной лозы, как правило, принята прививка однолетних черенков в комнатных условиях, или, как это обычно называют, прививка на столе. Заготавливаются привойные и подвойные черенки, которые соединяются по свежим срезам, затем стратифицируются и высаживаются в «школку». Необходимым условием сращивания между привоем и подвоем является образование у обоих компонентов особой ткани, так называемого каллюса.
Между соприкасающимися поверхностями привоя и подвоя образуется изолирующая прослойка, состоящая из отдельных участков некротизированной ткани (реже — сплошного слоя), которая в случае совместимости привоя и подвоя постепенно рассасывается. Чем совершеннее прививка, тем меньше некротизированных участков между тканями подвоя и привоя.
В свете всех изложенных представлений исход соприкосновения растительной клетки с клеткой фнтопатогена будет зависеть от взаимодействия поверхностных макромолекул, синтезируемых одним организмом, с макромолекулами на поверхности второго.
Соприкосновение возникает при взаимодействии поверхностей двух тел, сфер или фаз, в случае паразитизма — это поверхности соприкосновения взаимодействующих макро- и микроорганизма.
Представьте себе, что фитопатоген проникает в клетку растения. Происходит соприкосновение их поверхностей, в результате чего возникает так называемая поверхность взаимодействия двух организмов. Рецепторы клетки хозяина проверяют поверхность клетки паразита, ищут среди них чужеродные антигены. Если такие антигены найдены, рецепторы растительной клетки хозяина связываются с поверхностными антигенами паразита. И вот уже в центр клетки посылается приказ: «Включить защитные реакции!».
К настоящему времени описано не менее 40 типов поверхностей взаимодействия. Поверхность взаимодействия между клетками хозяина и паразита будет различна в зависимости от того, проникает ли паразит внутрь клетки или развивается в межклетниках. Например, большинство фитопатогенных бактерий родов Pseudomonas и Xanthomonas обитают не в клетках растений, а в пространстве между клетками, поэтому между протопластами хозяина и паразита находится клеточная стенка растения и оболочка бактерии. Гифы возбудителя фитофтороза картофеля проникают в его клетки, и поэтому поверхность их взаимодействия представляет собой соприкосновение цитоплазматической мембраны хозяина с клеточной стенкой паразита. Иногда протопласт паразита непосредственно соприкасается с протопластом хозяина, как это происходит при заражении растений семейства крестоцветных плазмодиальными паразитами, и тогда взаимное распознавание предполагает взаимодействие молекул на поверхности цитоплазматических мембран партнеров. С некротрофами дело обстоит несколько иначе, поскольку некротрофные паразиты не вступают в контакт с живыми клетками растения, а соприкасаются с уже убитыми ими с помощью токсинов. Возможно, что в этом случае следует принимать во внимание взаимодействие токсина с поверхностью еще живых клеток, от чего зависит, останутся ли они живыми или погибнут.
По-видимому, специфичность взаимодействия бобовых растений с клубеньковыми бактериями рода Rlnzobium также основана на взаимном распознавании макро- и микробионта. В этом процессе, как уже ранее говорилось, большая роль принадлежит лектинам, которые связывают бактерию, специфически приуроченную образовывать клубеньки именно на этом виде растения.
ИНДУКТОРЫ, ЭЛИСИТЕРЫ, ПРОВОКАТОРЫ
Что это такое? Это — вещества паразита, которые распознаются растением и служат тем самым для него сигналом к включению системы ответных защитных реакций. Вначале их назвали индукторами, поскольку они индуцируют устойчивость. Однако вследствие широкого толкования слова индуктор (индуцировать можно многие процессы, а не только устойчивость), термин индуктор предложили заменить на элиситер (от английского to elicit — вызывать, извлекать). А затем, на 12-м Ботаническом конгрессе в Ленинграде, слово элиситер было переведено как «провокатор», что оказалось очень близким к истине, поскольку никакое слово лучше этого не отражает истинную роль элиситеров в составе фитопатогенов. Элиситеры как бы «выдают» растению присутствие паразитов. Они представляют собой нечто вроде пятой колонны среди веществ паразита.
Тогда не может не возникнуть следующий вопрос, а затем же патогенам иметь в своем составе провокаторы-элиситеры? Ведь не для того же, чтобы их присутствие выдавало паразитов растениям, в которые они проникают? Конечно, нет. Дело не в том, что паразиты содержат элиситеры, чтобы позволить растению себя распознать, а в том, что растение приобретает способность распознавать среди метаболитов патогена вещества, которые начинают играть роль элиситеров. Роль элиситеров им навязывается растением. У паразита они служат совсем другим целям, выполняя нужные патогену функции. Чаще всего они являются структурными элементами клеточных оболочек либо экстрацеллюлярными ферментами и т. п.
Какова природа элиситоров и где они локализованы? Обычно это либо поверхностные, либо выделяющиеся паразитом вещества, т. о. именно то соединения, которые первыми соприкасаются с поверхностью растения. Ими могут быть хитин, хитозан, глюканы, гликопротеины, липополисахариды, липогликопротеидьг, пектолитические и протеолитические ферменты. Как правило, большинство элиситеров содержат в своем составе углеводы, роль которых в процессах распознавания общеизвестна. Одпако бывают и исключения. Так, элиситер фитоалексина гороха — пизатина — мопиликолин А, выделенный из паразита, вызывающего плодовую гниль яблок и грунт (Monilia fructigena), представляет собой полипептид с молекулярной массой около 8000 Дальтон.
Кстати, мониликолин А был первым элиситером, выделенным в Австралии в лаборатории профессора И. Круикчанка. Сейчас их известно несколько десятков, хотя не все они одинаково хорошо охарактеризованы. Часто один и тот же патоген содержит не один, а два или даже три элиситора, разной химической природы и по-разному локализованных в составе патогенов.
Так, в клеточных стенках и в составе выделений возбудителя фитофтороза сои присутствуют два элиситера, один из которых β-1,3-β-1,6-глюкан с молекулярной массой 100 000, а другой гликопротеин с молекулярной массой 150 000 Дальтон. В клеточпых стенках возбудителя фитофтороза картофеля нами обнаружены высокомолекулярные β-1,3-β-1,6-глюканы, а в цитоплазме липогликопротеидный комплекс. На поверхности бактерий находится липополисахаридный комплекс, который также выполняет функцию элиситера. Элиситеры индуцируют фитоалексины в крайне низких концентрациях, составляющих 10
-9 и даже 10
-13 М.
К настоящему времени более или менее очищен только одип элиситер из клеточных стенок возбудителя фитофтороза сои. Оказалось, что он представлял собой олигосахарид, состоящий из нескольих глюкозпых остатков, связанных β-1,3 и β-1,6-связями. Однако даже в составе наиболее очищенной фракции элиситера присутствуют полимеры глюкозы, связанные в положении 1–4, которые рассматриваются как «загрязнение».
Очень мало известно относительно механизма действия элиситеров. Поскольку каждое ботаническое семейство имеет фитоалексины разной химической природы, за синтез которых ответственны разные ферментные системы, то изучать механизмы индукции очень сложно. Предполагается, что в здоровом растении группа генов, ответственных за образование ферментов, синтезирующих фитоалексины, зарепрессирована. Индукция связана с дерепрессией этих участков генома клетки, в результате чего и начинается синтез ферментов. В пользу подобного предположения говорит тот факт, что под влиянием заражения несовместимыми паразитами в растительных тканях начинается активный синтез молекул РНК и усиленный синтез белка. Если инфицированные ткани обработать метаболическими ядами, ингибирующими синтез РНК и белка, образование фитоалексинов прекращается.
Пожалуй, наиболее убедительным доказательством в пользу справедливости этого предположения являются опыты с фитоалексином касторовых бобов — касбеном, который по химической природе представляет собой дитерпен. В тканях устойчивых бобов под влиянием заражения синтез касбена возрастал в 20–40 раз. Но самое интересное состояло в том, что заражение активировало не синтез дитерпенов вообще, а только одного дитерпена — касбена. Синтез других дитерпенов под влиянием заражения оставался в пределах нормы. Это говорит о том, что синтез касбена в результате инфицирования является результатом специфической индукции соответствующих ферментов, а не результатом усиления биогенеза дитерпенов вообще.
Существует и другая гипотеза, согласно которой ферменты, ответственные за образование фитоалексинов, хотя и существуют в здоровой клетке, по находятся в ней в неактивном состоянии, поскольку связаны некими ингибиторами. Под действием элиситера ингибирование снимается.
Большинство известных к настоящему времени элиситеров обнаружено на основании их способности вызывать накопление в растительных тканях фитоалексинов. Впоследствии стало ясным, что многие из элиситеров не только индуцируют синтез этих веществ, но и вызывают некротизацию тканей, являющуюся неотъемлемой составной частью реакции СВЧ, а также образованно раневой перидермы, отторгающей зону инфицирования от здоровой ткани. Таким образом, многие из числа ныне известных элиситеров индуцируют не только фитоалексины, по вызывают образование целого комплекса защитных реакций, связанных с реакцией СВЧ, и поэтому их правильнее называть элиситерами защитных реакций.
Еще Мюллер определил фитоалексины как продукты некротизации клеток. И действительно, подавляющее большинство элиситеров вызывают некроз клеток и накопление в них фитоалексинов. Однако есть и исключения, которым является мониликолин А. Он вызывает накопление фазеолина в клетках фасоли, которые остаются живыми, окрашиваются прижизненными красителями и сохраняют способность к плазмолизу (способность к плазмолизу и окраска прижизненными красителями — признак жизнеспособности клеток).
Индуцировать образование фитоалексинов могут не только паразитарные грибы, но и бактерии, вирусы, нематоды, а также некоторые насекомые. Кроме того, их образование может индуцироваться под влиянием ряда химических агентов и физических воздействий. Все существующие элиситеры были разделены на два класса: биотические (имеющиеся у микроорганизмов) и абиотические. К числу абиотических элиситеров относятся соединения, ничего общего не имеющие с продуктами метаболизма фитопатогенов. Сюда относятся соли тяжелых металлов, которые по своей способности индуцировать, например, образование пизатина располагаются в следующий ряд:
Hg++>Ag+>Cu++>Ni++>Fe+++>Cd+++>Zn++> >Со++>Мп++.
К числу абиотических элиситеров относятся также некоторые ингибиторы отдельных метаболитических процессов (иодацетат, фтористый натрий, 2,4-динитрофенол, некоторые антибиотики, фенольные соединения, цитокинины, циклический аденозинмонофосфат и, наконец, многие пестициды). Элиситерами являются и некоторые физические воздействия, например облучение ультрафиолетовыми лучами, частичное замораживание. Сообщалось даже, что умеренное постукивание по листьям конских бобов вызвало у них образование фитоалексинов, тогда как сильный удар — нет.
По-видимому, нет общей структуры вещества, которое определило бы его принадлежность к злиситерам, так же как и нет какой-либо определенной мишени для их действия у растительных клеток. В качестве элиситеров могут выступать вещества, различным образом повреждающие растительные клетки.
Если сравнить действие абиотических элиситеров с биотическими, то налицо их отличие, ио крайней мере по двум пунктам.
1. Биотические элиситеры вызывают образование фитоалексинов в значительно больших концентрациях, чем абиотические. Так, абиотические элиситеры оказываются активными в концентрациях 10
-3—10
-5 М, тогда как биотические — 10
-9 М и ниже, как мы это уже говорили. Причем даже в максимально индуцирующих концентрациях абиотические элиситеры редко могут вызвать образование фитоалексинов в концентрациях, приводящих к полному подавлению роста паразитов.
2. Под влиянием абиотических элиситеров в растительных тканях индуцируются не только фитоалексины, но и ряд других соединений, не имеющих никакого отношения к реакции несовместимости паразита и хозяина. Наоборот, биотические элиситеры вызывают прицельное индуцирование только фитоалексинов.
Недавно удалось выявить разницу в действии биотических и абиотических элиситеров. Оказалось, что накопление фитоалексинов под действием биотических элиситеров происходит за счет их интенсивного синтеза, который значительно преобладает над процессом их распада — метаболизма. Абиотические элиситеры действуют наоборот. Иными словами, многие абиотические агенты, повреждающие клетку, подавляют процесс метаболизма ксенобиотиков — фитоалексинов, благодаря чему последние и накапливаются. Более того, в силу повреждения систем метаболизма они могут сохраняться в этих тканях в течение определенного времени, тогда как под действием биотических элиситеров фитоалексины после достижения фунгитоксических концентраций исчезают из растительных тканей, поскольку элиситер не повреждает системы их метаболизма.
В свете этих данных по-новому можно представить себе, как отражается на содержании фитоалексинов обработка растения теми пестицидами, которые обладают способностью вызывать их образование. Под воздействием таких пестицидов индуцируется накопление в растительных тканях фитоалексинов, которые сохраняются в ткани, а не исчезают из нее, поскольку процессы их метаболизма подавлены. Обработка пестицидами может происходить несколько раз в сезон, поэтому количество фитоалексинов в растительных тканях все более и более возрастает и может накопиться в них в токсических для человека и животных концентрациях. Конечно, совсем не обязательно, чтобы все пестициды служили элиситерами фитоалексинов и при этом ингибировали их метаболизм. Число таких элиситеров достаточно велико, и трудно представить себе, чтобы все они имели единый механизм действия. Однако при испытании и отборе новых пестицидов следует все же иметь в виду их нежелательную способность вызывать накопление фитоалексинов в растительных тканях.
Итак, предполагается, что на поверхности паразитарных микроорганизмов имеются определенные вещества, которые распознаются растением и являются для него элиситерами защитных реакций, чаще всего реакции СВЧ. Казалось бы, все просто и ясно. Элиситеры вызывают СВЧ-гибель клеток хозяина и образование фитоалексинов, которые накапливаются в отмерших клетках и губят присутствующего там паразита. Однако в этой простой схеме не все концы сходятся с концами.
1. Элиситер паразита, нанесенный на поверхность растительной ткани, вызывает гибель поверхностных клеток, с которыми он непосредственно соприкасается, тогда как фитоалексины образуются в более глубокорасположенных слоях клеток, куда элиситер вряд ли проникает.
2. СВЧ-гибель клеток растения наступает вскоре после соприкосновения с элиситером паразита. Так, клетка листа или черешка картофеля погибает уже спустя 30 минут после того, как ее цитоплазматическая мембрана приходит в соприкосновение с гифой несовместимой расы возбудителя фитофтороза, тогда как фитоалексины начинают образовываться лишь спустя несколько часов после гибели клеток. Более того, если с помощью определенных приемов задержать СВЧ-гибель клетки, то настолько же задерживается и начало образования фитоалексина, и, наоборот, если СВЧ-гибель ускоряется, то ускоряется и образование фитоалексина.
Создается впечатление, что ответная реакция растительной ткани включает в себя как минимум два последовательных этапа: элиситер вызывает СВЧ-гибель соприкасающихся с ним растительных клеток, а уже СВЧ-реакция служит как бы вторичным сигналом к образованию фитоалексинов. Можно было бы предполагать, что сигнал к их возникновению передает некое вещество, которое освобождается из погибших (или погибающих?) клеток растения-хозяина.
И такое вещество было недавно обнаружено в сое американским фитоиммунологом П. Альберсхеймом. Им оказался фрагмент клеточных стенок сон, состоящий из 12 молекул галактуроновой кислоты, соединенных между собой α-1,4-глюкозидными связями. Близкий по строению фрагмент удалось выделить из препарата цитрусового пектина.
Полученные данные позволили понять механизм индукции фитоалексинов с помощью пектолитических ферментов, которые воздействуют на клеточную стенку растения и высвобождают из нее пектиновый фрагмент, являющийся своеобразным химическим посыльным, или вторичным элиситером, вызывающим образование фитоалексинов в прилегающих здоровых клетках. По-видимому, в погибающих под действием элиситеров паразита растительных клетках также активизируются собственные ферментные системы, разрушающие клеточную стенку и высвобождающие индуцирующий фитоалексины фрагмент. А поскольку гибель или повреждение клеток вызывают многие химикаты и физические воздействия, то неудивительно, что в ответ на их обработку в растительных тканях образуются фитоалексины. Ведь недаром их количество часто прямо пропорционально степени токсичности элиситера для клетки растений.
Таким образом, в ответ на широкий набор воздействий, убивающих или повреждающих растительные клетки, из их клеточных оболочек высвобождается своеобразный химический курьер, или иммунологический посыльный, который диффундирует в прилегающие клетки, где либо индуцирует образование защитных веществ, либо повышает способность к их образованию.
Таким образом, вновь полученные данные позволяют рассматривать в более широком аспекте вещества, защищающие растения от инфекции. Они позволяют сохранить целостность растениям, поврежденным токсическими химикатами либо физическими воздействиями, поскольку страхуют их от инфекционных болезней. Иммунная система в таких растениях ослаблена, в силу чего поврежденные участки их тканей оказываются незащищенными, через которые инфекция
беспрепятственно проникает.
Особое значение приобретают неспецифические свойства фитоалексинов, в силу чего они будут предохранять поврежденные растения от широкого набора потенциально опасных для них патогенов. Ведь мало ли каким патогеном может поразиться поврежденный участок.
Если подходить к защитной роли фитоалексинов с таких позиций, то следует признать, что они в известной мере оправдывают название «стрессовые метаболиты», как их определяют некоторые исследователи. Другое дело, что совсем не все растительные метоболиты, образующиеся в ответ на стресс, обладают антибиотическими свойствами, и поэтому понятие о стрессовых метаболитах шире. Тем не менее, если раньше фитоалексины определяли как индуцированные антибиотические вещества, защищающие растения только от патогенов, то теперь, вероятно, правильнее говорить, что эти вещества защищают растительные ткани от целого ряда стрессовых воздействий, в том числе и от некоторых химических и физических повреждений.
Альберсхейм с соавторами предлагает новый термин «олигосахарины» для углеводов, обладающих биологической активностью. В свете таких представлений к олигосахаринам относятся биогенные элиситеры, например структурные полисахариды клеточных стенок паразитарных грибов, которые способны распознаваться растениями и индуцировать у них защитные реакции. К этой же группе биологически активных углеводов принадлежат и эндогенные иммунологические посыльные пектиновой природы, высвобождаемые в ответ на стресс из состава клеточных стенок растений. Кстати, обнаружено, что такого рода посыльные не только вызывают образование фитоалексинов в соседних растительных клетках, но и индуцируют у них образование ингибиторов протеаз, инактивирующих ферменты патогенных микроорганизмов и насекомых, о чем уже шла речь в начале книги.
Возможно, что продукты деградации клеточных стенок действуют как гормоны, которые связываются со специфическими белками на плазмалемме, активизируя тем самым аденилатциклазу, синтезирующую циклическую АМФ. А в животной клетке циклическая АМФ является универсальным посредником при передаче гормонального сигнала.
Таким образом, полимеры клеточных стенок являются как бы хранителями регуляторных молекул, которые Альберсхейм по аналогии с животными считает возможным причислить к числу вторичных гормонов. Возможно, что вторичные растительные гормоны высвобождаются либо из стенок, либо других органелл растительных клеток не только в условиях стресса, но и под действием первичных гормонов, таких, как ауксины, гиббереллины, цитокинины и этилен. Таким образом, благодаря открытию Альберсхейма выступили из мрака контуры еще одного этапа в той цепи последовательных явлений, которые возникают при взаимодействии паразита с несовместимым для него хозяином.
Итак, клетка несовместимого хозяина, распознавшая элиситер проникающего паразита, гибнет в результате реакции СВЧ. Из ее клеточных стенок высвобождается олигосахарин, несущий весть об опасности в глубь ткани, заставляющий ее мобилизоваться для борьбы с врагом (одним словом, иммунологический буревестник). Налицо своеобразная эстафета индуцирования (рис. 10). Гибнущая клетка передает своим здоровым собратьям сигнал «SOS».
Значит, налицо, по крайней мере, два индуктора, один из которых является причиной СВЧ-гибели клетки, а другой — образования фитоалексинов. Действия этих индукторов разъединены во времени и в пространстве. Возможно, существует и третий индуктор, вызывающий образование раневой перидермы, отграничивающей область инфекции (см. рис. 10).
 Рис. 10. Иммунологические курьеры
Рис. 10. Иммунологические курьеры
В свете этих фактов несколько по-новому может быть интерпретировано одно из наиболее загадочных и до сих пор ускользающее от понимания исследователей явление СВЧ. Оказывается, жертва частью клеток во имя сохранения целого не так уж неоправданна и несовершенна. Именно умирающая клетка посылает в пространство сигнал об опасности. А это означает, что чем быстрее отомрет клетка, тем раньше узнают ее здоровые собратья о грозящей им опасности. А мы помним, что чем быстрее происходит СВЧ-гибель клетки, тем устойчивее сорт растения.
Некротизированная клетка — это отмершая клетка. Но под влиянием токсинов клетки хозяина тоже погибают. Образуется ли в них иммунологический посыльный, сигнализирующий о необходимости образования фитоалексинов, пока неизвестно. Возможно, что и нет.
Извещенные об опасности клетки срочно мобилизуют свои резервы, перестраивая обмен, синтезируют фитоалексины, которые направляются навстречу врагу, находящемуся в некротизированных клетках, где и накапливаются до концентраций, вызывающих гибель паразита. Почему они накапливаются именно в некротизированных клетках, нетрудно объяснить. Ведь для здоровых растительных клеток фитоалексины являются ядом (ксенобиотиками эндогенного происхождения), поэтому они подлежат немедленному уничтожению (метаболизму) с помощью уже упомянутой системы монооксигеназ. В некротизированных клетках эта система вряд ли в состоянии функционировать.
Получается, что некротизированные клетки, даже мертвые, продолжают верно служить своему растению, выполняя функции вместилищ яда. Иного выхода пет, поскольку то количество яда, которое нужно накопить, чтобы погубить паразита, пи одна живая клетка не в состоянии выдержать. Ведь яд настолько неизбирателен, что убивает подряд все живое, в том числе и собственные клетки, которые его производят.
Совершенно иное дело — иммунная система человека и животных, которая может позволить себе выбрасывать в кровяное русло антитела, нисколько при этом не опасаясь за жизнь клеток собственного тела. Антитела настолько специфичны, что будут уничтожать только свою мишень — антигены паразита, и не затронут собственных клеток.
Выходит, у растений нет и не может быть другого способа погубить паразита с помощью токсических фитоалексинов, кроме как накопить их в отмерших клетках. Возможно, что сигнал «SOS» можно было бы послать и минуя гибель собственных клеток, по концентрировать нужное для гибели паразита количество фитоалексинов можно только в отмерших клетках.
По-видимому, отмершая клетка не только посылает приказ живым клеткам синтезировать фитоалексины, но и каким-то образом руководит их направленным транспортом, являясь притягательным центром для их перемещения. Так ли это, покажет будущее.
ЗАГАДКА СПЕЦИФИЧНОСТИ
Нет ничего специфичнее иммунологического узнавания. Биологам это хорошо известно. Как же в таком случае микроорганизм может паразитировать, если на его поверхности находятся элиситеры, распознаваемые растением-хозяином? Значит, для того чтобы существовать, паразит должен суметь обмануть распознающие системы растения-хозяина. К таким «уловкам» паразит, действительно, прибегает, и их по меньшей мере может быть несколько. Попробуем рассказать о некоторых из них.
Первая возможность. Поскольку элиситеры, находящиеся на поверхности взаимодействия паразита и хозяина, позволяют последнему распознавать и отторгать паразита, то патогену проще всего было бы совсем убрать элиситеры из своего состава. Однако мы уже говорили, что те вещества паразита, которые растения превратили в своих провокаторов, выполняют у патогена определенные функции. Поэтому от провокаторов, хотя и нужно, но сложно избавиться, поскольку паразиту без них не обойтись. К тому же, если патоген ликвидирует один провокатор, то растение может приобретать способность распознавать другой, а затем и третий и т. д. Поэтому маловероятно, чтобы подобный путь «утери» элиситеров был генеральным способом избежать распознавания. Хотя кое-какие экспериментальные данные свидетельствуют в его пользу.
Это прежде всего явление большей выживаемости менее вирулентных рас по сравнению с более вирулентными, получившее название «стабилизирующий отбор». Такой отбор обеспечивает устойчивость данного вида или популяции к изменениям. Предположим, в популяции паразита появляется раса с генами вирулентности, позволяющими ей преодолеть комплементарные гены устойчивости хозяина. Это ставит ее в положение вне конкуренции с другими расами, поскольку только она способна поражать устойчивую форму растения, которую другие расы поражать не в состоянии. Но это же делает вирулентную расу менее жизнеспособной и конкурентоспособной в сапрофитной фазе или на восприимчивых сортах растений. На этой фазе простые расы, не имеющие генов вирулентности или имеющие их в меньшем количестве, будут вытеснять высоковирулентную расу. Излишняя вирулентность понижает способность к выживанию. Значит, раса приобретает новые гены вирулентности за счет неких потерь, которые сделали ее менее жизнеспособной. Поистине, любые блага приобретаются ценой потерь. Возможно, что потерей в данном случае и была утеря метаболита, выполняющего роль провокатора, хотя, так ли это в действительности, пока еще сказать трудно.
Вторая возможность избежать распознавания состоит не в утере паразитом элиситеров, а в их маскировке. Эяиситеры присутствуют, но только не на поверхности патогена, чтобы не быть распознанными растением. А что же находится на поверхности? А на поверхности паразита в целом ряде случаев могут располагаться вещества, которые имеются у самого растения. Да, да, мы не оговорились, на поверхности паразита располагаются вещества, имеющиеся у растения, или вещества, весьма похожие на вещества растения. Естественно, что растение не будет принимать за чужеродные свои собственные вещества. Оно как бы находится в заблуждении, клетки чувствуют себя неповрежденными, и ничто не оповещает их о грозящей опасности. Чем не тактика троянского копя? Вспомните, как греки длительное время безуспешно осаждали Трою и, наконец, оставили у ворот города сделанного ими коня, внутри которого находились вооруженные воины. Любопытные горожане втащили коня в городские ворота. Ночью из коня вышли воины, которые открыли ворота войскам противника. Примерно так же действует и паразит, и его тактика носит название «молекулярная мимикрия» (рис. 11, см. вклейку).
Мимикрия — маскировка. Помните бабочку, маскирующуюся под листок; червя, прикидывающегося сучком; безобидную муху угрожающей окраски. Явление молекулярной мимикрии в фитоиммунологии было впервые установлено Т. И. Федотовой, а затем подтверждено Н. Н. Гусевой и Б. Б. Громовой. Было замечено, что паразит и его растение-хозяин имеют общие вещества — антигены. И чем больше общих антигенов имеет паразит с растением, тем больше вероятность поражения растения данным паразитом.
Предполагается, что на поверхности паразита, взаимодействующего с растением, находятся хорошо знакомые растению антигены, тогда как индукторы паразита спрятаны где-то глубоко под ними. Паразит не распознается и беспрепятственно проникает в растение.
Третья возможность предотвратить распознавание паразита растением состоит в таком изменении молекулы элиситера, в результате чего она становится неузнаваемой для растения. Предполагается, что элиситеры, находящиеся на поверхности растения, являются как бы антигенной детерминантой для их распознавания рецепторными белками хозяина. При взаимодействии их продуктов включается реакция СВЧ и происходит отторжение паразита. Предположим, что у совместимого паразита или его расы происходит какое-то изменение молекул элиситера, в результате чего они перестают быть комплементарными рецепторным участкам растения и ими не распознаются. Такие элиситеры будут вызывать защитные реакции у устойчивых сортов растений и не будут вызывать их у восприимчивых, т. е будут обладать специфичностью действия. Поэтому пх и называли специфическими.
К сожалению, о специфических элиситерах пока еще мало известно. Но все-таки кое-какие данные на этот счет уже есть. Например, специфический элиситер был обнаружен в клеточных стенках и выделениях возбудителя фитофтороза сои. Элиситер оказался гликопротеином, состоящим на 70–90 % из белка и примерно на 10 % из углеводов, которые, по-видимому, и ответственны за его активность.
Поверхностными гликопротеинами оказались специфические элиситеры, выделенные из возбудителей антракноза фасоли и бактериоза сои.
Предполагается, что липополисахарид наружной мембраны бактерий обладает свойствами специфического элиситера. Мы уже писали, что сапрофитные, либо убитые нагреванием, либо, наконец, гетерологичные (несовместимые) для данного вида растений, бактерии иммобилизуются в их межклеточном пространстве. При этом бактерии как бы окутываются гранулярным и фибриллярным материалом, транспортирующимся везикулами из растительных клеток, и с помощью этого материала прикрепляются к клеточной стенке растения. В результате прикрепления наступает тесный контакт растительных клеток с липополисахаридом, который и распознается. Установлено, что его состав коррелирует со способностью бактерии, из которой он выделен, индуцировать реакцию СВЧ. Так, липополисахарид штаммов, которые индуцировали СВЧ, отличался от такового у неиндуцирующих штаммов молекулярной массой и отношением ксилозы и рамнозы к глюкозе.
Однако число элиситеров, обладающих специфическими свойствами, пока еще крайне ограниченно. Возможно, это зависит от того, что обнаружению специфических элиситеров мешают неспецифические, которые наряду со специфическими присутствуют у паразитов. Возможно, искать специфические элиситеры у фитопатогенов в искусственной культуре, как это делают исследователи, бесполезно, поскольку при этих условиях не могут проявиться их паразитические свойства.
Итак, наличие специфических элиситеров пока еще находится под сомнением, тогда как большинство известных к настоящему времени элиситеров обладает неспецифическими свойствами. Это означает, что они присутствуют у всех рас фитопатогенов, независимо от наличия у них генов вирулентности, и с их помощью можно индуцировать защитные реакции у всех сортов растений, независимо от присутствия у них генов устойчивости.
Маловероятно, чтобы растения обладали отдельными распознающими системами на каждый вид и расу фитопатогенов, с которыми им приходится сталкиваться в природе. Скорее всего, в роли неспецифических элиситеров могут служить структурные полисахариды клеточных стенок микроорганизмов.
Как же проникают в растения те фитопатогены, которые обладают незамаскированными неспецифическими элиситерами? Здесь мы расскажем о четвертой возможности, которая состоит в наличии у паразитов антиметаболитов, перекрывающих действие элиситеров. Речь идет о присутствии у некоторых паразитов антиэлиситеров, или супрессоров, которые в противовес элиситерам не индуцируют, а, наоборот, подавляют защитные реакции растений.
Супрессоры характерны не только для иммунных систем растений, но и животных. В начале 70-х годов была открыта особая группа лимфоцитов — супрессоров, оказывающая подавляющее действие на иммунные лимфоциты.
По-видимому, для каждой системы живого организма необходима специфическая антисистема, которая регулирует эту систему по принципу обратной связи.
В растительном мире супрессоры-антииндукторы были открыты через несколько лет после обнаружения индукторов-элиситеров. Предпосылкой к созданию таких представлений послужил давно установленный факт, что растение, зараженное совместимой расой того или иного патогена, становится восприимчивым не только к авирулентной расе того же патогена, но и к непатогенам, т. е. микроорганизмам, которые ранее не были способны поражать данный вид растения. Так, предварительное заражение ячменя вирулентной расой мучнистой росы сделало его восприимчивым не только к авирулентной расе того же паразита, но даже к возбудителю мучнистой росы дыни. Заражение картофеля совместимой расой возбудителя фитофтороза делало его восприимчивым к целому ряду микроорганизмов, обычно не поражающих неповрежденный картофель, даже к сапрофитам. Недаром существует выражение «вторичные инфекции», т. е. инфекции, возникающие на уже инфицированном растении.
Возможно, одной из причин вторичных инфекций являются супрессоры, подавляющие защитные реакции растения. По-видимому, супрессоры фитопатогенных микроорганизмов можно разделить на две группы: одни убивают или повреждают растительные клетки, тогда как другие только блокируют, задерживают защитные реакции. Если исходить из такой классификации, то к первой группе супрессоров следует отнести токсины паразитов. Естественно, что растительная клетка, убитая токсинами паразита, лишается способности распознавать патоген, а тем более отвечать защитными реакциями.
Ко второй группе супрессоров относятся импедины (от английского to impede — препятствовать, задерживать), название которых заимствовано из медицины, где он означает нетоксический бактериальный фактор, подавляющий защитный механизм хозяина. По-видимому, подобное разделение супрессоров на токсины и импедипы отражает разделение патогенов на некротрофы и биотрофы. Некротрофы продуцируют токсины, убивающие клетку на некотором расстоянии от самого паразита, биотрофы — супрессоры типа импединов, которые выделяются патогеном при непосредственном взаимодействии с растительной клеткой. О токсинах фитопатогенов мы уже говорили. А вот историю изучения импединов у растений интересно рассмотреть на примере открытия специфического супрессора у возбудителя фитофтороза картофеля.
В течение двух последних десятилетий исследователи, в том числе и авторы этой книги, упорно искали причину того, почему одна раса паразита поражает данный сорт картофеля, тогда как к другой авирулентной расе этот же сорт оказывается устойчивым (рис. 12). Обе расы морфологически не отличаются друг от друга, обе содержат в своем составе неспецифические индукторы защитных реакций, обе в равной мере чувствительны к токсическому действию фитоалексинов. Так в чем же, собственно, дело?
 Рис. 12. Поражение фитофторозом сорта картофеля с генами фитофтороустойчивости R1 и R4
Рис. 12. Поражение фитофторозом сорта картофеля с генами фитофтороустойчивости R1 и R4
И тогда ученые вспомнили старые опыты, согласно которым заражение картофеля вирулентной расой возбудителя фитофтороза лишало его способности отвечать реакцией СВЧ на последующее заражение авирулентной расой. Значит, в составе вирулентной расы есть какой-то фактор, который мешает отвечать картофелю защитными реакциями. Этот гипотетический фактор вначале так и назвали фактором совместимости. Присутствие этого фактора явно чувствовалось, когда из возбудителя фитофтороза картофеля начинали выделять индуктор защитных реакций липогликопротеидный комплекс. Неочищенный индуктор, выделенный из вирулентной расы паразита, оказался значительно менее активным, чем такой же, но выделенный из авирулентной расы. Явно чувствовалось, что в неочищенном индукторе присутствует некое вещество, которое мешает индуктору проявлять свою активность. Но мешает только в совместимой комбинации гриба и растения и не мешает, если комбинация является несовместимой. И такой супрессор был наконец выделен параллельно и независимо нами и объединенной группой американских и японских исследователей.
Ими оказались низкомолекулярные глюканы со связями β-1,3 в основной цепи и связями β-1,6 в местах разветвлений. Молекулярная масса глюканов составляла около 3000–4000 Дальтон.
Было испытано большое число комбинаций сортов хозяина и паразита, и всюду, где глюканы были выделены из совместимой к данному сорту расы гриба, они проявляли свойства супрессора, а если они были выделены из несовместимой расы, то таким действием не обладали. Иными словами, глюканы оказались специфическими, их действие в точности отражало взаимоотношения сорта и расы паразита.
Такие глюканы обнаружены внутри гиф паразита и в составе его выделений. Дело в том, что грибы рода Phytophthora имеют своеобразное строение клеточных стенок, которые на 75 % состоят из высокомолекулярных β-1,3—β-1,6-глюканов. Низкомолекулярные глюканы-супрессоры, по-видимому, служат строительным материалом для образования клеточных стенок, которые вместе с соответствующими ферментами в виде пузырьков везикул подходят к кончику гифы паразита, где и происходит построение клеточной стенки. Стейка гифы строится на ее растущем конце. Здесь глюканы изливаются из везикул и могут выделяться наружу и попадать в инфицированное растение, где и выполняют роль супрессоров.
Предполагается, что при взаимодействии паразита с цитоплазматической мембраной растения высокомолекулярные глюканы клеточных стенок (элиситеры) распознаются рецепторными белками на цитоплазматической мембране, индуцируя тем самым реакцию СВЧ. При совместимой комбинации хозяина и паразита взаимодействию элиситеров с рецепторами препятствуют низкомолекулярные глюканы — супрессоры, которые выделяются на конце растущей гифы.
На основании этих работ была предложена схема взаимодействия метаболитов хозяина и паразита, объясняющая на молекулярном уровне гипотезу ген — на — ген. Иными словами, кто кого преодолеет: элиситер супрессора или супрессор элиситера. Если рецепторный участок у растения захватит индуктор, включится устойчивость, если же, наоборот, супрессор — восприимчивость. Вспомните, в первой части книги мы предлагали гипотетическую схему возникновения генов устойчивости растения и вирулентности паразита в самых общих чертах и обещали интерпретировать ее на молекулярном уровне (рис. 13).
Гипотеза основывается на предположении, что у молекулы супрессора имеется два активных центра: неспецифическая группировка, которая конкурирует с элиситером за рецепторный участок у хозяина, и специфическая группировка, контролирующая распознавание молекул супрессора продуктами генов устойчивости растения (продуктами R-генов). Первая группировка молекулы супрессора, конкурирующая с индуктором, постоянна, вторая — изменчива, вариабельна.
Основным положением гипотезы является следующее: растение не поражается паразитом до тех пор, пока распознает на его поверхности элиситеры, которые служат сигналом для включения системы защиты. Для того чтобы преодолеть барьер неспецифического (видового) иммунитета, паразит приобретает супрессор, который, конкурируя с элиситером за рецептор растения и имея большее к нему сродство, занимает соответствующий рецепторный участок хозяина и тем препятствует включению защитных реакций с его стороны.
 Рис. 13. Предполагаемая схема молекулярного взаимодействия элиситоров в супрессоров в системе ген — на — ген
Рис. 13. Предполагаемая схема молекулярного взаимодействия элиситоров в супрессоров в системе ген — на — ген
В ходе сопряженной эволюции с паразитом у растения появляются гены сортовой устойчивости (или R-гены). Предполагается, что их продуктом является некое вещество, или рецептор, у растения, которое связывает супрессор по его вариабельной группировке и тем самым как бы «уводит» его с поверхности взаимодействия паразита и хозяина, оставляя элиситер вне конкуренции. Не имеющий более конкурентов элиситер вновь вступает во взаимодействие с рецептором, индуцируя у пего защитные реакции. Такое растение приобретает сортовую устойчивость к патогену, основанную на наличии у него R-гена.
В результате мутации вариабельная часть супрессора (та самая часть, которая связывалась продуктом R-гена) изменяется так, что теряет комплементарность к продукту R-гена и перестает им связываться. Супрессор вновь оказывается на свободе и начинает вытеснять элиситер в борьбе за рецептор. Так возникает ген вирулентности паразита, способный преодолевать ген устойчивости.
Далее процесс повторяется. Отдельные особи среди популяции растения приобретают способность узнавать и связывать вновь возникшую изменившуюся часть молекулы супрессора. Возникает ген устойчивости R
2, а паразит, изменяя свою вариабельную часть (ген вирулентности), преодолевает и эту уловку растения.
Вот вам тот же самый эволюционный марафон, с которого мы начинали книгу, но уже на молекулярном уровне. Растение убегает, паразит догоняет. Растение всегда впереди, стремясь уйти от инфекции, а паразит в лице преследующего легко нагоняет своего партнера.
Гипотеза не претендует на универсальность, к тому же она все еще остается одной из> многих гипотез, объясняющих взаимоотношения паразита и хозяина. Мы приводим ее, поскольку она наиболее наглядно позволяет изобразить те схемы взаимодействия продуктов генов устойчивости хозяина и авирулентности паразита, которыми сейчас увлекаются фитоиммунологи.
Кто-то назвал гипотезу строительными лесами вокруг здания, которые помогают его возводить, в том случае, если они поставлены верно. В противном случае их приходится разбирать и начинать все с самого начала.
Наиболее уязвимым местом этой и подобных гипотез является почти полное отсутствие сведений о рецепторе для индуктора и супрессора, кроме уверенности, что они должны быть. Фитоиммунологи последовательно вытягивают из темноты небытия цепь за цепью: сначала фитоалексины, которые позволили обнаружить элиситеры, элиситеры привели к обнаружению супрессора, а те и другие должны вытянуть рецепторы. Будем надеяться, что это вскоре произойдет.
СЕНСИБИЛИЗАЦИЯ,
ИЛИ ПОВЫШЕНИЕ ЧУВСТВИТЕЛЬНОСТИ
Термин сенсибилизация был введен в фитоиммулологию швейцарским фитопатологом Э. Гойманом. Под сенсибилизацией подразумевалось повышение реактивности организма, в итоге чего он становится способным к тому, к чему ранее способен не был. В медицинской иммунологии термин сенсибилизация используется уже давно. Например, состояние сенсибилизации развивается после первичного контакта организма с сывороточными белками другого вида животных, в результате чего наступает состояние резко повышенной чувствительности к этим белкам.
Состояние сенсибилизации у растений можно уподобить аллергической реакции животного организма. Аллергия (от греческого аллос — другой, аргон — действие) — повышенная норма реагирования организма животного и человека, наступающая после повторного введения болезнетворных микробов, чужеродных веществ и др.
У растений могут быть две формы сенсибилизации, одна из них развивается под влиянием самого возбудителя болезни, другая — под воздействием содержащихся в нем веществ. Первая форма сенсибилизации встречается в природе'. Так, заболевшее растение предохранено от реинфекции тем же, а иногда и другим патогеном. Однако подобная форма иммунитета, по мнению Гоймана, является лишь местной и проявляется либо в самом очаге заражения, либо рядом с ним. Исключение составляют вирусные инфекции, при которых вирусы наводняют весь организм хозяина и всюду вызывают местные защитные клеточные реакции, поэтому на первый взгляд создается впечатление гумморальной иммунизации, что, конечно, не соответствует действительности. Возможно, то же происходит и при трахеомикозах, когда возбудитель по сосудам распространяется по всему растению.
Что касается второй формы сенсибилизации, которая может быть достигнута под воздействием продуктов паразита, то она еще более ограничена в своем проявлении, чем первая. Гойман признавал ее только теоретически, поскольку считал, что в природе она не существует. Он полагал, что ввести в растение какой-либо продукт патогена еще возможно, по распределение его по растительным тканям уже маловероятно.
Мы столь подробно остановились на воззрениях крупнейшего фитопатолога лишь с целью показать, что даже он отрицательно относился к практической возможности иммунизации растений на основе их сенсибилизации. Невозможность иммунизации растений он объяснял двумя обстоятельствами:
1. Растения обладают слишком низкой температурой тела, для того чтобы в их тканях активно протекали защитные реакции. Так, клубень картофеля имеет собственную температуру 0,005 °C, тогда как температура человека составляет 36,6 °C. Если судить по количеству тепла, освобождающемуся при диссимиляции, то жизненные процессы в организме человека протекают в 7000 раз активнее, чем в клубне картофеля.
2. Клетки растений бедны протоплазмой. Если у животных клетки почти целиком заполнены протоплазмой, то у растений она только образует тонкий пристенный слой, тогда как все остальное пространство (примерно 9/10 объема) занимают вакуоли. А активные ответные защитные реакции протекают именно в протоплазме.
Все это и привело к выводу, что иммунизация растений, основанная на сенсибилизации, если и существует, то локальна и быстро преходяща.
И действительно, последующие исследования показали, что в зараженном растении устойчивость приобретают только клетки, либо расположенные в зоне заражения, либо непосредственно к ним примыкающие. Об этом писал еще Мюллер, основываясь на своих исследованиях по фитофторозу картофеля.
Противоположной точки зрения придерживался Н. И. Вавилов, который считал возможным наличие приобретенного иммунитета у растений. Успехи фитоиммунологии, достигнутые за последнее время, позволили подтвердить его пророческое предсказание. И все началось с обнаружения у патогенов метаболитов, которые распознаются растениями и выполняют у них роль элиситеров защитных реакций, о чем уже шла речь в предыдущей главе нашей книги. Например, оказалось, что элиситер, выделенный нами из возбудителя фитофтороза, липогликопротеидный комплекс вызывает у картофеля системную сенсибилизацию всего растения. Но для этого должно быть соблюдено, как минимум, два условия: 1) элиситер должен быть отделен от присутствующего у того же паразита супрессора (β-1,3-β-1,6-глюканов); 2) элиситер должен быть использован в строго определенной концентрации, не большей и не меньшей.
Высокие концентрации элиситера по вызывают сенсибилизации. Впрочем, слова «высокие концентрации» вряд ли вообще применимы к элиситеру ввиду его высокой биологической активности. Под высокими концентрациями подразумеваются концентрации элиситора порядка 100 мг/л, которые вызывают процессы образования фитоалексинов и некроза в обрабатываемой ткани. Еще не начав исследовании по иммунизации, мы твердо знали, что такие концентрации ни в коем случае не могут быть применимы для иммунизации, даже если бы и вызывали абсолютную защиту растений. Ведь фитоалексины — токсические соединения, обладающие широким спектром антибиотического действия. Однако оказалось, что этот вопрос — чисто риторический, поскольку те концентрации элиситера, которые вызывают в растениях некроз, не только не защищают их от последующего инфицирования, но, наоборот, в ряде случаев вызывают их восприимчивость. Почему же фитоалексины, будучи защитными веществами, не защищают растения, в которых образуются? Видимо, здесь действует тот принцип, о котором мы уже не раз говорили: антибиотическое вещество может оказать защитное действие, только если оно образуется в нужном месте, в нужное время и в нужной концентрации. Ни того, ни другого, ни третьего не происходит при обработке тканей высокими концентрациями элиситера. Фитоалексины при этом образуются заранее, локализуются в тех клетках, которые некротизируются под действием элиситера, да к тому же могут и вообще исчезнуть к моменту инфицирования, поскольку метаболизируются растительной тканью. Паразит же при последующем инфицировании, естественно, обходит погибшие клетки и проникает в соседние интактные, которые уже израсходовали свои пластические и энергетические ресурсы на образование фитоалексинов для соседних некротизированных клеток.
Все дело в том, что фитоалексины в отличие от антител нельзя заготовлять впрок, как это происходит при иммунизации человека и животных. Во-первых, они вскоре после накопления исчезают в ходе метаболизма, так как являются эндогенными ксенобиотиками, а во-вторых, они не могут транспортироваться из одних некротизированных клеток в другие. Антитела же свободно передвигаются по кровяному руслу и могут длительное время сохраняться в нем, поскольку не представляют опасности для организма, а нацелены только на строго специфическую мишень — определенный чужеродный антиген.
Итак, те концентрации липогликопротеидного комплекса, которые вызывают образование некроза, не могут предохранить растения от инфекции. Это могут сделать другие, значительно более низкие концентрации элиситера, порядка 5—10 мг/л. При обработке тканей этими концентрациями фитоалексины в них не накапливаются и клетки не некротизируются, но последние в два раза меньше поражаются совместимыми расами возбудителя фитофтороза, поскольку превращаются в сенсибилизированные.
Хотя такие клетки и не образуют фитоалексины в ответ на обработку элиситером, они оказываются хорошо подготовленными к последующей встрече с инфекцией. Если предварительно обработанные элиситером сенсибилизированные клетки заразить совместимой к ним расой возбудителя фитофтороза, то они значительно скорее, чем контрольные, отвечают реакцией СВЧ, образованием фитоалексинов и раневой перидермы, т. е. быстро и интенсивно реагируют на инфекцию комплексом ответных реакций, позволяющих им быстро локализовать инфекцию. Иными словами, ведут себя как несовместимые.
За счет чего же это происходит? Проведенные электронно-микроскопические наблюдения показали, что в сенсибилизированных клетках происходит определенная перестройка ультраструктуры. В большинстве клеток клубня картофеля в пластидах откладываются зерна крахмала, тогда как протоплазма занимает крайне малый объем и локализуется пристенно. Смысл сенсибилизации как раз и состоит в том, чтобы превратить клетки из метаболитически инертных — запасающих в метаболитически активные — биосинтезирующие. Доля цитоплазмы в сенсибилизированных клетках возрастает, увеличивается число митохондрий, возрастает число лейкопластов. Все это свидетельствует о приведении клеток в боевую готовность в том случае, если им предстоит встретиться с инфекцией.
Однако дело не сводится только к возрастанию общей метаболитической активности клетки. Нет. Такие клетки приобретают специализированные особенности, свидетельствующие о том, что их обмен переключен на синтез фитоалексинов. Последние у картофеля, как это ранее уже говорилось, относятся к числу сесквитерпеноидных соединений. А согласно современным представлениям, терпеноиды в клетке синтезируются в двух типах органел, или, как еще говорят, в двух компартментах. Первый — это агранулярный эндоплазматический ретикулум, а второй — ретикулум пластидного аппарата. Некоторые исследователи считают, что оба компартмента действуют независимо, т. е. одна часть терпеноидов синтезируется в первом, а другая — во втором, другие полагают, что они действуют последовательно. Иными словами, в одних компартментах начинается процесс биосинтеза, в других — он закапчивается. Но это уже детали.
Так вот, в сенсибилизированных клетках происходят именно эти изменения: в ультраструктуре доля агранулярного ретикулума возрастает, увеличивается также количество полиморфных лейкопластов. Все это свидетельствует о том, что сенсибилизированные клетки приобретают отдельные черты, характерные для специализированных терпеноидных клеток. В таких клетках уже осуществляется биосинтез сесквитерпеноидов, а не накапливаются они в силу того, что образующиеся фитоалексины тут же разлагаются в процессе их метаболизма.
Если такие клетки заразить, то им не придется затрачивать время и средства для того, чтобы создавать аппарат для биосинтеза фитоалексинов, он уже подготовлен, и именно поэтому они быстрее и интенсивнее отвечают защитными реакциями на инфекцию. Очень вероятно, что подготовка не ограничивается только биосинтезом терпеноидов, но состоит также в ускоренном образовании фенольных соединений и суберина, которые также несут защитные функции.
Однако главное, что удалось установить, это то, что возникающая под воздействием элиситера сенсибилизация является отнюдь не локальной, но системной. В фитоиммунологии существуют понятия локальной и системной устойчивости, о которых мы вскользь уже упоминали. Локальная устойчивость развивается только в месте развития инфекции или нанесения препарата и не распространяется по растительным тканям. Напротив, системная устойчивость проявляется у растения на расстоянии от точки взаимодействия с патогеном или веществом и захватывает все растительные ткани. Так вот, устойчивость, вызываемая липогликопротеидным комплексом, оказалась не локальной, а системной.
Это значит, что она не ограничивается только теми тканями, на которые был нанесен элиситер, а распространяется от него в глубь клубня. Если клубень картофеля опрыснуть элиситером в сенсибилизирующей концентрации, то через 1–2 дня защищенными окажутся поверхностные ткани, через 3–4 дня — ткани, расположенные за ними, и, наконец, через 7 дней сенсибилизация достигнет середины клубня и весь клубень окажется защищенным от болезней. Для достижения состояния сенсибилизации необходимо время, в течение которого и происходит упомянутая перестройка ультраструктурного аппарата клетки. Пройдет несколько месяцев. И точно так же в таком же порядке сенсибилизация будет постепенно отступать: сначала она исчезнет из центра клубня, потом из его серединной части и последней из периферических слоев.
Мы пробовали усилить защитные свойства индуктора, давая не одну, а две с интервалом в один день обработки клубня. Однако таким образом усилить эффект сенсибилизации не удалось. Зато его можно продлить, если на исходе первичной сенсибилизации еще раз иммунизировать клубни, в результате этого сроки сенсибилизации вдвое удлиняются.
Для сенсибилизации клубня необязательно, чтобы он весь был обработан элиситером, достаточно его нанести на часть клубня с одним обязательным условием: элиситер должен попасть на глазки. Вероятно, именно через глазки элиситер и проникает внутрь клубня, поскольку покрывающая клубень перидерма является для него естественной преградой. Кроме того, и это наиболее вероятно, сигнал сенсибилизации передается по сосудистой системе клубня, которая подходит к глазкам.
Эффект сенсибилизации оказался настолько сильным, что не ограничивался только клубнями, но распространялся с них на выросшие из обработанных клубней растения и далее на клубни нового урожая. В течение 7 лет опыты, проведенные совместно с Белорусским научно-исследовательским институтом картофелеводства и плодоовощеводства, показали, что предпосадочная обработка клубней картофеля индуцировала устойчивость вегетирующих растений, а затем клубней нового урожая. К весне устойчивость постепенно исчезала и вновь появившиеся растения мало чем отличались по своей устойчивости от контрольных. Об этих опытах подробнее мы будем говорить в главе об иммунизации.
Замечательно, что элиситер, выделенный из возбудителя фитофтороза, защищает клубни не только от фитофтороза, но и от целого комплекса болезней, вызываемых паразитарными грибами (фитофтороза, ранней сухой пятнистости, ризоктопиоза и парши). Меньшее поражение этими болезнями наблюдалось как на вегетирующих растениях картофеля, так и на полученном от них урожае клубней. Это не удивительно, поскольку одним из защитных механизмов сенсибилизации является образование фитоалексинов, которые, как известно, подавляют рост многих паразитарных грибов.
Сам индуктор, липогликопротеидный комплекс — не обладает свойством расовой специфичности. Это означает, что он присутствует у всех исследованных рас паразита, независимо от наличия у них генов вирулентности, и с его помощью может быть индуцирована устойчивость у всех сортов картофеля, независимо от присутствия у них генов фитофтороустойчивости. Так и получилось. Иммунизация позволяла защитить от инфекции все исследованные сорта картофеля, даже те, которые не обладают ни одним геном фитофтороустойчивости. Таким образом, неснецифическое действие элиситера в данном случае оказалось на руку исследователям.
Ну а как же специфический супрессор, который, как мы это уже ранее говорили, обладает антиэлиситерным действием и, следовательно, должен препятствовать действию элиситера. Представьте себе, что на поле произрастают защищенные липогликопротеидным комплексом сенсибилизированные растения и на них попадают споры вирулентных рас возбудителя фитофтороза. Ведь у этих рас есть супрессоры, преодолевающие действие элиситера. Почему же они не снимают его сенсибилизирующий эффект? Значит, либо супрессоры не обладают антиэлиситерным действием, либо комплекс не может защитить растения от совместимых рас патогена. Начались исследования, которые дали следующий ответ. Супрессор может только предотвратить наступление сенсибилизации, вызываемой элиситером, по не может нарушить уже наступившую. Вся суть действия супрессоров, по-видимому, и состоит в том, чтобы предотвратить, не допустить той ультраструктурной перестройки, которая происходит в сенсибилизированных тканях. Если подобная ультра-структурная перестройка уже произошла, то супрессор оказывается беспомощным и ничего не в состоянии сделать. Именно поэтому он не способен нарушить устойчивость в уже сенсибилизированных клубнях и растениях картофеля в полевых условиях.
Какие же защитные реакции, кроме реакции СВЧ, образования фитоалексинов и раневой перидермы могут объяснить высокий уровень устойчивости сенсибилизированной ткани. Отдельные эксперименты позволили предполагать, что в сенсибилизированных тканях ускоряются процессы лигпификации, активизируется процесс образования ингибиторов протеиназ, а также ферментов пероксидазы и полифенолоксидазы.
Совершенно открытым остается интереснейший вопрос о том, что же является химическим посыльным сенсибилизации? Что такой посыльный существует, сомнений быть не может, хотя бы потому, что липогликопротеидный комплекс вряд ли способен проникнуть в ткани внутренних слоев клубня картофеля. Скорее всего, как и в случае со вторичным иммунологическим курьером, здесь имеется некий посыльный,
который дает сигнал к сенсибилизации клеток. Однако, что он из себя представляет и как распространяется, пока еще неясно.
Практически сенсибилизация, о которой речь шла в этой главе, является прообразом полигонной, полевой, относительной или неспецифической устойчивости. Об этом свидетельствуют следующие факты: 1) защита от болезней не является абсолютной, а составляет около 50 %; 2) защитные свойства не являются специфическими, поскольку проявляются по отношению ко всем исследованным патогенам и их расам.
Однако именно неспецифическая устойчивость, как известно, является наиболее перспективной для селекционеров и позволяет избежать опустошительных эпифитотий и поддерживать в природе то равновесное состояние, которое сложилось между хозяином и паразитом.
Сенсибилизация может встречаться в природе, примером чему могут служить ткани растений, несущие в себе микоризу. Известно, что в таких растениях повышено количество ингибиторов, в результате чего в корне и вокруг него создается токсическая зона, предохраняющая их от поражения многими почвенными микроорганизмами. Таким образом, даже заражение микоризой растение оборачивает себе на пользу, повышая за счет этого свой защитный потенциал. В самом растении устанавливается своеобразный вооруженный мир, который обычен для партнеров — симбионтов; растение сдерживает агрессивность грибов микоризообразователей, не позволяя им из симбионтов превратиться в паразитов, а грибы со своей стороны очень умеренно используют питательные вещества растения.
Сейчас, когда многое прояснилось, можно предположить, почему исследователей прошлых лет преследовали неудачи в их попытках иммунизировать растения. Причин этому несколько:
1. Не было известно, чем именно следует иммунизировать растения. Попытки иммунизировать растительные ткани проводились вслепую. Поскольку о существовании элиситеров не подозревали, в рабочие растворы, которыми обрабатывали растения с целью повысить их устойчивость, элиситеры могли и не попадать. Единых методов получения иммунизирующих растворов нет и быть по могло, поскольку элиситеры у разных патогенов различны по химической природе и по-разному локализованы: одни находятся на их поверхности, другие в составе выделений, третьи локализованы внутриклеточно, но выходят из клеток паразитов при соприкосновении с растением.
2. В том случае, если даже элиситер попадал в состав иммунизирующей жидкости, он мог попасть в нее в иной, чем это необходимо, концентрации. Мы уже говорили о том, что, для того чтобы растение оказалось в состоянии сенсибилизации, элиситер в составе рабочей жидкости должен быть в строго определенной концентрации — не большей и не меньшей. Малая концентрация элиситера окажется недостаточной для сенсибилизации, тогда как слишком большая вызовет некроз и образование фитоалексинов. Существует довольно ограниченная область концентраций, в которой элиситер вызывает нужный эффект, и эта концентрация вряд ли может быть случайно достигнута.
3. Если даже допустить, что элиситер оказался в рабочих растворах и попал в них в нужной концентрации, то его действие могло оказаться перекрытым супрессором, присутствующим у этого же патогена. Иными словами, выделить элиситер из паразита, отделив его от супрессора, и затем использовать в строго определенной концентрации вслепую, конечно, не представлялось возможным, чем и объясняются неопределенные результаты прошлых опытов по иммунизации растений,
ОТ ТЕОРИИ К ПРАКТИКЕ
Суха, мой друг, теория везде, а древо жизни пышно зеленеет.
И. Гете[18]
ИНТЕГРИРОВАННАЯ СИСТЕМА
ЗАЩИТЫ РАСТЕНИИ
Главным в современной тенденции защиты растений является стремление к объединению всех доступных методов борьбы с вредителями и болезнями. Вот это-то и носит впечатляющее название — интегрированная система защиты растений. Другое дело, что удельный вес каждого из используемых методов совершенно различен. В целом современная стратегия защиты растений основывается на двух главных методах — это выделение устойчивых сортов и применение пестицидов. Этим методам посвящены две специальные главы настоящей книги. Здесь же поговорим о других мероприятиях.
Одним из них является карантинная служба. Так, законодательным порядком запрещается ввоз зараженного растительного материала на территорию тех стран, где вредители и болезни ранее отсутствовали. В большинстве стран Имеются законы, контролирующие импорт растительного материала или его перевозку из одной части страны в другую. Например, благодаря этим законам в Англии до сих пор отсутствует столь страшный враг картофеля, как колорадский жук. Правда, карантин следует считать только первой линией обороны, которая рано или поздно будет нарушена.
Уменьшить ущерб, причиняемый вредителями или болезнями, может также фитосаинтария — одни из наиболее простых и наиболее древних способов защиты растений. Фитосанитария сводится к уменьшению количества ипокулума при уничтожении любого зараженного растительного материала, будь то больные растения или их остатки в поле после уборки урожая. Фитосанитарные мероприятия, если их проводить достаточно тщательно, могут принести ощутимый эффект, особенно в садах и огородах, где сравнительно легко уничтожить весь зараженный растительный материал.
Очень существенным для целей защиты растений является высокая культура земледелия. Задача земледелия состоит не только в том, чтобы создать условия, обеспечивающие наилучший рост растений и высокий урожай, но и в том, чтобы защитить растения от патогенов и вредителей. Прежде всего чрезвычайно важен качественный посадочный материал. Для посева следует использовать только здоровые, зрелые, неповрежденные семена, обладающие высокой всхожестью. Посев должен обеспечить оптимальную для данной культуры густоту стояния растений. Очень вагина глубина посева. Длительный период между прорастанием и появлением всходов над поверхностью почвы благоприятствует развитию патогенов. Поэтому факторы, тормозящие прорастание и начальный рост растений, способствуют поражению растений и увеличивают их потери еще на ранних стадиях развития.
Растения, восприимчивые на одной стадии развития, в другое время проявляют большую устойчивость. В связи с этим возможно так изменять сроки посева, чтобы стадия развития, на которой растения являются наиболее восприимчивыми, приходилась на то время, когда патоген отсутствует или не может заразить растения.
На поражаемость растений болезнями влияют удобрения. Считается, что высокое содержание в почве азота, особенно если он находится в легкоусвояемой форме, например в виде нитратов, обычно повышает восприимчивость растений к болезням. Возрастание содержания в почве калия и фосфора неблагоприятно сказывается на развитии некоторых патогенов.
Очень полезным способом защиты растений от патогенов является севооборот, который дает особенно хороший результат в борьбе с почвенными патогенами. В случае болезни корней он является одним из главных способов защиты. Длительность сохранения патогенов в почве зависит от долговечности их зимующих форм, от микробиологических и химических условий в почве и других факторов. Это очень важно для эффективности севооборотов, потому что защита от болезней может быть успешной лишь в том случае, если интервал между посевами восприимчивых культур больше, чем срок жизни патогена.
Таким образом, для решения проблемы защиты растений следует познать естественные связи в биосфере, понять, какие из них нарушаются, и попытаться их компенсировать.
А теперь вернемся к двум основным мерам, на которых основывается современная интегрированная система защиты растений.
СОРТ РЕШАЕТ УСПЕХ ДЕЛА
Ничто другое так не способствует защите растений, как развитие селекции сортов, устойчивых к болезням и вредителям. Именно благодаря широкому распространению устойчивых сортов некоторые вредители и болезни к настоящему времени уже не представляют опасности.
Устойчивость — только одна из многих целей селекции, хотя и одна из наиболее важных. Ведь даже высокоустойчивые сорта вряд ли будут выращиваться, если они малопродуктивны и худшего качества, чем остальные. Часто эти два фактора противоречат друг другу. Поэтому обычно селекционер вынужден искать компромисс между стремлением добиться у создаваемого им сорта приемлемых уровней урожайности, качества и других агрономических признаков, с одной стороны, и повышением уровня устойчивости — с другой.
Одна из задач современной селекции — создание сортов, обладающих комплексной устойчивостью, т. е. одновременной устойчивостью к нескольким болезням и вредителям, поскольку в сельском хозяйстве невыгодно выращивать сорта, устойчивые только против одного паразита, и в то же время регулярно применять пестициды против другого.
Большинство видов культурных растений было создано в период примитивного ведения сельского хозяйства, вначале благодаря естественному, а затем сознательному отбору земледельцами растений с желательными признаками. Это был период отбора сортов — популяций, в ходе которого земледелец оставлял на потомство наиболее выносливые и урожайные экземпляры.
Таким образом, до сравнительно недавнего времени культурные растения были представлены популяциями, приспособленными к местным условиям и дающими удовлетворительные урожаи качественной продукции. Однако у них было очень важное преимущество: они были гетерогенными по наличию в них генов вообще и генов устойчивости в частности.
Затем наступило время выведения чистолинейных сортов культурных растений, имеющих одинаковые гены, и, следовательно, популяции таких растений оказались гомогенными. А отсюда пошли все беды. Принцип монокультуры начал повсеместно применяться в сельском хозяйстве. На огромных площадях высевались одни и те же сорта, у которых все признаки, в том числе и признак устойчивости, были выравнены. Культуры стали однородными, они давали наивысшие возможные урожаи, одновременно созревали, росли на стеблях одной высоты и плотности, по, к сожалению, одинаково хорошо были восприимчивы к болезням. А история эпифитотий учит, что узкая генетическая основа посевов резко ограничивает их способность сопротивляться болезням. Жизнь устойчивых сортов становилась все короче и короче. В этих условиях селекционерам приходилось работать по непрерывной программе, т. е. быстро выводить один сорт на смену другому с тем, чтобы опередить расообразовательные процессы у паразита. Необходимы были новые методы борьбы с болезнями, которые сейчас усиленно разрабатываются.
О некоторых из них попробуем рассказать. Как ужо известно из предыдущего изложения, чаще всего, хотя и не всегда, паразит преодолевает вертикальную устойчивость растений, основанную на свойстве СВЧ. Такой тип устойчивости связан с присутствием в растении единичных генов, которые относительно просто вводить во вновь создаваемые сорта. Значительно сложнее обстоит дело с горизонтальной устойчивостью, которая контролируется многими генами, а потому с ними значительно сложнее работать селекционеру. До самого недавнего времени большинство селекционеров при наличии выбора предпочитали работать с явно выраженной вертикальной устойчивостью. При этом часто случалось, что в процессе селекции на вертикальную устойчивость у сорта терялись гены горизонтальной устойчивости. Судьба многих сортов культурных растений с вертикальной устойчивостью оказалась плачевной. Их первоначальный успех во многих случаях со временем сменялся полным поражением. Поэтому сейчас внимание селекционеров все более и более привлекает горизонтальная устойчивость. Создается парадоксальная ситуация: относительно слабый уровень горизонтальной устойчивости оказывается выгоднее, чем полная вертикальная устойчивость. Но все дело в том, что вертикальная устойчивость рано или поздно в большинстве случаев преодолевается расообразовательными процессами паразитов, тогда как горизонтальная устойчивость оказывается, как правило, долгосрочной. Эффект ее хотя и недостаточен, но зато действие стабильно. Особенно перспективным представляется сочетание в одном и том же сорте и вертикальной, и горизонтальной устойчивости.
Как известно, основной принцип защиты растений состоит в предотвращении эпифитотий. В случае сочетания горизонтальной и вертикальной устойчивости последняя задерживает начало эпифитотий, поскольку необходимо, чтобы раса с определенным геном вирулентности накопилась в популяции паразита до определенного уровня. Когда она, хоть и с запозданием, накапливается, тогда вступает в действие горизонтальная устойчивость, которая замедляет развитие болезни уже после ее начала. Благодаря присутствию у сорта горизонтальной устойчивости на нем образуется меньшее число спор, дающих начало новым поражениям, сами поражения развиваются медленнее, больше времени требуется для формирования нового поколения спор и т. д. В результате развития эпифито-тий не происходит.
Предполагается каждый сорт, обладающий тем или иным геном устойчивости, высевать раз в несколько лет, с тем чтобы в промежутке стабилизирующий отбор уничтожал расы, обладающие излишними генами вирулентности. Целесообразно также на пути миграции спор паразита высевать сорта, различающиеся генами устойчивости. При этом должны учитываться места зимовки инфекции, направление ветра, с помощью которых распространяются споры паразита. Подобная «чересполосица» предотвратит накопление инфекционного начала, необходимого для начала эпифитотий, поскольку при попадании спор с одной полосы на другую их популяция должна будет перестраиваться, в результате чего инфекция не сможет накопиться до нужного для эпифитотий уровня.
Таким образом, на смену ранее существующего принципа максимальной однородности, выравненности сортов по всем признакам, в том числе и по устойчивости, сейчас приходит новый принцип наибольшей гетерогенности при максимально возможном сохранении качества сорта. Происходит своеобразный возврат к прежним крестьянским сортам-популяциям, с той, однако, разницей, что они выводятся не эмпирически, а на основании современной теории. Примером тому являются современные многолинейные или мультилинейные сорта-популяции.
Многолинейные сорта выращивают, смешивая семена нескольких составляющих их линий. По сути дела, эти липни почти изогенные, которые выравнены по всем (или по большинству) признаков, кроме одного — генов устойчивости. Предположим, колосья их равны по высоте, в одной и то же время созревают, выравнены по мукомольным качествам и так далее. Поэтому поле, засеянное многолинейным сортом, внешне кажется совершенно однородным и ничем не отличающимся от поля засеянного чистолинейным сортом. Но для паразита растущие на поле растения резко различаются, поскольку являются для него смесью различных генотипов хозяина, затрудняющих их массовое поражение.
Предположим, что создается четырехлинейный сорт, включающий в себя композицию генов вертикальной устойчивости R
1, R
2, R
3, и R
4. в равных соотношениях. Раса 1 способна поражать сорт с геном R
1, но не сорта с геном R
2, R
3 и R
4. Раса 2 поражает сорт с геном R
2, но не остальные, раса 3 — с геном R
3 и так далее. Раса 1.2.3.4 может в равной мере поражать все составляющие сорт линии, но на каждой из линий она имеет по три ненужных (лишних) гена вирулентности, и поэтому стабилизирующий отбор будет работать против нее, в результате чего она не сможет выдержать конкуренции с простыми расами, не имеющими лишних генов. То же самое будет происходить и с расами, несущими по три гена вирулентности, — 1.2.3, 1.2.4, 1.3.4 и 2.3.4, с той разницей, что у них будут лишними не три, как у расы 1.2.3.4, а по два гена, а также с расами 1.2, 1.3, 1.4, 2.3, 2.4 и 3.4, у которых будет по одному липшему гену.
Простые расы 1, 2, 3 и 4 не будут нести лишних гемов, по каждая из них сможет поразить только 25 % популяции, тогда как 75 % окажутся к ней устойчивы. В результате получится, что все расы паразита, которые могут существовать на посевах многолинейного сорта-популяции, будут плохо развиваться. Сложные расы будут ослабляться стабилизирующим отбором, лишающим их конкурентоспособности с простыми расами, а простые расы страдать от затрудненного распространения (считается, что популяция растений оказывается защищенной, если не менее 40 % растений в ней обладают устойчивостью). И то и другое приведет к снижению скорости инфекции, что характерно для горизонтальной устойчивости. Поэтому внешне многолинейный сорт будет вести себя почти так же, как и сорт горизонтально устойчивый.
Чем больше число линий, составляющих многолинейный сорт, тем больше будет эффект. К тому же и композицию отдельных сортов можно каждый год менять в соответствии с составом генов популяции паразита. Из-за этого популяция паразита будет все время вынуждена перестраиваться, и угроза эпифитотий тем самым ликвидируется.
Таким образом, мультилинейные сорта будут совмещать в себе преимущества чистолииейиых сортов (выравненность по всем хозяйственно цепным признакам, кроме генов устойчивости) и сортов-популяций прошлого, поскольку они будут гетерогенными по генам устойчивости. Сейчас создаются специальные селекционные программы по получению мультилинейных линий-популяций для пшеницы, овса, ячменя и других цепных культур.
Согласно мнению IO. Т. Дьякова, существующий сейчас в защите растений принцип «слабая поражаемость лучше полной устойчивости» представляет собой один из парадоксов фитоиммунитета, поскольку создание абсолютно непоражаемого сорта не только не осуществимо, по и порочно в своей основе. Маловероятно уничтожить какой-либо вид патогенного микроорганизма, поскольку он имеет преимущества перед растением-хозяином из-за способности быстро размножаться и мигрировать на большие расстояния. Но даже если и допустить, что это удастся, то на освободившуюся экологическую нишу всегда найдется какой-либо претендент, потенциальную опасность которого заранее трудно предсказать. Так что лучше не рисковать.
Исходя из вышесказанного, решение проблемы защиты культурных сортов от эпифитотий предполагает установление того равновесия между растениями и их паразитами, которое было нарушено чистолинейной селекцией, и лежит на пути расширения генетического разнообразия культурных сортов. Создавая и внедряя монокультуры, человек переусердствовал и слишком решительно и недальновидно вмешался в законы природы, чем не замедлили воспользоваться патогены.
Селекция на устойчивость — длительный процесс, и часто требует не менее 10–15 лет упорного труда. Правда, развитие теории, так же как и оснащение селекционного процесса современной техникой и оборудованием, позволяет сейчас сократить эти сроки. В поисках доноров устойчивости чрезвычайно перспективным представляется искусственный мутагенез, с помощью рентгеновских и ультрафиолетовых лучей, гамма-излучения или нейтронов, а также химических мутагенов. Под влиянием этих воздействий возникают индуцированные' мутации, которые могут дополнить природное разнообразие таким образом, что шансы на редкие гены будут увеличены. В результате разработки новых более топких методов индуцированные мутации будут становиться все более важными в селекции на устойчивость, особенно по мере истощения естественных источников генетического разнообразия. Обнадеживают также новые методы клеточной селекции и слияние протопластов для получения гибридов между нескрещивающимися сортами.
Устойчивые сорта в будущем, по-видимому, будут играть все более и более важную роль в борьбе с вредителями и болезнями растений. И в этом отношении селекция на устойчивость начинает успешно конкурировать с разработкой и применением пестицидов.
ПЕСТИЦИДЫ:
ВЧЕРА, СЕГОДНЯ, ЗАВТРА
История биологии знает немало событий, взволновавших широкую общественность. Однако есть события совершенно особые, и к ним можно отнести применение пестицидов.
Пестициды в дословном переводе с латинского — заразоубивающие средства (пестис — зараза, цедерс — убивать). Сам термин «пестициды» собирательный: это и инсектициды, используемые для борьбы с насекомыми (инсектум — насекомые); и фунгициды, направленные против паразитарных грибов и бактерий (фунгус — гриб); и гербициды против сорняков (герба — трава); и зооциды против грызунов; и нематоциды против нематод. Имеются и другие пестициды, обладающие комплексным действием. Например, нитрофан является инсектицидом против тлей, клещей, щитовок и фунгицидом для борьбы с возбудителями таких болезней, как рак, антракноз, мильдью.
Наибольший накал страстей ученых и общественности, несомненно, вызвал инсектицид под сокращенным названием ДДТ — дихлордифенилтрихлорэтан. В его оценке одно быстро сменяло другое. Радужные надежды и горькие разочарования, восторженные отзывы и тяжкие обвинения. Пестициды называли чудом современной химии и непростительной ошибкой ученых.
ДДТ был синтезирован еще в 1874 г. Но только в 1937 г. было открыто его инсектицидное действие. Автору открытия швейцарскому ученому П. Рихтеру была присуждена Нобелевская премия. Двадцать лет спустя Швейцария запретила применение этого препарата.
Чем же так провинился ДДТ, и почему он впал в такую немилость? По данному вопросу написаны тысячи статей и высказаны взаимоисключающие точки зрения.
Еще продолжалась вторая мировая война, когда ДДТ одержал свою первую крупную победу. С его помощью удалось предотвратить вспышку сыпного тифа. Прошло немного лет, и с помощью ДДТ удалось подавить размножение комаров — переносчиков возбудителей малярии, и тем самым спасти жизнь миллионов людей. Затем последовали победы над многими сельскохозяйственными вредителями. Америка вдвое увеличила сбор хлопка благодаря защите хлопчатника от долгоносика. В Советском Союзе были сохранены миллионы тонн пшеницы от вред-пой черепашки. Началось триумфальное шествие препарата по странам и континентам. За 20 лет во всем мире его было использовано около 4,5 миллионов тонн.
Но постепенно над ДДТ сгущались тучи. Появились серьезные опасения, что пестицид оказывает вредное влияние на окружающую среду. Однако вера в пего была настолько сильна, что вначале опасения вообще не воспринимали всерьез, а капиталистические монополии не желали считаться ни с чем, кроме прибыли. Отчеты ученых просто скрывали. Целых семь лет власти США не разрешали опубликовать отчет об ответственности ДДТ за гибель тихоокеанской рыбы.
Отчет был составлен специалистом по исследованию качественного состава воды Уолтером Томсоном. Сравнение содержания пестицида на полях Америки с показателем уловов сардинки привело к «ошеломляющим» результатам: поголовье сардинок убывало в такой же пропорции в какой возрастало применение ДДТ. Попадание пестицида в районы лова рыбы объяснялось размыванием почвы при разливах рек, которые несли его с собой в океан. Он оказался ядовитым не только для рыб, по и для организмов, которыми они питаются.
Последовали и новые отчеты о вреде ДДТ для других организмов, в частности птиц. Скрывать их от общественности становилось все труднее. Гром разразился в 1962 г. с выходом в США книги Речел Карсон под названием «Безмолвная весна». В ней изображены страшные последствия применения ДДТ. Не слышно пения птиц, не плавают в водоемах рыбы, многие виды животных на грани полного исчезновения. В США книга вызвала бурю протестов (подогреваемых радио, телевидением, газетами) против ДДТ и других пестицидов, которые могли принести вред окружающей среде.
В ноябре 1972 г. на заседании Продовольственной и сельскохозяйственной организации ООН (ФЛО) выступил с лекцией Норман У. Борлоуг — лауреат Нобелевской премии мира 1970 г. за личный вклад в так называемую зеленую революцию. Лекция была названа «Человечество и цивилизация снова на распутье». В ней была отдана дань уважения Р. Карсон, сделавшей очень много для понимания красоты природы. Но книга была подвергнута резкой критике, прежде всего за то, что она внушала читателям, что без пестицидов будто бы можно обойтись. По мнению Берлоуг, такой день если он вообще наступит, пока еще очень далек.
Лекция была страстной, но одновременно и пристрастной. Теории противников были объявлены шаткими, а приводимые факты сокращения численности животных объяснялись не столько применением пестицида, сколько другими причинами нарушения экологического равновесия, например сбросом в водоемы отходов промышленных предприятий.
Факты, однако, со всей неоспоримостью говорили об отрицательном влиянии ДДТ. Приведем лишь некоторые
[19].
Если в воде концентрация ДДТ составляет 0,000005 единицы, то в планктоне она достигает 0,04 единицы, т. е. увеличивается почти в 10 тысяч раз; в мелкой рыбе, питающейся планктоном, увеличивается до 0,5 единицы; в крупной рыбе, пожирающей мелкую, до 2. Тогда как у птиц, кормящихся крупной рыбой, до 2,5, т. е. в 500 тысяч раз выше по сравнению с исходной концетрацией. Превышение содержания ДДТ в фитопланктоне лишь на несколько частей на миллиард уже отражается на процессе фотосинтеза. А ведь фитопланктон производит почти 70 % кислорода, содержащегося в воздухе.
На острове Калимантан в Индонезии ящерицы «устроили пир» из мух, убитых ДДТ. Но это поистине был пир во время чумы, так как вскоре погибли и ящерицы. Вслед за ними стали гибнуть и кошки, питавшиеся ящерицами. В результате крысы, которых уничтожали кошки, наводнили деревни и создали угрозу возникновения эпидемий.
Но это лишь одна сторона проблемы. Другая, не менее острая, состоит в том, что вредители, против которых применялся ДДТ и считавшиеся к нему весьма чувствительными, в итоге меньше всего от него пострадали. Объясняется это тем, что они приобрели к нему настолько высокую устойчивость, что даже при увеличении концентрации препарата в несколько раз против разрешенных нормами, они оставались живыми. Более того, насекомые, ставшие устойчивыми к ДДТ, приобрели устойчивость и к некоторым другим хлорорганическим пестицидам. Неудивительно, что в большинстве стран мира, в том числе и в нашей стране, применение ДДТ было запрещено в законодательном порядке или крайне ограничено. Заметим, что в нашей стране, когда ДДТ применялся, допустимые его остатки в полевых, плодовых и овощных растениях были самыми низкими: в 7 раз ниже, чем в США и Канаде, в 2 раза ниже, чем в Швейцарии. И сейчас у нас действуют самые строгие критерии оценки всех пестицидов.
Итак, ДДТ во многом помог, во многом навредил, но и многому научил, хотя и дорогой ценой. Этот урок следует всегда помнить, когда речь заходит о применении пестицидов, без которых, по признанию ведущих ученых мира, пока обойтись нельзя.
Подсчитано, что в настоящее время чуть ли не каждые 10 минут в мире создается соединение, обладающее пестицидными свойствами. Но подавляющая их часть отбрасывается уже после первичного отбора (скрининга). После полевой оценки и определения безвредности остаются считанные единицы. И в этом тоже один из уроков, извлеченных из применения ДДТ.
В прошлом при испытании пестицидов ограничивались определением степени их токсичности путем скармливания препаратов животным. В дальнейшем уже стали определять, как быстро препарат выводится из организма и не накапливается ли он в каком-либо органе, например в печени. Далее начали выяснять, а не оказывает ли препарат или продукты его распада, возникающие в организме, мутагенное и канцерогенное действие.
Правда, чрезвычайно низкий выход пестицидов из огромного числа синтезируемых соединений связан еще с несовершенством самого принципа их синтеза и первичного отбора, основанного на методе проб и ошибок. Вполне попятно, что при этих условиях с каждым годом стоимость нового пестицида сильно возрастает. По мнению специалистов, при более рациональном использовании ужо существующих пестицидов они могут давать вдвое больший эффект по сравнению с ныне получаемым. Особо важное значение имеет улучшение службы прогнозирования времени появления вредителей и возбудителей болезней, а также усовершенствование техники нанесения пестицидов.
Пестициды могут быть эффективны только в том случае, если они применяются в нужное время, в нужном количестве и нужном месте. В противном случае пестициды приносят больше вреда, чем пользы. Несоблюдение этого требования служит причиной порою возникающих и поныне бесплодных дискуссий о пользе или вреде химизации сельского хозяйства.
Поскольку наша книга посвящена устойчивости растений к болезням, остановимся лишь на фунгицидах.
Фунгицидами называются вещества, убивающие или подавляющие споры грибов или их мицелий. Еще с древних времен было известно, что подобным действием обладает сера. О ней вновь вспомнили во Франции, когда в середине прошлого века туда проник из Америки одни из паразитарных грибов — оидиум, нанесший большой ущерб виноградникам. Опыливание виноградников серой помогло приостановить распространение этой опасной болезни.
Однако сера оказалась малоэффективной против другого, также проникшего с Американского континента паразита виноградников, называемого мильдью. И тогда против пего во Франции была предложена смесь, состоящая из медного купороса и негашеной извести, в которой действующим началом является медь. Подобная смесь, названная бордосской жидкостью, оказалась весьма эффективной не только против мильдью, но и других возбудителей болезней, благодаря чему была признана царицей фунгицидов. Так было положено начало промышленного производства фунгицидов с использованием для этих целей как неорганических веществ, так и сложных органических соединений.
С увеличением числа фунгицидов их стали классифицировать по биологическому действию, химическому составу, способу применения и ряду других важных признаков. В самом общем виде существующие сейчас фунгициды можно разделить на контактные и системные.
Первое поколение фунгицидов относится к контактным, для действия которых необходим контакт фунгицида с патогеном. Все они обладают широким спектром биологической активности. Они действуют не избирательно, а сразу на многие структуры клетки и разные звенья обмена веществ, являющиеся более или менее общими для всех живых организмов. Благодаря этому паразитарным грибам и бактериям довольно трудно приспособиться к контактным фунгицидам и многие из них используются в течение длительного времени. Примером является та же бордосская жидкость.
Однако все эти фунгициды вступают в контакт с паразитом только на поверхности растения. Ни один из них не может проникать в ткани растений в сколько-нибудь эффективных дозах. Кутикула, покрывающая поверхность листьев, стеблей, плодов, является для них непреодолимым барьером. Это и хорошо, и плохо. Хорошо потому, что, не проникнув в растение, они не оказывают на них токсическое действие. Но по этой же причине они являются эффективными лишь против паразитов, развивающихся на поверхности растения. И это плохо. Они могут оказаться эффективными и против паразитов, проникающих в растение, по лишь в том случае, если вступают
с ними в контакт еще до их проникновения в растительную ткань, когда они находятся на поверхности растения. Следовательно, против таких паразитов контактные фунгициды приходится применять еще до появления симптомов болезни. Но до встречи с паразитом фунгицид может разложиться, его может смыть дождь или сдуть ветер. Кроме того, и это весьма существенно, контактными фунгицидами надо покрывать всю поверхность растения, как верхнюю, так и нижнюю сторону листьев. В небольшом хозяйстве такая обработка растений хотя и является трудоемкой, по осуществима. На больших же площадях достичь такого равномерного нанесения фунгицида с помощью существующих технических средств практически невозможно.
Ученые начали создавать второе поколение пестицидов системного действия, т. е. соединения, которые способны проникать внутрь растения и перемещаться в нем. О возможности вводить в растения чужеродные вещества давно известно. Еще в XII в. пытались получать плоды
с новым ароматом, вкусом и цветом путем введения в просверленные в стволах отверстия пряностей и красителей.
13 конце 30-х годов нашего столетия на основе учения о передвижении веществ в растениях был создай первый системный инсектицид, а четверть века спустя — первый системный фунгицид.
Отличительная особенность системных фунгицидов состоит в том, что они способны проникать в растение, продвигаться по нему, убивать или подавлять в нем рост проникшего паразита, не повреждая при этом растительную клетку. Иными словами, мирно существовать с пей. Все они действуют избирательно, т. е. на определенное звено в системе обмена организма. Если контактные пестициды бьют бесприцельно, то системные поражают лишь определенную мишень. Так, одни из первых системных фунгицидов — бенлат, или беномил, связывается только с одним белком тубулином, а такие контактные фунгициды, как карбаматы, инактивируют более 20 белков-ферментов.
Следует, правда, признать, что действие системных фунгицидов все же еще мало изучено. Многими работами показано, что системные фунгициды в отличие от контактных оказывают влияние преимущественно на биосинтетические процессы, т. е. ведущие к получению нового вещества, необходимого для роста и сохранения организма. Контактные фунгициды в первую очередь вызывают нарушение структуры клетки.
Конечно, прицельный удар системного фунгицида является более точным и поэтому более сильным. Действительно, все системные фунгициды оказались более сильными по сравнению с контактными. Но в то же время и паразитам легче приспосабливаться к фунгицидам, которые бьют только по определенной мишени. Так оно и произошло. В результате вместо уничтоженных рас паразита стали появляться новые, еще более агрессивные, а взамен одних видов микроорганизмов начали усиленно развиваться другие. Знакомая ситуация. Возникли новые проблемы, и начался поиск путей их решения. Проблемы в общем уже знакомые, близкие к тем. которые возникали при использовании сортов растений с вертикальной и горизонтальной устойчивостью. Подобно сорту с вертикальной устойчивостью, системный фунгицид хотя и действует сильно, однако быстро теряет свою активность в силу адаптации к нему фитопатогенов. А подобно сорту с горизонтальной устойчивостью, действие контактного фунгицида хотя и менее сильно, но зато более продолжительно, поскольку к нему трудно адаптироваться. Поэтому как при выведении сортов необходимо заботиться о многообразии генов их устойчивости, так и при подборе фунгицидов нужно стремиться к многообразию их химического строения. Это может быть достигнуто разными путями. Во-первых, созданием фунгицидов в виде смесей из различных химических соединений, обладающих синергическим действием. Во-вторых, чередованием применения разных фунгицидов. Задача не простая, так как пока еще далеко не ясно, что с чем смешивать, в какой последовательности и что с чем чередовать.
Но даже системный фунгицид, каким бы избирательным действием он пи обладал, если он убивает или подавляет паразита, то может оказаться токсичным и для других организмов и, следовательно, потенциальным нарушителем экологического равновесия со всеми вытекающими отсюда отрицательными последствиями. Наряду с дальнейшим совершенствованием системных фунгицидов перед учеными встала новая задача — найти соединения, которые не были бы ни фунгитоксичными, ни фитотоксичными, а усиливали бы естественные механизмы устойчивости растений к болезням.
Сравнительно недавно удалось синтезировать соединения, которые, будучи слабо токсичными или вовсе нетоксичными для паразитов, тем не менее защищали от них растения. Механизм их действия пока не изучен, но поскольку они не токсичны для паразитов, то можно предположить, что они каким-то образом действуют на механизмы, контролирующие фитоиммунитет, хотя это и необязательно.
Некоторые фитопатологи предлагают па. звать соединения, действующие по принципу повышения устойчивости растений к болезням, системными терапевтантами. Но дело не в терминологии. Главное — найти вещества, которые действительно повышали бы устойчивость растений к болезням, не оказывая при этом отрицательного влияния на биосферу. Их поиск начат, и постепенно выявляются трудности, которые придется преодолеть.
ИММУНИЗАЦИЯ
Триумфом медицинской иммунологии является избавление человечества от многих весьма опасных в прошлом болезней. Достигнуто это путем искусственно привитого иммунитета. Достаточно упомянуть о ликвидации таким путем оспы, уносившей в прошлом сотни тысяч человеческих жизней. Вакцины созданы и против других тяжелых болезней. Иммунологи рассчитывают на новые вакцины, включая вакцины против рака. Ничего этого пока нет в активе фитоиммунологии. Нет «пока», но мы убеждены, что будет, причем будет в самом ближайшем будущем. Для подобного оптимизма сейчас уже имеются достаточные основания.
Перенесемся мысленно в не столь далекое прошлое, когда повсюду свирепствовала оспа. Из каждой сотни людей шестьдесят заболевали оспой, двадцать из них умирали, а оставшиеся сорок оставались на всю жизнь обезображенными. Древние китайцы и арабы давно знали, что люди, перенесшие оспу, редко заболевают ею вновь. Предполагается, что именно на Востоке впервые начали вырабатывать у людей искусственный иммунитет, заражая их веществом из гнойников больных и вызывая при этом несильный приступ болезни.
Однако прививание здоровому человеку вещества из оспенных язв больного было крайне опасной процедурой: само оспопрививание вызывало немалую смертность, а прививки могли стать причиной эпидемии.
Существовала и еще одна форма оспы — коровья. Люди ею почти не болели или болели в легкой форме. При этом бытовало поверье, что человек, переболевший коровьей оспой, никогда не заболевает настоящей. Этим заинтересовался английский врач из Глочертершира Эдвард Дженнер, который несколько лет своей жизни посвятил изучению этого явления. В 1798 г. он привил мальчику коровью оспу, а затем заразил его настоящей. Мальчик не заболел. Отсюда и возник термин вакцинация, поскольку инфекционный материал брался от коровы, а корова по-латыни — vacca.
И хотя Дженнер подарил человечеству способ предупреждать оспу, тем не менее его опыты не создали общего принципа защиты от болезней. Прошло еще 85 лет, прежде чем Луи Пастер открыл закон стимуляции иммунитета с помощью вакцин. Пастер разработал универсальный принцип: искусственный иммунитет можно создать, если в организм предварительно ввести ослабленный возбудитель той болезни, к которой нужно выработать невосприимчивость.
В последующие годы ученые добивались успеха, иммунизируя животный организм возбудителями той же болезни, ослабленными с помощью обработки теплом, светом, недостатком кислорода, культурами убитых микроорганизмов, токсинами микроорганизмов и, наконец, анатоксинами (обезвреженными токсинами, сохранившими свои антигенные группировки). Использовалась и пассивная иммунизация, в том случае когда в организм вводили готовые антитела, полученные от других животных.
С помощью этого арсенала средств удалось создать действенные вакцины против оспы, бешенства, сибирской язвы, дифтерии, полиомиелита и других опасных болезней. Практическая иммунология торжествовала.
Естественно, что фитоиммунология, которая к тому времени только зарождалась как наука, не могла оставаться в стороне от успехов медицины. И столь же естественно, что она пошла по пути, проложенному медиками. Исследователи вводили в растения, поливали их ослабленными культурами микроорганизмов, их авирулентными штаммами, либо экстрактами. Мнения ведущих фитоиммунологов того времени относительно возможности приобретенного иммунитета у растений расходились. Так, Овенс в своем обстоятельном трактате «Принципы растительной патологии» в 1928 г. писал: «Заметного прогресса в этом направлении пет, и сомнительно даже, будет ли он когда-нибудь в отношении растений, как это имеет место применительно к животному царству. Структура растений так отлична от животных, особенно в отношении сообщающейся системы, что кажется мало вероятным, чтобы когда-либо были достигнуты успехи в этом направлении»
[20].
Однако отдельные успехи все же были. Уже в начале века были проведены опыты, подтверждающие возможность иммунизации растений. Так, Г. Ноэль и Н. Бернар показали, что орхидеи после заражения их паразитарными грибами из рода Rhizoctonia приобретали иммунитет к повторным заражениям. Т. Боверн и О. Рай еще в 1901 г. применили вакцинацию бегонии, овса, фасоли и люпина как ослабленными культурами грибов и бактерий, так и экстрактами из этих культур и получили положительные результаты. Итальянские исследователи Д. Карбоне и С. Арнауди провели многочисленные опыты по иммунизации растений путем воздействия на них ослабленными культурами или экстрактами из микроорганизмов. На основании их исследований установлен факт усиления или образования заново иммунитета у растений.
В общем имелись отдельные примеры успешной иммунизации растений путем вакцинации как ослабленными культурами паразитарных грибов и бактерий, так и экстрактами. К сожалению, срок действия такой иммунизации, как правило, был очень невелик.
Но наряду с этим существовали противоположные результаты, свидетельствующие о неудачах иммунизации, и таких было большинство.
Положение было крайне неопределенным. Многие авторитеты резко возражали против попыток иммунизации растений. Но не Н. И. Вавилов, который писал, что возможность приобретенного иммунитета у растений в результате вакцинации и влияния паразитов может считаться доказанной. Неясна только природа такого иммунитета. Последнее, вероятно, и служило возможной причиной неудач. Ведь опыты по иммунизации
растений проводились вслепую, методом проб и ошибок. Никакой теории иммунитета не существовало. Было совершенно неясно, что именно в растениях следует иммунизировать и чем следует проводить иммунизацию. Простая аналогия с принципами иммунизации животных к успеху не приводила. Нужна была теория, вскрывающая отличия и единство животных и растений, чтобы с ее помощью вооружить исследователей методом иммунизации.
Каково же положение с иммунизацией растений к настоящему времени? Если в теоретических аспектах иммунизации наметился определенный прогресс, то в вопросах ее практической реализации исследователи недалеко ушли по сравнению с состоянием дел в начале нашего века. Пожалуй, единственным исключением из этого правила является метод вакцинации, предохраняющий от вирусных болезней.
Вакцинация, защищающая растения от вирусов, предполагает индуцирование в них вирусоустойчивости в ответ на обработку ослабленными штаммами вирусов. В вакцинированных растениях вирулентные штаммы вирусов развиваются медленнее, их распространение сдерживается, а вредоносность снижается. Считалось, что в вакцинированном растении происходит антагонистическое взаимодействие или интерференция между двумя вирусами, один из которых представляет собой предварительно введенный в растение вакцинный штамм, а другой — патогенный штамм, которым осуществляется заражение.
Наибольшие успехи достигнуты при вакцинации томатов вирусом табачной мозаики для борьбы с мозаичным заболеванием томатов в условиях закрытого грунта. Такого рода вакцинация уже сейчас успешно осуществляется в Голландии, Швеции, Дании, Англии, Канаде, США, Японии. В СССР вакцинация томатов против вирусов официально одобрена и рекомендована к внедрению МСХ СССР. Теоретически не отвергается возможность вакцинации и других культур. Уже появляются отдельные экспериментальные работы по использованию слабо-патогенных вирусов в борьбе с вирусными болезнями картофеля и огурцов.
Однако метод вакцинации ослабленными штаммами вирусов должен использоваться с соблюдением осторожности, поскольку не исключено, что в популяции вакцинного штамма могут появиться агрессивные формы, которые при благоприятных условиях могут поразить устойчивые сорта. Именно поэтому метод пока еще не выходит за пределы закрытого грунта.
О механизмах вакцинации пока еще известно мало. Существует несколько гипотез, ни одна из которых пока еще не может считаться окончательно доказанной. Одна из таких гипотез, так называемая гипотеза белкового пленения, состоит в предположении, что РНК патогенного вируса захватывается в белковую оболочку вакцинного штамма. Вторая — построена на конкурирующих антагонистических отношениях между двумя типами вирусов. Однако при такой трактовке сводится на нет активный защитный ответ клеток вакцинируемого растения. Поэтому другая часть гипотез рассматривает вакцинацию как активацию защитных функций самого хозяина. Здесь наиболее популярна возможность образования в вакцинированном растении иптерферопоподобпых белков, ингибирующих развитие вируса. Возможно также, что в вакцинированных растениях возрастает активность рибонуклеаз, которые разлагают РНК вирулентного вируса, образуя так называемый РНКазный барьер.
Другим примером успешной иммунизации, правда пока еще в порядке экспериментов в условиях закрытого грунта, является иммунизация растений против фитопатогенов с помощью самих фитопатогенов, которой в течение последних лет активно занимается американский фитоиммунолог Д. Куч. Для защиты растений он использует ослабленные культуры патогенов, непатогены данного растения, их авирулентные расы и, наконец, сами агрессивные патогены. В основном объектом его усилий являются растения из семейства тыквенных (огурцы, дыни, арбузы). Вот один из его типичных опытов.
Первый настоящий лист огурцов заражают патогенным штаммом Colletotrichum lagenarinm, вызывающим антракноз. Через 4 дня все вышерасположенные листья, а также листья, которые еще не раскрылись, становились защищенными от инфекции. Удаление листа иммунизатора на иммунизацию не влияло, по для ее достижения нужно время, в течение которого зараженный лист остается на растении. Можно срезать вышерасположенные иммунизированные листья, и они в срезанном состояния будут продолжать оставаться иммунизированными. Из пазушной почки иммунизированного растения вырастает побег, который после удаления и укоренения превращается в иммунизированное растение.
Сигнал об иммунизации в большей мере передается снизу вверх, чем сверху вниз. Если иммунизировать подвой, то ранее не иммунизированный привой окажется иммунизированным. Если же, наоборот, иммунизировать привой, то подвой иммунизированным не становится.
Кольцевание листа иммунизатора полностью прекращает иммунизацию, что позволяет предполагать, что иммунизирующий сигнал распространяется по флоэме. Одно иммунизирование защищает огурцы на 4–5 недель, а затем устойчивость постепенно теряется. Однако если провести вдогонку повторное иммунизирование, то период устойчивости удается продлить.
Чрезвычайно интересен установленный Кучем факт, что иммунизация становится невозможной, как только огурцы зацветают и начинают плодоносить. Предполагается, что цветение изменяет гормональный баланс, что препятствует иммунизации.
Первые опыты Куча проводились в теплицах, но с 1980 г. они были частично перенесены на опытные делянки. Однако к числу полевых опытов их причислить нельзя, во-первых, потому что иммунизирующее заражение проводилось в теплице, после чего растения высаживали в поле, а во-вторых, потому что результаты иммунизирования проверялись посредством искусственного заражения, а не естественного.
Поскольку в качестве иммунизаторов Куч использовал сами патогены, иногда авирулентные или ослабленные, а иногда жизнеспособные и вирулентные, то в адрес его опытов постоянно раздаются упреки, что они представляют собой опасность распространения той болезни, с помощью которой проводилась иммунизация. Пытаясь заменить патогены выделенными из них веществами, Куч испробовал в качестве иммунизаторов препараты клеточных стенок грибов и бактерий, их экстрацеллюлярные экстракты, эксудаты флоэмы, экстракты инфицированных и иммунизированных тканей, однако все опыты оказались безуспешными.
Американский фитоиммунолог и бактериолог Л. Секвейра, анализируя явления иммунизации растений, приводит ее следующие недостатки:
1) индуцирование устойчивости является энергопотребляющим процессом, поэтому иммунизация может приводить к уменьшению урожая;
2) иммунизация может приводить к накоплению в растительных тканях нежелательных метаболитов вторичного происхождения, обладающих токсическим действием;
3) если в качестве иммунизатора используется патогенный микроорганизм, то заражение им может вызвать эпифитотии;
4) если в качестве иммунизаторов используются индукторы паразита, то они должны быть низкомолекулярными, чтобы проникнуть в растительную ткань, тогда как большинство известных к настоящему времени индукторов обладают большой молекулярной массой
Из возбудителя фитофтороза картофеля нам удалось выделить иммунизатор системного действия — липогликопротеидный комплекс, который свободен от перечисленных недостатков. Комплекс принадлежит к числу элиситеров, или индукторов фитоалексинов и некрозообразования картофеля, в том случае, если используется в концентрациях 0,01 %, но не вызывает некроз, а сенсибилизирует растительные ткани, повышая их устойчивость к последующей встрече с инфекцией, если применяется в концентрациях 0,0005—0,001 % (мы уже рассматривали действие этого комплекса в главе «Сенсибилизация»).
Индуктором опрыскивали клубни картофеля перед посадкой, затем клубни высаживали в грунт на опытных делянках Белорусского НИИ картофелеводства и плодо-овощеводства. Первое, что удавалось наблюдать, — это большая высота выросших побегов и большее их число в кусте. Дело в том, что глазки необработанных контрольных клубней сильно страдают от ризоктониоза, который ведет к изреживанию побегов в кусте и их ослаблению. Обработка индуктором более чем в три раза уменьшает потери от ризоктониоза, в результате чего большее число глазков образуют ростки, которые вырастают в полноценные побеги. Выросшие растения примерно на 40–50 % меньше контрольных поражались фитофторозом и ранней сухой пятнистостью. Клубни нового урожая оказались на 2/3 меньше пораженными паршой и ризоктопиозом, как сразу после уборки, так и во время храпения. В результате этого потери клубней от болезней в период хранения уменьшились почти вдвое.
К весне эффект сенсибилизации постепенно затухал, и растение, выросшее из второго поколения клубней, мало чем отличалось по степени свей устойчивости от контрольных.
В целом тот уровень устойчивости, который наблюдался у обработанных индуктором растений, не только не уступал по своей эффективности защите при использовании фунгицидов (опрыскивание вегетирующих растений фунгицидом цинеб, а также предпосадочная обработка клубней фунгицидом ТМТД), но в отношении устойчивости к парше и ризоктопиозу превосходил ее. Учтите при этом, что опрыскивание фунгицидом, как правило, приходится проводить в зависимости от прогноза 3–4 раза в сезон. Подобные результаты получались в течение нескольких лет подряд. Таким образом, реальный эффект от предпосадочной иммунизации клубней выражался в следующем:
1) число побегов в кусте так же, как и высота побега, возрастало на 10 % из-за меньшего поражения картофеля ризоктопиозом;
2) вегетирующие растения вдвое меньше поражались фитофторозом и ранней сухой пятнистостью;
3) клубни второго поколения на 2/3 были меньше поражены паршой и ризоктопиозом;
4) прибавка урожая в зависимости от года составляла 12–27 %;
5) сенсибилизированные клубни второго поколения быстрее и интенсивнее залечивали механические поражения;
6) потери клубней во время хранения сокращались наполовину;
7) в силу меньшего поражения клубней второго и первого поколения ризоктопиозом улучшались их семенные качества;
8) сокращались расходы на обработку посевов (одна предпосадочная обработка вместо 3—4-кратного опрыскивания растений фунгицидами в поле);
9) предпосадочная обработка клубней вместо вегетационной позволяла избежать поранения растений, а следовательно, их заражения вирусами, которые проникают в растения через пораненные участки.
Близкие результаты были получены и при предпосевной обработке семян томатов тем же индуктором (опыты на делянках БНИИКПО). Замачивание семян томатов в суспензии индуктора (0,0005 %) сократило поражение листьев фитофторозом, макроспориозом, септориозом и черной бактериальной пятнистостью на 34 %, а поражение плодов фитофторозом, макроспориозом и другими гнилями — на 65 %. В результате прибавка урожая составила около 30 %. Примерно тот же самый и даже несколько меньший эффект достигался при предпосадочной обработке растений фунгицидом ТМТД (3,5 %) наряду с 4-кратным опрыскиванием ботвы томатов цинебом.
Индуцированная устойчивость томатов почти в 2 раза превосходила эффект от применения фунгицида — хомицина (при 4-кратном опрыскивании вегетирующих растений 0,5 %-ной суспензией). Прибавка урожая в этом последнем случае составила 22 % в вариантах опытов с индуктором и только 6 % — при применении фунгицида.
Однако самый лучший эффект давало сочетание замачивания семян томатов в растворе индуктора с однократной обработкой им ботвы. В этом случае вегетирующие растения и выросший из них урожай оказывались защищенными на 84 %, что более чем вдвое превосходило эффект от применения фунгицидов.
Хорошие результаты были получены при использовании элиситера из возбудителя серой гнили (Botrytis cinerеа) для индуцирования устойчивости сахарной свеклы к болезням (совместные исследования с НИИ сахарной промышленности). Семена свеклы замачивали в растворе элиситера в течение 2 ч, высушивали и высевали на опытных участках размером 3 га.
Сразу же удалось заметить снижение повреждения проростков сахарной свеклы корнеедом — комплексным заболеванием свеклы, которое могут вызвать около 100 видов грибов и бактерий. Чаще всего это грибы из родов Fusarium, Pythium, Aphanomyces, Phoma, Rhizoctonia, Penicillium. Поражение и гибель проростков при их подземном развитии приводит к недружности и изреженности всходов. Из-за этого возникают редкие посевы, отстающие в развитии растения, в результате чего снижается урожай и ухудшается его качество.
Значительно выше оказались урожайность и технологические качества опытной свеклы. Так, основной показатель — расчетный выход сахара — составлял у обработанной индуктором свеклы 105–107 %.
Фитопатологический анализ корней сахарной свеклы, проведенный спустя 75 суток после закладки на хранение, показал, что такая свекла значительно более устойчива, чем выращенная из необработанных семян. Опытная свекла имела на 52 % меньше загнивших корнеплодов, а гнилая масса составляла только 11 % по сравнению с контрольной. Такая свекла и после хранения имела значительно лучшие технологические свойства: выход сахара из нее был на 4,8 % выше, чем из необработанной.
Неоднократно предпринимались попытки использовать сами фитоалексины или их аналоги в качестве естественных пестицидов для борьбы с болезнями растений. Канадские фитопатологи обнаружили, что фитофтороустойчивость томатов и картофеля заметно возрастала после их обработки фитоалексинами перца (капсидиолом) и картофеля (ришитином). Однако эта работа не имеет перспективы, так как для защиты необходимы высокие концентрации препаратов.
Попытки химически синтезировать фитоалексины успехом не увенчались. Их синтез достаточно сложен, так как многие из них имеют асимметрический углеродный атом и лабильные функциональные группировки. Английские исследователи показали, что фитоалексины бобовых — птерокарпаны вообще не эффективны как фунгициды. С нашей точки зрения, такой путь защиты вообще малоперспективен, так как вновь созданные пестициды вряд ли окажутся менее опасными для окружающей среды, чем уже существующие. К тому же токсичность фитоалексинов для грибов меньше, чем токсичность фунгицидов, тогда как для людей они ничуть не менее токсичны. Синтезировать фунгициды по прообразу индуцирования антибиотиков означает «рассекретить» оружие растений и облегчить тем самым адаптацию к ним паразитов.
Особенно важным представляется тот факт, что с помощью иммунизации достигается стимулирование в растениях свойства неспецифической или общей устойчивости. В какой-то мере она напоминает полевую, полигонную или относительную устойчивость, с той, однако, разницей, что последняя является генетически определенным признаком того или иного сорта, тогда как мы регулируем свойство устойчивости в пределах онтогенеза данного генотипа. В результате обработки индуктором изменяется не геном, а только его функционирование или, вернее, меняется уровень дерепрессии генов. Иными словами, часть генов контрольного необработанного растения функционирует не полностью, а часть генов вообще зарепрессирована. Обработка индуктором снимает часть репрессоров и тем самым как бы раскрепощает гены растения. Эффективная защита хозяина требует правильно выбранного индуктора в строго определенной концентрации, для того чтобы достигнуть нужного уровня дерепрессии клеточного генома. Индуктор при этом как бы является ключом к замку, открывающим устойчивость растения-хозяина и освобождающим его защитные силы (рис. 14 см. вклейку). Не исключено, что индуктор у паразита также зарепрессирован и освобождается только при взаимодействии с растением. Поэтому происходит взаимное индуцирование, в ходе которого растение высвобождает индуктор паразита, а последний дерепрессирует образование фитоалексинов у растения. Этот факт служит лишним доказательством того, что всякому виду и сорту растения, даже по несущему гены устойчивости, свойственна потенциальная способность к защите. Найти тот способ, с помощью которого можно индуцировать эту потенцию к жизни, и является задачей индукции.
Хотя иммунизация обеспечивает не абсолютный, ио только относительный уровень устойчивости, зато он проявляется по отношению ко всем расам паразитов, а также ко многим другим видам фитопатогеиов. Вспомните, обработка картофеля липогликопротеидным комплексом защищает его от фитофтороза, ризбктониоза, ранней сухой пятнистости, парши и других болезней. Иммунизированная свекла оказывается защищенной от комплекса болезней, вызывающих корневые и кагатные гнили. В то же время именно индуцирование неспецифической устойчивости позволяет иммунизировать все сорта растений, независимо от присутствия у них генов вертикальной болезнеустойчивости.
Подобный уровень устойчивости вряд ли сможет преодолеваться паразитами именно в силу своего неспецифического действия, т. е. отсутствия определенной мишени, к которой мог бы легко адаптироваться паразит. Ведь известно, что полигонная устойчивость не преодолевается патогенами, хотя ее уровень относительно невелик. Именно поэтому полевая, или полигенная, устойчивость и представляет наибольшую перспективу для селекционеров, занимающихся выведением болезнеустойчивых сортов.
Иными словами, иммунизация, о которой шла речь, сочетает в себе достоинства контактных и системных пестицидов и лишена недостатков, присущих каждому из них. Подобно контактным пестицидам, она неспецифичиа в действии и поэтому позволяет избежать привыкания фитопатогенов, подобно системным — обладает действием, распространяющимся по всему растению и настигающим паразита, сколь глубоко бы он ни проник в ткани.
Нам представляется, что иммунизация растений на настоящем этапе может дополнить арсенал современных средств защиты растений. Обойтись сейчас без пестицидов сельское хозяйство не в состоянии. Поэтому иммунизация ни в коей мере не отменяет химических средств защиты растений, так же как иммунизация животных не отменяет использования антибиотиков — этих, по образному выражению Куча, «пестицидов животного царства». Уже сейчас иммунизация позволяет существенно сократить затраты пестицидов на обработку посевов: в сочетании с иммунизацией можно использовать одну обработку посевов фунгицидами вместо четырех и даже пяти. Вряд ли имеет смысл доказывать, насколько важно уменьшить число обработок растений фунгицидами при одновременном повышении эффективности их действия.
Значение выполненных исследований состоит в том, что ими показана принципиальная возможность иммунизации растений с помощью биогенных индукторов защитных реакций и намечены пути их использования. Так как применение биогенных индукторов основано на тех же принципах, что и самозащита растений, то можно ожидать, что она не создаст угрозы нарушения экологического равновесия в биосфере. Обработка индуктором не вызывает у растений образования токсических фитоалексинов, а лишь подготавливает их к возможной встрече с патогеном. И все же, как и при любом другом вмешательстве в природу, следует учитывать не только видимый эффект, но и его возможные отрицательные последствия в будущем. Явление иммунизации нуждается в детальном изучении, чего можно достигнуть только при объединении усилий специалистов разных профилей.
* * *
Интегрированная система защиты растений в какой-то степени напоминает множественность защитных реакций, с помощью которых растение само себя защищает от фитопатогенов в природных условиях. И так же, как и при естественной защите растений, отдельные мероприятия интегрированной защиты часто действуют синергически, усиливая и взанмодополняя друг друга.
Вот те преимущества, которые имеет интегрированная система защиты растений.
1. Все слагаемые этой системы взаимодополняют и усиливают друг друга, в результате каждое из них приходится использовать в меньших дозах и с меньшей частотой.
2. Использование устойчивых или иммунизированных сортов растений позволяет заметно снизить число обработок пестицидами. К примеру, использовать вместо 3–4 обработок в сезон только одну или две.
3. Сокращение числа обработок пестицидами не может не иметь значения, для того чтобы избежать нарушения того экологического равновесия в биосфере, которое неизбежно возникает под воздействием обработок химикатами.
4. Интегрированная система защиты позволит выводить сорта, устойчивые к более широкому кругу возбудителей болезней и вредителей.
5. Использование устойчивых или иммунизированных сортов в интегрированных системах защиты растений удешевит борьбу с болезнями, так как применение устойчивых сортов значительно менее дорогостоящее мероприятие, чем ежегодное производство и использование пестицидов.
ВСЕ ВЫРАЩЕННОЕ СОХРАНИТЬ
Издавна в народе говорят: «Не тот урожай, что на полях, а тот, что в закромах». В условиях индустриального сельского хозяйства вовремя убрать и хорошо сохранить урожай порою бывает даже труднее, чем его получить. Многие традиционные способы хранения, вполне соответствующие требованиям небольшого хозяйства, не позволяют сохранить клубни картофеля или корнеплоды свеклы с механическими повреждениями, в какой-то мере неизбежными при уборке урожая с помощью машин; Не рассчитаны эти способы на хранение больших количеств продукции, убранной в ненастную погоду, чего также трудно избежать в крупном хозяйстве.
Особенно велики потери при храпении картофеля, овощей и плодов, а их качество после храпения часто оставляет желать много лучшего. Судя по данным мировой статистики, примерно 15 % мирового урожая уничтожают болезни и вредители во время хранения.
Еще древние врачи наделяли плоды и овощи чудодейственной силой и рекомендовали их для лечения многих болезней. Наоборот, в 50—70-х годах прошлого столетия значение плодов и овощей в питании почти полностью игнорировалось, так как считалось, что ценность всякого пищевого продукта определяется прежде всего его калорийностью. Поскольку плоды и овощи в большинстве обладают малой калорийностью, они рассматривались лишь как вкусовые вещества. И только после установления роли витаминов, минеральных солей, органических кислот и многих других соединений, которыми богаты плоды и овощи, стала ясной их исключительно важная роль для поддержания нормальной жизни человека, его работоспособности и долголетия.
Все это несомненно интересно и важно, но какое, собственно, оно имеет отношение к фитоиммунологии, которой посвящена настоящая книга? Дело в том, что основой всех практических мероприятий по хранению овощей и плодов является прежде всего максимальное использование присущей растительным тканям устойчивости к фитопатогенным микроорганизмам, фунгициды при хранении не применяются. Игнорирование этого условия неизбежно приводило к серьезным просчетам. И, наоборот, на основе подробного изучения физиологических и биохимических процессов, происходящих в растительных тканях на протяжении всего периода хранения урожая, удалось разработать весьма эффективные способы защиты его от болезней и другого вида потерь применительно к требованиям, предъявляемым производственной практикой.
Конечно, свести потери к нулю невозможно, поскольку приходится хранить биологические объекты, в которых при хранении активно продолжаются сложные физиологические и биохимические процессы.
Сложность проблемы состоит в том, что нередко забота о предотвращении одного источника потерь приводит к увеличению потерь другого рода. Например, весьма падежные способы предупреждения прорастания клубней при хранении могут вызвать поражения их возбудителями инфекционных болезней. Эффективные способы защиты плодов от инфекционных болезней подчас вызывают в растительных тканях функциональные расстройства, внешне проявляющиеся в побурении плодов, потере свойственной им консистенции. Совершенно очевидно, что решить проблему хранения урожая можно не с помощью какого-то одного метода, каким бы эффективным он ни был, а введением целой системы организационных и технических мероприятий, направленных одновременно против всех видов потерь. В общем та же интеграция, в основу которой должен быть положен иммунологический контроль за жизнью отделенных от материнского растения органов — клубней, корнеплодов, плодов, луковиц.
Тем не менее время от времени появляются сенсационные сообщения о возможности решения проблемы хранения если не целиком, то в основном с помощью одного способа, применяемого чуть ли не в любых условиях. Это наносит серьезный ущерб делу. Во-первых, отвлекает силы и средства на проведение разного рода проверочных опытов. Во-вторых, и это не менее важно, создает ложное представление о проблеме в целом и необоснованно упрощает пути ее решения.
Несостоявшаяся революция
Крупные успехи, достигнутые в области ядерной физики, стимулировали проведение многочисленных исследований по изучению возможности защиты различных пищевых продуктов, в том числе плодов и овощей, от болезней с помощью ионизирующей радиации в виде радиоактивных изотопов (главным образом Со
6») и ускоренных электронов. Характерной особенностью ионизирующих излучений (γ-лучи, Х-лучи) является их способность превращать атомы и молекулы веществ в электрически заряженные частицы — ионы. Поглощение даже сравнительно больших доз ионизирующих излучений практически не вызывает повышение температуры. В этом состоит одна из специфических особенностей ионизирующей радиации, очень важной для использования ее с целью сохранения пищевых продуктов.
Под действием ионизирующей радиации в первую очередь изменениям подвергается вода, являющаяся главной составной частью живых организмов, в частности плодов и овощей. Происходит радиолиз воды — возникновение свободных радикалов. В таком состоянии они могут свободно существовать лишь в течение очень короткого срока времени, измеряемого миллионными долями секунды. Но все свободные радикалы химически весьма активны. Поэтому даже за короткий срок они вступают в самые различные реакции. Свободные радикалы легко реагируют с растворенными в воде веществами, в результате чего усиливаются процессы окисления, распада сложных органических веществ до более простых соединений, образования новых ранее отсутствующих соединений.
Разные организмы, органы, ткани, различные звенья обмена веществ и даже отдельные вещества неодинаково реагируют на действие ионизирующей радиации. Некоторые из них являются очень радиочувствительными, тогда как другие обладают весьма высокой радиоустойчивостью.
Клубни картофеля полностью теряют способность к прорастанию при дозе 10 крад; многие вредители зерна погибают при дозе 30 крад; а некоторые насекомые выдерживают дозу 300 крад. Для уменьшения количества микроорганизмов в 10 раз требуется для одних видов 25 крад, а для других — 400 крад. Сухой чистый крахмал не изменяется при дозе 300 крад, а в клубнях картофеля на него влияет уже доза 3 крад.
Лица, малознакомые с действием ионизирующей радиации, нередко вообще возражают против облучения пищевых продуктов, так как ошибочно полагают, что радиоактивное облучение может привести к таким же отрицательным последствиям, как радиоактивное заражение. Между тем это совершенно различные явления. Кроме того, облучение продуктов осуществлялось лишь при такой дозе ионизирующей радиации, при которой наличие в них наведенной радиоактивности полностью исключается.
Опасность можно ожидать с другой стороны. Поскольку облучение влечет за собой изменения в химическом составе продуктов, нельзя игнорировать возможность возникновения различного рода токсичных, канцерогенных и мутагенных веществ. Поэтому этим вопросам уделяется особое внимание. Судя по результатам длительных исследований в СССР, США, Канаде, Англии, Франции и других странах, во всех испытанных продуктах, облученных при рекомендованных дозах, не было обнаружено вредных веществ. На этом основании в ряде стран органы здравоохранения выдали разрешение на использование в пищу некоторых облученных продуктов. В СССР было, например, выдано разрешение на облучение картофеля с целью предупреждения его прорастания при хранении и на облучение зерна с целью уничтожения вредителей (дезинсекцию). Выдано было также разрешение на облучение плодов с целью уничтожения большей части содержащихся на их поверхности фитопатогенных микроорганизмов и защиты тем самым от инфекционных болезней — основного источника потерь при хранении.
После первых же экспериментов уже стали раздаваться голоса, особенно в США, о предстоящей революции в области храпения пищевых продуктов. Однако революции не произошло, во многих лабораториях исследования были свернуты, а их результаты оказались весьма поучительными. Так, было показано, что облучение плодов при дозе 200 крад позволяет сразу же снизить количество микроорганизмов, содержащихся на их поверхности, в 400 и даже в 1000 раз. Более низкие дозы малоэффективны, а более высокие вызывают ухудшение внешнего вида плодов. Однако уже через 5—10 дней число микроорганизмов на поверхности плодов вновь возрастает почти до исходного уровня, и тем быстрее, чем выше температура хранения. Основная причина этого явления — реактивация, восстановление жизнедеятельности микроорганизмов, не погибших, по лишь «нокаутированных» при облучении. Облученные плоды начинают поражаться даже теми микроорганизмами, которые, хотя и присутствуют на их поверхности, но в обычных условиях не причиняют им вреда. Объясняется это тем, что облучение вызывает серьезные повреждения в иммунной системе растительных тканей, вследствие чего они легче поражаются болезнями. Эти повреждения выражаются в ослаблении способности тканей образовывать раневую перидерму в ответ на механические поранения и синтезировать фитоалексины в ответ на контакт с фитопатогенными микроорганизмами. Облучение вызывает повреждение отдельных структур клетки, в частности ее энергетических центров — митохондрий. В результате происходит разобщение процессов окисления и фосфорилирования, которое состоит в том, что выделяющаяся в процессах окисления энергия рассеивается, а не запасается в особых соединениях (АТФ) для последующего использования на синтетические процессы. Кроме того, значительную часть энергии клетка вынуждена отвлекать от защитных реакций против паразита на репарацию своих собственных повреждений, вызываемых облучением.
Все это показывает, к каким отрицательным последствиям может привести стремление защитить урожай при хранении путем полного уничтожения самих возбудителей болезней. Собственно, такую задачу даже и нельзя ставить. Нельзя забывать, что паразитизм в биологии — явление нормальное. Поэтому если пытаться полностью уничтожить паразитов, поражающих урожай при храпении, то в результате можно лишь создать благоприятные условия для искусственного отбора среди них наиболее агрессивных форм, защитить урожай от которых станет уже во много раз труднее. Именно поэтому современная стратегия борьбы с болезнями растений основывается на интеграции всех доступных методов регулирования численности популяции паразитарных микроорганизмов при максимальном использовании защитных сил самих растений. Еще больше оснований руководствоваться этим принципом для защиты от инфекционных болезней хранящихся картофеля, овощей, плодов, сахарной свеклы. Ведь поскольку они являются органами, отделенными от материнского растения, в которых развиваются процессы старения, они становятся гораздо чувствительнее к антимикробным воздействиям, чем вегетирующее растение. Не случайно ни один фунгицид не нашел сколько-нибудь широкого применения при храпении картофеля, овощей и плодов, тогда как при их выращивании сейчас без фунгицидов обойтись нельзя.
Сказанное вовсе не означает полную бесперспективность поиска средств антимикробного воздействия при хранении клубней, корнеплодов и других запасающих органов растений, в которых активно продолжаются биохимические процессы. Интересными являются исследования по использованию для этих целей пирокатехина — соединения фенольной природы. Обработка свеклы перед закладкой на хранение 0,5 %-пым водным раствором пирокатехина способствовала одновременному решению нескольких задач. Во-первых, более интенсивному заживлению механических поранений. Во-вторых, меньшему поражению свеклы болезнями. В-третьих, сокращению потерь свеклой сахаров в акте дыхания. Хотя пирокатехин в такой концентрации обладал некоторым фунгитоксическим действием, по одновременно он выполняет и роль индуктора защитных реакций.
Все это лишний раз убеждает в том, что генеральной линией в защите урожая от болезней является максимальное использование устойчивости к болезням самих растительных тканей при одновременном подавлении жизнедеятельности фитопатогенных микроорганизмов. А это может быть достигнуто лишь на основе детального изучения механизмов устойчивости и разработки эффективных путей управления ими.
В воздушном потоке
В соответствующем разделе книги мы уже писали об активации окислительных и энергетических процессов в пораненных тканях растений. На месте механического поранения образуется раневая перидерма, представляющая собой как механический, так и химический барьер, препятствующий проникновению инфекции.
Для успешного образования раневой перидермы нужны строго определенные условия внешней среды: температура от 10 до 20 °C, относительная влажность воздуха 80–95 % в содержание кислорода не менее 15 %. Результаты выполненных исследований послужили биологической основой метода хранения картофеля в условиях активного вентилирования, при котором воздух с определенной температурой, влажностью и скоростью продувается через всю массу картофеля и получает доступ к каждому клубню. Последнее чрезвычайно важно, поскольку нормальному доступу кислорода в клубень препятствует образующаяся на нем пленка из воды, углекислого газа и других продуктов жизнедеятельности. Для того чтобы ее удалить, нужна активная вентиляция, одной естественной вентиляции здесь явно недостаточно. При неблагоприятных внешних условиях иммунная система растений не в состоянии активно функционировать. Иммунная система, защищающая организм, сама нуждается в защите, т. е. в создании оптимальных условий для ее деятельности. Создавая их, человек помогает наиболее полной реализации потенциальных возможностей иммунитета.
Нужно сказать, что активное вентилирование уже давно применялось для просушивания зерна и удаления из пего тепла, которое выделяется при дыхании и приводит к самосогреванию. В какой-то мере это необходимо и при хранении картофеля. Однако основная цель вентилирования картофеля состоит в интенсификции у механически пораненных клубней защитных реакций, приводящих к быстрому и падежному залечиванию механических поранений.
Для этого необходим иной режим вентилирования на различных этапах хранения в зависимости от физиологического состояния клубней. Вначале режим вентилирования рассчитан на успешное залечивание механических поранений, тогда как потом — на поддержание способности клубней продуцировать в случае инфицирования защитные вещества.
Проведенные исследования и передовой производственный опыт показали, что меньшие потери при хранении картофеля в условиях активного вентилирования позволят почти на треть увеличить емкость картофелехранилищ.
Активное вентилирование оказалось также эффективным и при хранении овощей — капусты, свеклы, моркови, репчатого лука, и оно широко внедряется в практику.
На страже покоя
Клубни картофеля, отделенные от материнского растения, определенный период не способны прорастать, даже при самых, благоприятных условиях. Они не прорастают потому, что находятся в состоянии глубокого естественного покоя. Как только период покоя заканчивается, клубни начинают настолько сильно прорастать, что для их сохранения приходится прибегать к искусственному продлению периода покоя. То же происходит и при храпении репчатого лука, моркови, свеклы, хотя период покоя у них менее четко выражен. Способность пребывать в состоянии покоя выработалась у растений в процессе эволюции как важное приспособление к неблагоприятным условиям для роста.
Хорошо известно, что в зонах умеренного климата с наступлением холодов деревья сбрасывают листья и рост побегов прекращается до весны. В степях и пустынях у многих травянистых растений после бурного роста и цветения отмирают листья и стебли; и растения переживают засушливое лето в виде клубней, луковиц, корневищ. Даже и у растущих растений отдельные органы и ткани до определенного момента находятся в состоянии покоя. Например, спящие пазушные почки древесных растений трогаются в рост лишь тогда, когда повреждается верхушечная почка.
В тропиках долгое время не удавалось получать урожай яблок, так как яблони, как и другие растения умеренной зоны, приспособились пребывать в состоянии покоя под влиянием низких температур, которых нет в тропиках. Опыт последних лет показал, что и вблизи экватора можно получить товарное производство яблок. Нанесение деревьям механических поранений выводит их из состояния покоя, так же как и холод. Поэтому начиная с годовалого возраста яблони в тропиках подрезают и срывают листья. Их продолжают срывать и в дальнейшем после каждого сбора. Подобная работа является очень трудоемкой. Но, по свидетельству плодоводов, качество получаемых плодов вполне удовлетворительное.
К. А. Тимирязев в своих замечательных лекциях о жизни растений, прочитанных 100 лет тому назад, отмечал, что пи одно явление в жизни растений не обращало на себя так много внимания, как пробуждение растений от покоя. С тех пор сделано немало, чтобы приподнять завесу над этой тайной природы. И хотя многое остается еще неясным, в ряде случаев уже удается найти новые подходы для управления, состоянием покоя.
Преждевременное прорастание является серьезным источником потерь при храпении картофеля и некоторых овощей. Собственно, в состоянии покоя пребывают лишь меристематические ткани, на долго которых в клубнях картофеля, например, приходится лишь 1 % их массы. Это так называемые «глазки»..
Клубни картофеля в силу высокого содержания в них воды и питательных веществ особенно уязвимы для фитопатогенных микроорганизмов. Причем особенно уязвимы меристематические ткани, те самые, которые находятся в состоянии покоя. Они могут позволить себе пребывать в покое только потому, что находятся под иммунологической охраной паренхимных тканей клубней.
Именно тогда, когда меристематические глазки «спят», паренхимные ткани клубня стоят на их страже: они обладают особенно высокой способностью быстро и интенсивно образовывать фитоалексины, отвечать реакцией СВЧ в ответ на внедрение патогена, активно образовывать раневую перидерму. Клубни тех сортов картофеля, которые обладают более длительным периодом покоя, характеризуются и более продолжительной устойчивостью к болезням. Как только период покоя у меристем: заканчивается, они могут прорастать и давать, начало новому растению, т. е. выполнять возложенную на них природой функцию, устойчивость паренхимных тканей быстро падает. Отсюда следует: важный практический вывод: сумейте продлить период покоя, и вы сумеете продлить период устойчивости и тем самым одновременно решить две задачи: задержать преждевременное прорастание клубней и защитить их от болезней. Причем речь идет именно о продлении периода, покоя, а не об искусственном подавлении прорастания клубней, поскольку это совершенно разные вещи.
Дело в том, что большинство методов, препятствующих прорастанию, основало на его необратимом подавлении. Примером этому является уже упомянутое γ-облучение, под действием которого деградируют точки роста у клубней, нарушается их структурная и функциональная целостность, а следовательно, и иммунологический контроль. Такие клубни мало синтезируют фитоалексинов и слабо образуют перидерму. Выходит, что методы, подавляющие прорастание клубней и тем самым предохраняющие их от одного вида потерь, как правило, нарушают свойство их естественной устойчивости, из-за чего возрастают потери другого рода.
Способ продления периода покоя для предупреждения прорастания принципиально отличается от подавления прорастания тем, что иммунологический контроль клубней в этом случае не только сохраняется, но продолжает оставаться на высоком уровне. Но для того чтобы продлить покой, нужно знать механизмы, его регулирующие. Наиболее изученными механизмами, регулирующими покой и переход к активному росту, являются фитогормоны. В клубнях картофеля обнаружены все известные фитогормоны, причем в состоянии покоя ведущую роль играет абсцизовая кислота. Это позволяет логически продолжить ту формулу, которая уже была приведена: если хотите продлить период покоя, а следовательно, и устойчивости, то сумейте индуцировать в клубнях образование фитогормона либо найдите способ поддерживать его содержание на нужном уровне. Индуктором образования абсцизовой кислоты оказался этилен, физиологическая активность которого была открыта более 80 лет тому назад Д. Н. Нелюбовым. Новые возможности для применения этилена как фитогормона появились сейчас благодаря синтезу его продуцентов, выпускаемых под разными названиями: этрел, гидрел, дигидрел, компазан. Под влиянием обработки гидрелом в картофеле индуцировалось образование абсцизовой кислоты, содержание которой оставалось на довольно высоком уровне. После обработки гидрелом клубни картофеля не прорастали даже после весьма длительного хранения. Но как только содержание абсцизовой кислоты снижалось и период покоя заканчивался, клубни начинали нормально прорастать и могли быть с успехом использованы для семенных целей.
Так, теоретическая разработка природы состояния покоя, проведенная в нашей лаборатории Н. П. Кораблевой, позволила создать метод предупреждения прорастания клубней без нарушения их иммунологического контроля.
Применение продуцентов этилена оказалось столь же эффективным и для сокращения потерь при храпении репчатого лука и корнеплодов.
Старение есть не столько прожитое, сколько нажитое
Согласно ранее существовавшим представлениям, старение всех организмов наступало вследствие истощения у них энергетических и пластических ресурсов. Поэтому основное внимание исследователей в области храпения плодов долгое время было сосредоточено на использовании возможно более низких температур, для того чтобы замедлить дыхание плодов и подавить деятельность фитопатогенных микроорганизмов. Но сейчас уже хорошо известно, что старение плодов, как и других микроорганизмов, связано не столько с истощением их ресурсов, сколько с накоплением различного рода токсических веществ, которые в молодых плодах вовлекаются в обмен. К примеру, после 9-месячного храпения некоторых сортов яблок в холодильниках содержание сахаров в них уменьшилось лишь на 30 %, тогда как содержание спирта возросло в 2 раза, ацетальдегида — в 4 раза, а некоторых перекисей — в 10 и более раз. В результате способность яблок продуцировать фитоалексины падала в 4–5 раз. Одновременно развивались неинфекционные болезни, внешне проявляющиеся в патологическом побурении плодов.
Уже более 50 лет изучается обнаруженный при созревании многих видов плодов кратковременный подъем дыхания, названный климактерическим, который соответствует кульминации процессов созревания, вслед за чем наступает перезревание. Климактерическому подъему дыхания предшествует биосинтез этилена — основного гормона созревопия, который индуцирует ряд ферментов, характерных для процесса старения. Одним из таких ферментов является малатдегидрогенеза декарбоксилирующая, названная ферментом старения. Его максимальное активирование совпадает с пиком климактерического подъема дыхания. Фермент декарбоксилирует (освобождает СО
2) яблочную кислоту, превращая ее в пировиноградную, которая также декарбоксилируется, образуя ацетальдегид. Ацетальдегид и другие токсические продукты, накапливающиеся в ткани, нарушают структуру мембраны вакуоли, в результате чего содержащиеся в ней полифенолы проникают в цитоплазму, где и окисляются полифенолоксидазой, вызывая побурение тканей.
В плодах на последних этапах роста обнаружен сесквитерпеноидный углеводород — фарнезен. Будучи соединением с двумя сопряженными связями, фарнезен легко подвергается перекисному окислению с образованием перекисей, гидроперекисей и свободных радикалов. Имеется большое число работ о накоплении продуктов перекисного окисления липидов в самых различных стареющих организмах, в том числе и у растений.
Из вышесказанного вытекают пути регулирования старения плодов. Именно регулирования, а не подавления, поскольку там, где имеет место только подавление, нарушается иммунологический контроль, что зачастую превращает, победу в поражение. Если старение характеризуется усилением окислительных процессов и повышенным выделением СО
2, то пониженно содержания кислорода в среде задержит процессы перекисного окисления липидов и биосинтез этилена, а повышение содержания углекислого газа по законам обратной связи затормозит процессы декарбоксилирования яблочной и пировиноградной кислот, а следовательно, накопление в клетке ацетальдегида и спирта. Тем самым отдаляется наступление климактерикса и старения.
Другой путь избежания накопления в тканях плодов высокотоксических гидроперекисей основан на применении антиоксидантов, действующих по тому же принципу, как и природные антиоксиданты, содержание которых в растительных клетках по мере старения уменьшается. Хорошие результаты были получены при применении антиоксиданта — дилудииа, который тормозит окисление фарнезена и накопление его перекисных продуктов и тем самым позволяет избежать побурения яблок. Теоретические основы старения плодов и способов его регулирования успешно разрабатываются в нашей лаборатории Е. Г. Сальковой.
Здесь приводятся примеры храпения клубней и плодов, уже отделенных от материнского растения, не только потому, что они являются хорошей биологической моделью для изучения иммунологического контроля при покое и старении, по и потому, что сохранить их не менее трудно, чем получить.
Весьма актуальным является дальнейшее более разностороннее изучение периода покоя растений и их перехода к активному росту, а также механизмов старения и связанного с ним возникновения функциональных расстройств и ослабления иммунной системы, создающей угрозу поражения болезнями. Такие исследования могут открыть новые подходы для защиты урожая от потерь.
ЗАКЛЮЧЕНИЕ
Наша книга подходит к концу. Вместе с читателем мы проделали большой и нелегкий путь. При этом, хотя мы и стремились к систематичности и равномерности изложения материала, все же невольно останавливались на том, что нас более всего занимало. А занимали нас те проблемы, над которыми на протяжении многих лет работает коллектив лаборатории иммунитета растений Института биохимии им. А. Н. Баха АН СССР.
В своих устремлениях мы забегали даже туда, где почва еще не отвердела и движение вперед пока затруднительно. Наша книга не изобилует примерами, поскольку большое число их, по нашему мнению, нарушает стройность изложения. Основная мысль, которую мы стремились проводить на протяжении всей книги, — это необходимость познания явлений природы, с тем чтобы следовать ее же законам в реализации на практике своих достижений. Иными словами, познавать сущность явления, чтобы его использовать. При этом мы руководствовались мудрым правилом: «пет и не может быть ничего практичнее, чем хорошая теория». Примеров здесь может быть сколько угодно.
Если для успешного протекания раневых реакций нужна повышенная температура, влажность и кислород, то активное вентилирование хранящихся плодов и овощей воздухом определенных параметров должно создавать условия для успешного залечивания поранений.
Если продление периода покоя клубней картофеля удлиняет период их устойчивости, то поддержание на определенном уровне фитогормонов, контролирующих покой, должно защитить картофель как от прорастания, так и от поражения фитопатогенными микроорганизмами.
Если включение защитных реакций растений зависит от распознавания элиситеров фитопатогенов, то задача заключается в том, чтобы отыскать у паразита такой элиситер. Если же природа позаботилась о том, чтобы заблокировать элиситер соответствующим супрессорой, то следует обнаружить не только элиситер, но и супрессор, отделить один от другого, а затем уже освобожденным элиситером в соответствующей концентрации иммунизировать растения.
Иммунизация растений с помощью элиситеров практически означает придание растению временного свойства горизонтальной устойчивости, которая с трудом преодолевается фитопатогенами.
Использование мультилинейных сортов основано на создании разнообразия генофонда устойчивости, что является характерным для дикорастущих растений.
Число таких примеров можно было бы увеличить. Причем мы говорим только о тех из них, которые перестали быть белыми пятнами в науке о болезнях растений. А сколько их еще вокруг нас. Усилиями различных коллективов исследователей во мраке неизведанного время от времени удается высвечивать контуры тех или иных форм, основания которых продолжают теряться в неизвестности. Недаром говорят, что у науки есть одно печальное свойство — она рассеивает очарование тайны.
Фитоиммунология — относительно молодая дисциплина. Все основные открытия у нее еще впереди. И мы надеемся, что в приоткрытую нами дверь в эту интереснейшую область науки войдут новые исследователи, которые и совершат эти открытия.
ПОСЛЕСЛОВИЕ
Когда материалы этой книги были уже подготовлены к печати, скончался один из ее авторов — заслуженный деятель науки, профессор Лев Владимирович Метлицкий, который почти в течение четверти века бессменно возглавлял лабораторию иммунитета растений Института биохимии им. А. II. Баха АН СССР. Профессор Л. В. Метлицкий являлся создателем оригинального направления в фитоиммунологии, согласно которому главная функция иммунитета растений, как и животных, состоит не только в защите организма от инфекционных болезней, по и в поддержании его структурной и функциональной целостности на основе способности организма распознавать и отторгать генетически чужеродную информацию.
Профессор Л. В. Метлицкий был признанным лидером советской школы индуцированного фитоиммунитета. Под его руководством впервые в стране развернулись исследования по изучению индуцированных антибиотиков высших растений — фитоалексинов, а также индукторов и супрессоров их образования. На этом основании им совместно с доктором биологических наук О. Л. Озерецковской был разработан и апробирован принципиально новый метод иммунизации растений, основанный на использовании тех же механизмов, с помощью которых растения защищаются в природных условиях.
Профессор Л. В. Метлицкий постоянно стремился реализовать на практике результаты своих теоретических достижений. Он обладал обостренным чувством нового и никогда не упускал из виду конечной цели своих исследований. Методы его работы и объекты всегда определялись поставленной целью.
Профессору Л. В. Метлицкому был присущ незаурядный научно-организаторский талант. Он обладал способностью притягивать к себе людей, вовлекать их в орбиту своей деятельности и заражать их присущим ему оптимизмом. Природа щедро наградила профессора Л. В. Метлицкого острым и проницательным умом, тонким чувством юмора, смелостью мысли и поступков.
В лице профессора Л. В. Метлицкого наука потеряла одного из ведущих фитоиммунологов, крупного научного организатора и обаятельного человека.
Член-корреспондент АН СССР И. В. Березин
ИЛЛЮСТРАЦИИ[21]


Рис. 4. Естественная (а) и раневая (б) перидерма картофеля

Рис. 5. Реакция СВЧ картофеля на проникновение несовместимой расы фитофторы

Рис. 6. Опыт Мюллера с заражением картофеля фитофторой

Рис. 11. Молекулярная мимикрия (пояснения в тексте)

Рис. 14. Схема взаимной индукции в системе паразит — хозяин (пояснения в тексте)
СПИСОК РЕКОМЕНДУЕМОЙ ЛИТЕРАТУРЫ
Арчаков А. П. Оксигеназы биологических мембран: 32-е Баховское чтение, М.: Наука, 1983. 54 с.
Вавилов Н. И. Проблемы иммунитета культурных растении. М.; Л.: Наука, 1964. Т. 4. 516 с.
Ван-дер-Планк Я. Устойчивость растений к болезням. М.: Колос, 1972. 253 с.
Ван-дер-Планк Я. Генетические и молекулярные основы патогенеза у растений. М.: Мир, 1981. 236 с.
Вердеревский Д. Д. Иммунитет растений к инфекционным заболеваниям. Кишинев: Картя Молдовеняска, 1968. 216 с.
Генетические основы селекции на иммунитет. М.: Наука, 1973. 231 с.
Говалло В И. Парадоксы иммунологии. М.: Знание, 1983. 166 с.
Гойман Э. Инфекционные болезни растений. М.: Изд-во иностр, лит., 1954. 608 с.
Гольшин П. М. Фунгициды в сельском хозяйстве. М.: Колос, 1982. 270 с.
Грунтенко Е. В. Иммунитет «за» и «против». М.: Знание, 1982. 206 с.
Девеполл В. Дж. Защитные механизмы растений. М.: Колос, 1980. 128 с.
Дьяков IO. Т. Физиолого-биохимические основы устойчивости растений к грибным болезням: — В кн.: Итоги науки и техники. Защита растений. М.: ВИНИТИ, 1981, т. 3, с. 5.
Дьяков Ю. Т. О болезнях растений. М.: Колос, 1985.
Купер Э. Сравнительная иммунология. М.: Мир, 1980. 422 с.
Метлицкий Л. В., Озерецковская О. Л. Фитоиммунитет. М.: Наука, 1968. 92 с.
Метлицкий Л. В., Озерецковская О. Л. Фитоалексины. М.: Наука, 1973. 175 с.
Петров Р. В. Беседы о новой иммунологии. М.: Мол. гвардия, 1976. 222 с.
Петров Р. В. Иммунология. М.: Медицина, 1983. 367 с.
Попкова К. В. Учение об иммунитете растений. М.: Колос, 1979, 271 с.
Прокофьев О. Н. Защита растений: настоящее и будущее. Новосибирск: Наука, 1983. 159 с.
Рассел Г. Селекция растений на устойчивость к вредителям и болезням. М.: Колос, 1982. 420 с.
Рубин В. А., Арциховская Е. В., Аксенова В. А. Биохимия и физиология иммунитета растений. М.: Высш. шк., 1975. 320 с.
Румянцев С. И. Конституциональный иммунитет и его молекулярно-экологические основы. М.: Наука, 1983. 209 с.
Стратегия борьбы с вредителями, болезнями растений
и сорняками в будущем. М.: Колос, 1977. 381 с.
Сухоруков К. Т. Физиология иммунитета растений. М.: Изд-во АН СССР, 1952. 147 с.
Тарр С. Основы патологии растений. М.: Мир, 1975. 587 с.
Токин Б. П. Целебные яды растений: Повесть о фитонцидах. 3-е изд. Л.: Изд-во ЛГУ, 1980. 279 с.
Уилсон Л. Тело и антитело. М.: Мир, 1974. 286 с.
Эванс Э. Болезни растений и химическая борьба с ними. М, Колос, 1971, 286 с.
INFO
М 54 Метлицкий Л. В., Озерецковская О. Л. Как растения защищаются от болезней. М.: Наука, 1985. 192 с. (Серия «Наука — сельскому хозяйству».)
38.3
М 3803040000-432/042 (02)-85 *88-85-НП
[Лев Владимирович Метлицкий]
Ольга Леонидовна Озерецковская
КАК РАСТЕНИЯ
ЗАЩИЩАЮТСЯ ОТ БОЛЕЗНЕЙ
Утверждено к печати
Редколлегией серии научно-популярных изданий
Академии наук СССР
Редактор издательства Е. Р. Воронцова
Художник В. П. Хлебников
Художественный редактор Н. А. Фильчагина
Технические редакторы З. Б. Павлюк, М. Л. Анучина
Корректоры Е. Н. Белоусова, Н. Г. Васильева
ИВ № 29028
Сдано в набор 26.04.85
Подписано к печати 24.07.85
Т-15517. Формат 84X108 1/32.
Бумага книжно-журнальная. Импортная
Гарнитура обыкновенная
Печать высокая
Усл. печ. л. 10,29. Усл. кр. отт. 10, 5. Уч. изд. л; 11,1.
Тираж 38 000 экз. Тип. зак. 1370
Цена 70 коп.
Ордена Трудового Красного Знамени
издательство «Наука»
1117864 ГСП-7, Москва В-485.
Профсоюзная ул., 90
2-я типография издательства «Наука»
121099, Москва, Г-99, Щубинский пер., 6
…………………..
FB2 — mefysto, 2024
Примечания
1
М.: Колос, 1979. 271 с.
(обратно)
2
М.: Мир, 1974. 286 с.
(обратно)
3
М.: Знание, 1983. 166 с.
(обратно)
4
Лонгфелло Г. Песнь о Гайавате/Пер. И. А. Бунина. М.: Дет. лит., 1956, с. 41.
(обратно)
5
Рассел Г. Селекция растений на устойчивость к вредителям и болезням. М.: Колос, 1982. с. 8.
(обратно)
6
Поляков Н. М. Защита растений — проблемы и перспективы, — Наука и жизнь, 1972, № 9, с. 70.
(обратно)
7
Гамзатов Р. Мой Дагестан. — Собр. соч.: В 5-ти т. / Пер. В. Солоухина. М.: Худож. лит., 1981, т. 4, с. 7.
(обратно)
8
Уилсон Д. Тело и антитело. М.: Мир, 1971, с. 16.
(обратно)
9
Петров Р. В. Иммунология. М.: Медицина, 1983, с. 10.
(обратно)
10
Дьяков Ю. Т. Предисловие. — В кн.: Генетические основы селекции растений на иммунитет. М.: Наука, 1973, с. 8,
(обратно)
11
Рассел Г. Селекция растений на устойчивость к вредителям и болезням. М.: Колос, 1982, с. 11.
(обратно)
12
М.: Мир, 1975, с. 178.
(обратно)
13
Вавилов Н. И. Проблемы иммунитета культурных растений — Избр. тр. М.; Л.: Наука, 1964, т. 4, с, 331,
(обратно)
14
Там же, с. 317,
(обратно)
15
Токин Б. П. Целебные яды растений: Повесть о фитонцидах. 3-е изд. Л.: Изд-во ЛГУ, 1980, с. 278.
(обратно)
16
Петров Р. В. Предисловие. — В кн.: Уилсон Д. Тело и антитело. М.: Мир, 1974,
с. 7.
(обратно)
17
Купер Э. Сравнительная иммунология. М.: Мир, 1980, с. 25.
(обратно)
18
Гете И. Фауст/Пер. Н. Холодковского. М.: Дет. лит., 1977, с. 112, 140
(обратно)
19
Прокофьев О. Н. Защита растений: настоящее и будущее. Новосибирск: Наука, 1983, с. 159.
(обратно)
20
Цит. по:
Вавилов Н. И. Природа иммунитета, — Избр. тр. М.; Л., Наука, т. 4, с. 317.
(обратно)
21
Вклейка, упоминаемая в тексте —
Примечание оцифровщика.
(обратно)
Оглавление
ОТ АВТОРОВ
ЖИЗНЬ РАСТЕНИЙ ПОД УГРОЗОЙ
ОСНОВОПОЛОЖНИК УЧЕНИЯ
О ФИТОИММУНИТЕТЕ
МЕДИЦИНСКАЯ ИММУНОЛОГИЯ
И ИММУНОЛОГИЯ РАСТЕНИИ
КТО, ЧТО И КАК ВЫЗЫВАЕТ БОЛЕЗНИ РАСТЕНИЙ
ВОЗБУДИТЕЛИ БОЛЕЗНЕЙ РАСТЕНИИ
И ОРУДИЯ ИХ АГРЕССИИ
СОПРЯЖЕННАЯ ЭВОЛЮЦИЯ РАСТЕНИЯ И ПАТОГЕНА,
ИЛИ КАК САПРОФИТ СТАЛ ПАРАЗИТОМ
СТРАТЕГИЯ САМООБОРОНЫ
СУММА СЛАГАЕМЫХ
РАНЕВОЙ СТРЕСС
РАЗОРУЖЕНИЕ ПАРАЗИТА
МЕХАНИЧЕСКИЕ ПРЕГРАДЫ
СВЕРХЧУВСТВИТЕЛЬНОСТЬ,
ИЛИ ГИБЕЛЬ ЧАСТИ ВО ИМЯ
СПАСЕНИЯ ЦЕЛОГО
ИММОБИЛИЗАЦИЯ ПАТОГЕНА
ФИТОНЦИДЫ
ФИТОАЛЕКСИНЫ
ЛИШЕНИЕ ПАТОГЕНА САМОГО НЕОБХОДИМОГО
ЧТО ТАКОЕ КСЕНОБИОТИКИ
И КАК ОНИ УНИЧТОЖАЮТСЯ
РАСПОЗНАТЬ,
ЧТОБЫ ПРИНЯТЬ ИЛИ УНИЧТОЖИТЬ
КТО ЕСТЬ КТО
ИНДУКТОРЫ, ЭЛИСИТЕРЫ, ПРОВОКАТОРЫ
ЗАГАДКА СПЕЦИФИЧНОСТИ
СЕНСИБИЛИЗАЦИЯ,
ИЛИ ПОВЫШЕНИЕ ЧУВСТВИТЕЛЬНОСТИ
ОТ ТЕОРИИ К ПРАКТИКЕ
ИНТЕГРИРОВАННАЯ СИСТЕМА
ЗАЩИТЫ РАСТЕНИИ
СОРТ РЕШАЕТ УСПЕХ ДЕЛА
ПЕСТИЦИДЫ:
ВЧЕРА, СЕГОДНЯ, ЗАВТРА
ИММУНИЗАЦИЯ
ВСЕ ВЫРАЩЕННОЕ СОХРАНИТЬ
ЗАКЛЮЧЕНИЕ
ПОСЛЕСЛОВИЕ
ИЛЛЮСТРАЦИИ[21]
СПИСОК РЕКОМЕНДУЕМОЙ ЛИТЕРАТУРЫ
INFO
*** Примечания ***